
Mohannad G. AL-Saghir
Not Available
Duncan M. Porter
Not Available
Asian Journal of Plant Sciences
Year: 2006 | Volume: 5 | Issue: 6 | Page No.: 1002-1006







ABSTRACT
Phylogenetic relationships among four Pistacia species were studied by randomly amplified polymorphic DNA (RAPD) analysis. The studied species were: P. khinjuk. P. lentiscus, P. terebinthus and P. vera. Genomic DNA was extracted from leaf tissue and RAPD analysis was performed using 23 primers. A total of 248 fragments were generated and 139 bands were polymorphic at the inter-specific level. Subjecting these data to phylogenetic analysis yielded a shortest cladogram that is 34 steps long comprising two main groups. P. khinjuk, P. terebinthus and P. vera, were included in one group, while P. lentiscus formed the second group. The first group included deciduous species with whereas the species included in the second group mostly grow as evergreen shrubs. The cladogram showed that the closest pairs of species were P. khinjuk and we suggest that P. khinjuk may be a descendant of P. vera.
PDF Abstract XML References Citation
How to cite this article
Mohannad G. AL-Saghir and Duncan M. Porter, 2006. Random Amplified Polymorphic DNA (RAPD) study of Pistacia species (Anacardiaceae). Asian Journal of Plant Sciences, 5: 1002-1006.
DOI: 10.3923/ajps.2006.1002.1006
URL: https://scialert.net/abstract/?doi=ajps.2006.1002.1006
DOI: 10.3923/ajps.2006.1002.1006
URL: https://scialert.net/abstract/?doi=ajps.2006.1002.1006
INTRODUCTION
It was planned to use DNA sequence analysis in this study. However, I found that Tingshuang Yi and Jun Wen from the Department of Botany of the Field Museum of Natural History, Chicago (2005, Personnel Communication) have performed a 5-gene analysis of Pistacia to study its phylogeny and biogeography using both nuclear and chloroplast genes (NIA-i3 region, ITS4 and ITS5, trnC-trnD region, ndhF gene and trnL-F regions), which was exactly what I had planned to do. Therefore, present research switched to RAPD technology to study relationships in Pistacia.
Certain properties are desirable for a molecular marker such as highly polymorphic behavior, codominant inheritance, frequent occurrence in the genome, even distribution throughout the genome, selectively neutral behavior, easy access, easy and fast assay and high reproducibility (Weining and Henry, 1995). Examples of such DNA molecular markers are random amplified polymorphic DNA (RAPDs) (Welsh and McCelland, 1990; Williams et al., 1990) and amplified fragment length polymorphisms (AFLPs) (Vos et al., 1995). RAPD markers have proved to be good genetic markers to assay and evaluate the genetic diversity between and within the same species, populations and individuals (Warburton and Bliss, 1996). RAPD markers depend on the amplification of a DNA sequence by the polymerase chain reaction using only a single primer of arbitrary nucleotide sequence. The technique has proved to be fast and simple needs small quantities of template DNA and detects relatively small amounts of genetic variation (Warburton and Bliss, 1996).
Molecular studies addressing the genus Pistacia are few. The pollen isozyme patterns of nine different enzymes in P. lentiscus, P. terebinthus and P. vera were studied by Louskas and Pontikis (1979) in order to assay inter-specific relationships. They found a closer phylogenetic relationship between P. terebinthus and P. vera than between P. lentiscus and P. vera. Isozyme and DNA markers were used to distinguish and characterize the germplasm of P. vera (Hormaza et al., 1994, 1998; Dollo et al., 1995; Vezvaei, 1995; Barone et al., 1996; Dollo 1996; Caruso et al., 1998). Parfitt and Badenes (1997) were the first to provide a classification of 10 Pistacia species at the molecular level. They characterized these species based on chloroplast DNA profiles and subdivided the genus into two sections, Terebinthus (P. atlantica, P. chinensis, P. khinjuk, P. integerrima, P. terebinthus and P. vera) and Lentiscus (P. lentiscus, P. mexicana, P. texana and P. weinmanniflolia).
Kafkas and Perl-Treves (2001) addressed the taxonomic relationships and genetic variation of wild Pistacia germplasm in Turkey using morphological data and random amplified polymorphic DNA (RAPD). In the resulting molecular phylogeny, Pistacia species are clearly separated from each other. Pistacia terebinthus is the most divergent species and the closest pair was P. atlantica and P. eurycarpa. Kafkas and Perl-Treves (2002) addressed the interspecific relationships in Pistacia based on RAPD fingerprinting. The following species were included: P. atlantica, P. eurycarpa, P. khinjuk, P. lentiscus, P. mexicana, P. palaestina, P. terebinthus and P. vera. Phylogenetic analysis yielded two main groups: P. atlantica, P. eurycarpa, P. khinjuk, P. integerrima and P. vera while P. lentiscus, P. mexicana, P. palaestina and P. terebinthus formed the second group. The authors suggested that P. palaestina is a variety of P. terebinthus. This study showed that P. eurycarpa is a distinct species from P. atlantica rather than a variety of that species.
Katsiotis et al. (2003) used RAPD and amplified fragment length polymorphism (AFLP) to study the relationships of native and introduced Pistacia species in Greece. The AFLP and RAPD phenograms were comparable, with minor clustering differences. The results revealed two main groups one contains the evergreen species P. lentiscus and the resin-producing P. lentiscus cv. Chia (cultivated only on Island of Chios) and the other contains the deciduous trees P. palaestina, P. terebinthus and P. vera. Pistacia chinensis was clustered either with the first group (by RAPD results) or with the second one (by AFLP results). Barazani et al. (2003) characterized the genetic variability of Mediterranean P. lentiscus genotypes by morphology, composition of essential oils and RAPD. The RAPD dendrogram showed two main groups according to the species’ geographical origin. One group included the Tunisian, Spanish and Israeli accessions and the second group included Cyprian accessions. Golan-Goldhirsh et al. (2004) assessