
P.C. Korir
Not Available
J.O. Nyabundi
Not Available
P.K. Kimurto
Not Available
Asian Journal of Plant Sciences
Year: 2006 | Volume: 5 | Issue: 1 | Page No.: 24-32







ABSTRACT
The aim of this study was to determine drought response of thirty bean genotypes under glasshouse and field conditions. The experiment was laid in a Randomized Complete Block in split-plots design with three replications. Significant (p<0.05) genotypic differences in drought resistance were observed under moisture stress conditions. Significant interaction (p<0.01) between variety and moisture level were observed both in the glasshouse and in the field. Drought resistant genotypes showed high relative water content in stress environment as compared to susceptible ones. They also took relatively longer time to wilt, consequently showing low soil water content at wilting and lower biomass reductions under moisture stress conditions. A significant negative correlation was obtained between days to permanent wilting and relative water with biomass reduction. Relative water content was positively and significantly correlated to stomatal conductance and days to permanent wilting. Correlation between stomatal conductance and biomass yield reduction was not significant. The responses relative water content and stomatal conductance, in certain conditions, were recognized as beneficial drought resistance indicators and may be used as selection criteria in bean breeding programme.
PDF Abstract XML References Citation
How to cite this article
P.C. Korir, J.O. Nyabundi and P.K. Kimurto, 2006. Genotypic Response of Common Bean (Phaseolus vulgaris L.) to Moisture Stress Conditions in Kenya. Asian Journal of Plant Sciences, 5: 24-32.
DOI: 10.3923/ajps.2006.24.32
URL: https://scialert.net/abstract/?doi=ajps.2006.24.32
DOI: 10.3923/ajps.2006.24.32
URL: https://scialert.net/abstract/?doi=ajps.2006.24.32
INTRODUCTION
Common bean (Phaseolus vulgaris L.) is one of the most important crops grown in the semi-arid areas of Kenya. It is the most important source of proteins in Kenya and ranks second to maize in importance as source of food, especially amongst rural communities. Drought conditions in Kenya are widespread covering up to 80% of landmass, therefore beans are frequently exposed to drought stress conditions. Studies on the crop in these regions indicate a wide genetic variability in drought resistance among the unimproved bean cultivars[1].. Information on drought resistance amongst improved cultivars was reported by Nyabundi et al.[2], however, the information is inadequate and therefore there is need to test various genotypes to ensure that losses due to drought are reduced. To achieve this, identification of traits associated with drought is necessary. Runkulatile et al.[3] reported that land races from dry areas maintain a consistently higher stomatal conductance, transpiration rates, net photosynthesis and relative water content. Alternatively, Mcree and Richardson[4] suggested that the ability of a drought adapted variety to root deeper in the soil horizon and extract more water from the receding waterfronts may help maintain high relative water content of the plant. This is associated with high photosynthetic activity without a significant reduction in stomatal closure and transpiration rate. Cultivars with these abilities tend to exhibit delay in dehydration as indicated by the number of days to permanent wilting with the onset of drought[5]. In common beans, information on responses of these traits to drought stress is scanty, especially in tropical regions.
Relative Water Content (RWC) has been suggested to be a better measure for plant’s water status than the thermodynamic state variables (water potential, turgor potential and solute potential). Slatyer[6] showed that when the water potential was -20 bars, the RWC was 50% in tomato leaves, but about 90% in Acacia anaeura phyllodes, implying that the tomato plant had a higher water deficit than Acacia. Similarly, Fischer[7] found that RWC was directly related to soil water content and suggested that RWC might also be used to indicate soil water. Leaf RWC in plants decrease as water stress levels increase. It would be expected that under non-stress conditions, RWC of the plant would be near 100% with water potential values approaching zero. Bennet et al.[8] and Schonfeld et al.[9] noted that superior performance of drought tolerant soybeans, maize and wheat varieties under water stress environment is attributed to osmoregulation and stomatal closure when stress sets in. These cultivar differences in RWC could be used to select high yielding genotypes that maintain cell turgor under water stress environment to give high relative yield[10]. Water stress reduces plant growth more than all other stresses combined[11]. Bradford and Hsiao[12] demonstrated that small reductions in growth rates in the early growth stage due to water stress would, because of exponential nature, compound with time into large reductions in biomass productions. Water stress depresses dry matter production in most crops[13,14] but it is the stress intensity, duration of application and crop’s growth stage that determines the degree of biomass depression[15]. There is little information on the response of Kenyan land races and their crosses to drought stress. Information on selection criteria is also scanty. Therefore, the objective of this study was to evaluate the value of morphological and physiological traits as indicators that can be used in selection for drought resistance in common beans. The hypothesis tested was that common bean response to water stress was similar across all the commercial check land races and their crosses and that no trait was associated with drought in beans.
MATERIALS AND METHODS
Site description: The study was conducted between March 1994 and March 1994 in Central province of Kenya, at Kabete Campus, University of Nairobi (1°45 20 S and 36°45 20 E). The site is in agriculturally high potential zone, LH3, within the Kenya highlands[16]. Kabete is at an altitude of 1829 m above sea level and receives annual rainfall of about 1000 mm with a mean monthly maximum temperature of 32°C and minimum of 12°C. The soils are well-drained friable clay with humic Nitosols. The mean minimum and maximum glasshouse temperature during the experimental period was 14.8 and 36.2°C, respectively.
Bean genotypes: Thirty bean genotypes (Table 1) obtained from the Bean Project of the Department of Crop Science; University of Nairobi comprised five seed types namely Rose Coco (RC), Canadian Wonder (CW), Mwitemania (MW), Mwezi Moja (MM) and Red Haricot (RH). Each seed type had a number of nearly homozygous F7, F8 and F9 generation bean lines selected from populations created from crosses among seven parents including five local commercial cultivars GLP-2, GLP-24, GLP-92, GLP-585 and GLP-1004 which were included in this study. Based on the days to maturity, the genotypes were designated as Early (E), Medium (M) or late (L) maturing with days to maturities of 80 to 85, 86 to 96 and more than 96 days, respectively.
| Table 1: | Bean cultivars used in this study |
 | |
| RC = Rose Coco, CW = Canadian Wonder, MW = Mwitemania, MM = Mwezi Moja, RH = Red Haricot, ** Commercial check cultivars, E = Early, M = Medium and L = Late maturing, + Cultivars used in determination of RWC in the field | |
Experimental design and sampling procedures: Thirty genotypes were planted in the glasshouse in March to May 1994 in 8L pots. Each pot comprised of topsoil mixed with gravel and sand in a ratio of 2:1:1 and DAP (18:46:0) at the rate of 100 kg ha-1. A 2 x 30 factor experiment was arranged as a split-plot in Randomized Complete Block Design replicated three times. Two watering levels (water stress and water non-stress) were assigned as whole plots while the genotypes were ascribed to the sub-plots. Six seeds were planted in each pot and watered to field capacity. The seedlings were thinned to four per pot seven days after emergence. Stress treatment was initiated at the third trifoliate leaf stage (at about 28 Days After Emergence (DAE)) by bringing all the pots to field capacity in the morning of day zero. There after, water was withheld until dawn time (6 am) wilting was evident in each genotype. This was assumed to be the permanent wilting point[17]. Data in the glasshouse were taken on three randomly selected plants from each sub-plot (genotype) for the leaf RWC, Days to Permanent Wilting (DPW), Soil Moisture Content at wilting (SMC) and stomatal conductance at wilting. The RWC values were determined as the water content at sampling relative to that at full turgor as described by Bennet et al.[8], thus percent RWC = (Fresh weight-dry weight/turgid weight- dry weight) x 100. Days to permanent wilting represented the number of days from when water was withheld till dawn time wilting was evident in each genotype. Stomatal conductance (cm sec-1) was determined on the abaxial leaf surface of wilted plants using a steady state porometer (Li-1600). Soil moisture content at wilting was determined gravimetrically by oven-drying the soil sample at 105°C for 24 h to get the dry weight for each genotype. The soil moisture content was expressed as percent by weight using the formula: % Water by weight = (Wet weight-dry weight/dry weight) x 100[18].
The field experiment was conducted during the dry spell between January and March 1995. Sixty treatments were arranged as a split-plot in a RCBD replicated three times. Three levels of watering (high, medium and low) were assigned to whole plots measuring 2x30 m. The sub-plots measured 1x2 m and the treatments consisted of the genotypes, each in five rows spaced 0.5 m apart. Seeds were planted at an intra-row spacing of 10 cm giving a plant population of 200,000 plants ha-1. Seedlings were thinned to one per hill ten days after emergence. Irrigation water was applied using a line source sprinkler irrigation set that provided a moisture gradient, decreasing with increase in distance from the sprinkler line. Sprinkler stands were spaced at 30 m and sprinkler throw of about 15 m was achieved. The high, medium and low level of watering treatments received a total of 413, 312 and 174 mm of water, respectively. These amounts included little rainfall (144 mm) that fell during the experimental period. Field data were collected from each sub-plot region receiving designated amounts of water (excluding double irrigated overlap areas) and included the following response variable: Total plant biomass accumulation, leaf relative water content and biomass yield reductions from the high, medium and low watering levels. Total aboveground biomass was determined on five plants per sub-plot at fortnightly interval starting 14 DAE. The plants were cut at ground level and dried at 70°C for 72 h to constant weight. Leaf RWC was measured as described for the glasshouse experiment at 14 day interval, beginning 15 DAE up to 43 DAE. The biomass yield reductions for the medium and low water treatments were expressed as percentage of the high water treatment biomass. The high water level biomass was assumed to be non-stress while the medium and low water levels were differentially stressed, thus biomass yield reduction was computed as follows: % Biomass reduction = (Biomass stress/biomass non-stress) x 100.
Data analysis: Data were subjected to analysis of variance using the General Linear Model (GLM) and means separated by Duncan’s Multiple Range Test (p<0.05) on SAS software[19]. Correlation analysis was conducted for the data according to Steel et al.[20].
RESULTS
The mean RWC across the genotypes in the glasshouse water stress treatment generally decreased with increasing number of Days After Withholding Water (DAWW) (Table 2) with the highest mean value being 87.59% (0 DAWW) and lowest value of 68.92% at wilting. The mean leaf RWC across the genotypes remained relatively stable throughout the first three days after watering was withheld, however, by the sixth day, the mean leaf RWC across the genotypes had dropped to 80.88%. Further drops to means of 73.6 and 69.31% were similarly observed for nine and twelve DAWW and at wilting, respectively. At dawn time wilting, cultivars M19, M16, M18, GLP-1004 and GLP-92 had the highest RWC means while E4, L46, M15, L37 and GLP-585 had the lowest mean values at the same time. Cultivar GLP-1004 took the longest period to permanent wilting (22 days) followed by GLP-92 and M18 (21 days each) and M19 that wilted after 20 days (Table 3). These genotypes appeared to maintain cell turgidity even at comparatively low soil moisture contents at which wilting occurred. These genotypes maintained their stomatal conductance at high values (> 0.30 cm sec-1) despite the low moisture contents at wilting. In contrast, E6, E10, M11, M14, M15, M26 and L37 took minimum time to loose turgidity, wilting after 12 days. At this time, the soil moisture contents were comparatively high and the stomatal conductance values were low compared with the seemingly drought resistant genotypes such as GLP-1004, GLP-92, M16 and L44, which had lower SMC and higher stomatal conductance under the same treatment conditions. Days to Permanent Wilting (DPW) were significantly negatively correlated to soil water content at wilting (r= 0.88**)(Data not shown). It was however significantly and positively correlated to leaf relative water content at wilting (r=0.81*). Significant negative correlation was found between soil moisture content at wilting and leaf RWC at wilting (r=-0.65*). Relative water content at zero DAWW associated significantly positively with DPW but negatively with SMC in the glasshouse (r= 0.54* and -0.51*, respectively).
| Table 2: | Relative water content (%) for stressed bean genotypes at different DAWW |
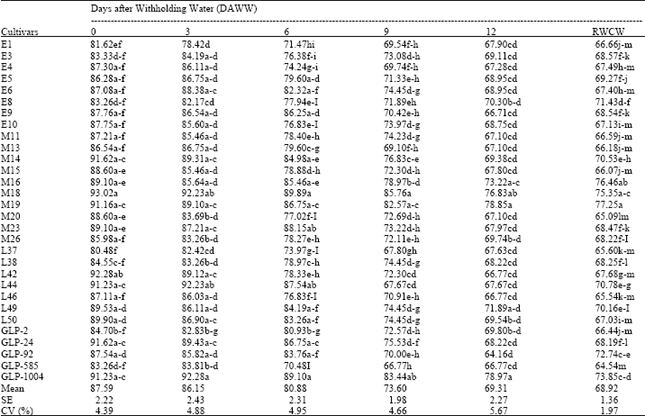 | |
| RWCW = Relative Water Content at Wilting | |
| Table 3: | Days to permanent wilting, stomatal conductance and percent soil water content at wilting, for bean cultivars in the glasshouse experiment |
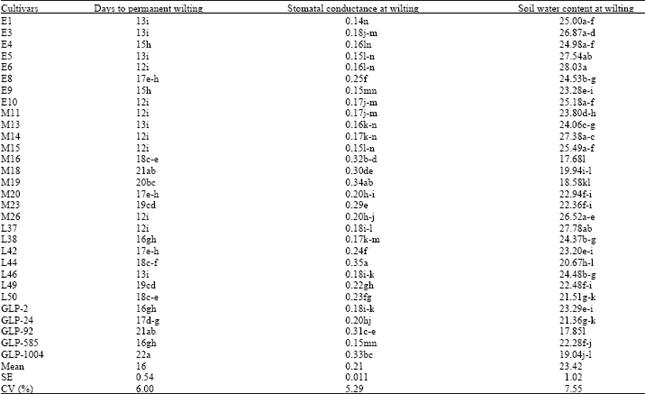 | |
| Table 4: | Varietal differences in percent leaf relative water content under different watering levels at different DAE of field experiment |
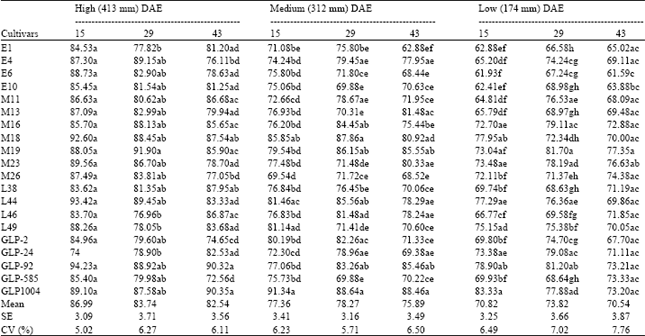 | |
| Table 5: | Varietal differences in biomass accumulation (g m-2) under different watering levels at different DAE |
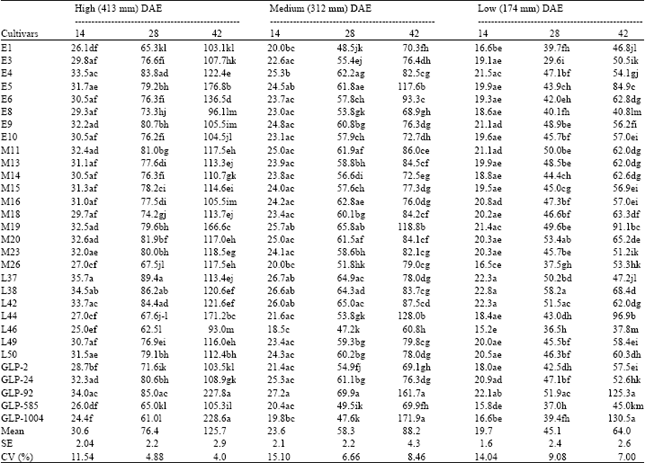 | |
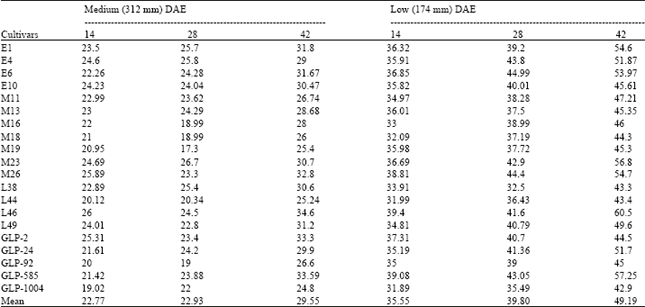
| Table 6: | Biomass yields reduction of twenty bean cultivars at different DAE under medium and low watering levels |
 | |
There were varietal differences in percent leaf RWC of the three watering levels at different DAE for the field experiment (Table 4). At 43 DAE of the high water level, varieties GLP-1004, GLP-92, L38 and M18 had relatively higher mean absolute RWC values, while comparatively lower values were scored by the cultivars GLP-585, GLP-2, M26 and E4. Higher mean values were noted with cultivars GLP-1004, GLP-92, M19 and M18 at 43 DAE of the medium watering level. Among the cultivars that exhibited relatively higher RWC values at 43 DAE of the low water treatment were M19, M23, M26, GLP-92 and GLP-1004. Significant watering x variety interaction was observed at 15 DAE (p<0.05) and 29 DAE (p<0.01). It can be deduced from the above observations that the cultivars GLP-1004, GLP-92, M18 and M19 maintained fairly high relative water content under the three watering levels while genotypes E6, M26, GLP-2 and GLP-585 were among those with lowest values under similar conditions.
Plant biomass was significantly (p<0.01) reduced by water stress at 28 and 42 DAE in all cultivars (Table 5). Significant watering x variety interaction was observed at 14 DAE (p<0.05) and at 28 and 42 DAE (p<0.01). The mean across the genotypes reveals that biomass accumulation was lower at low irrigation level followed by medium level at similar number of days after emergence. Though there was a general increase in biomass accumulation with the number of days after emergence, the increase was affected by the irrigation level, being highest in the high irrigation level and lowest in the low level. At 42 DAE, genotypes GLP-1004, GLP-92, L44, M19 and E6 had significantly more biomass under all the watering levels. GLP-1004 had the highest biomass followed by GLP-92 for all watering levels at 42 DAE. Varieties E1, L46 and GLP-2 were among those with the least values at this time under the three watering levels. The mean percent biomass reduction in the medium water treatment was lower than that of the low treatment at all DAE (Table 6). The reduction tended to increase with the number of days after emergence under both treatments, but the reduction was marginally small between 14 and 28 DAE under medium water treatment, being 0.16%. The margin under low water treatment between these two samplings was comparatively large. Greatest mean yield reduction of 49.19% was observed at 42 DAE under low watering level. At 42 DAE in medium water treatment, cultivars L46, GLP-585, GLP-2, M26, E1 and E6 suffered the greatest biomass yield reductions. However, least biomass reduction was observed in GLP-1004, L44, M19, M11, GLP-92 and M18. Depression of biomass yield at 42 DAE in low water treatment was more pronounced. Genotypes L46, GLP-585, M23, E1, E6 and M26 experienced the greatest reductions. Of the cultivars that suffered least biomass reductions at 42 DAE of medium water treatment, GLP-1004, L44, M19 and M18 still had relatively least percent reductions at 42 DAE in the low water treatment. Days to permanent wilting correlated negatively and significantly with percent biomass reduction at 42 DAE, low watering level (r = -0.63*) of the field experiment. However, SMC and RWC at wilting respectively associated positively (r = 0.58*) and negatively (r =-0.53*) with percent biomass reduction at similar DAE of low watering level.
DISCUSSION
The bean genotypes tested in this study showed variability in phenology and physiological responses to water deficits. Water stress decreased leaf relative water content and biomass in all the tested genotypes. Similar observations have been reported in blackgram, greengram, cowpeas and soybeans[21], in common bean[3] and in sugar beet[4]. Variety differences were observed in leaf RWC in the glasshouse and in the field. Varieties M18, L44, GLP-24, M19, GLP-1004 and GLP-92 had consistently higher RWC values in the glasshouse and in the medium and low watering levels in the field. This observation indicated that these genotypes had a better plant water status and hence better yields as compared with those that had generally lower scores for the traits under similar conditions.
This genotypic variation in RWC may be attributed to differences in the ability of the varieties to absorb more water from the soil and/or the ability to control water loss through the stomates. These findings are in agreement with those reported by Sinclair and Ludlow[22]. It may also be due to differences in the ability of the tested varieties to accumulate and adjust osmotically to maintain tissue turgor and hence physiological activity[9]. Varietal differences in RWC may also be a result of varieties maximizing on soil water reserves by fully extracting water in the existing rooting zone and/or extending rooting depth to increase water reserve for the crop[23]. Other researchers have investigated the role of increased root length and density in maintenance of guard cell turgidity under water stress. Lorens et al.[24] attributed the cultivar differences in net photosynthetic rates to the differential abilities for the better-adapted cultivars to root deeper and thus maintain higher plant water potential. Runkulatile et al.[3] found that drought resistant varieties (Ulonzo, GLP-1004 and White Haricot) exhibited faster downward root penetration than the less drought resistant varieties. In the present study, the findings concur as drought resistant genotypes such as GLP-1004, GLP-92, M18 and M19 showed higher RWC and stomatal conductance apart from taking longer periods to wilt and at low pot soil moisture content at wilting under glasshouse compared to the drought susceptible genotypes such as E6, E10, M11, M26 and L37. These genotypes showed similar trends in the field under the medium and low watering levels. The drought resistant genotypes consistently maintain higher RWC and suffered least biomass reductions as compared to their drought susceptible counterparts.
Osmotic adjustment would also explain the maintenance of high RWC values of these cultivars in the glasshouse, despite the low soil moisture contents at wilting. Martin et al.[25] proposed the mechanism for drought resistant barley cultivars that maintained high RWC under water stress. Similarly, stomatal conductance may be a useful indicator in discriminating drought resistant and non-resistant cultivars in the glasshouse. However, water stress develops quickly in the glasshouse due to limited soil volume in the pots. Therefore, plants may not have time to adjust their stomata as in the field where response to water stress may allow for osmoregulation, which causes a relatively higher RWC and stomatal conductance. Water stress depends on variety and to a greater extent on the length of its exposure, plus that of temperature in the growing conditions[26].
The delay in dehydration exhibited by the cultivars GLP-1004, GLP-92, GLP-24, M18, M19, L44 and L50 may have important implications in terms of crop productivity. These varieties had comparatively higher RWC values under stress conditions, tended to accumulate higher biomass yields and suffered least depressions in biomass under all levels of water stress. Days to permanent wilting for these genotypes were longer than their counterparts such as E6, E10, M11, M14 and M15 but the soil water content at their permanent wilting was comparatively lower. Since cultivars such as GLP-1004, GLP-92, GLP-24, M16, M18, M19 and L44 had comparatively higher RWC in the glasshouse treatment, it may possibly be concluded that they maintain a relatively higher water potential gradient between the roots and the transpiring leaves. This discrepancy could be due to the differences in leaf solute potentials since the evaporative demand and the initial soil water content was similar for all the varieties. Differences in root length density may also have played part, but this factor is not likely to be important in influencing water uptake by potted plants whose roots ramify extensively into the potting medium. However, it could come into play in the field grown plants as has been reported by Runkulatile et al.[3]. Morgan and Condon[27] and Santamaria et al.[17] also showed that genotypes of wheat and sorghum with high osmotic adjustment produced more root biomass and greater root length than genotypes with low osmotic adjustment. The results of this study therefore may suggest that osmotic adjustment principally enabled the cultivars GLP-1004, M18, M16, M19 and GLP-24 to exploit moisture from relatively dry soils and thus taking comparatively longer periods to wilt.
Differential effect of watering level on biomass accumulation was observed at all days after emergence. But due to inherent genetic differences in biomass accumulation among genotypes, percent decrease in biomass accumulation would provide a better indicator of drought resistance than differences in total biomass per se. Cultivars such as GLP-1004, L44, M18 and M19 were least affected under both low and medium watering levels compared to GLP-585, E1, E6, or M23. This disparity in their response could also serve as evidence that the former genotypes have higher ability to resist drought. Their higher RWC in the glasshouse and in the field could support their ability to osmoregulate and thus maintained growth under water-limited conditions. Similarly, studies on wheat[27] and sorghum[17] have indicated that osmotically non-adjusting varieties are more significantly affected by water stress than the osmotically adjusting.
The combination of both morphological and physiological attributes of drought resistance with high yield potential forms the selection basis for better yield performance under drought conditions. Although common beans are prone to short water stress periods, differences in drought resistance have been observed in this study. There was evidence that the genotypes that showed superior performance in the glasshouse also suffered fairly low biomass reduction under the medium and low watering levels in the field. Strong correlations occurred between traits such as days to permanent wilting, leaf relative water content at wilting and soil moisture content at wilting as well as stomatal conductance at wilting of the glasshouse and biomass reduction in the field. These suggest the possibility of screening the genotypes in the glasshouse and also the likelihood of selecting the cultivars on the basis of their leaf RWC. Therefore, cultivars M16, M18, M19, L44, GLP-24, GLP-92 and GLP-1004 seem to be most adapted to the dry-land conditions among the tested genotypes.
REFERENCES
- Mccree, K.J. and S.G. Richardson, 1987. Stomatal closure vs osmotic adjustment a comparison of stress responses. Crop Sci., 27: 539-543.
Direct Link - Schonfeld, M.A., R.C. Johnson, B.F. Carver and and D.W. Mornhinweg, 1988. Water relations in winter wheat as drought resistant indicators. Crop Sci., 28: 526-531.
Direct Link - Santamaria, J.M., M.M. Ludlow and S. Fukai, 1990. Contribution of osmotic adjustment to grain yields in sorghum under water-limited conditionswater stress before anthesis. Aust. J. Agric. Res., 41: 51-65.
CrossRef - Sinclair, T.R. and M.M. Ludlow, 1985. Who taught plants thermodynamics? The unfulfilled potential of plant water potential. Aust. J. Plant Physiol., 12: 213-217.
Direct Link - Cortes, P.M. and T.R. Sinclair, 1986. Water relations of field-grown soybean under drought. Crop Sci., 26: 993-998.
Direct Link - Lorens, G.F., J.M. Bennett and L.B. Loggale, 1987. Differences in drought resistance between two corn hybrids. 1: Water relations and root length density. Agron. J., 79: 802-807.
Direct Link - Ackerson, R.C., 1981. Osmoregulation in cotton in relation to osmotic adjustment. Plant Physiol., 67: 489-498.
PubMed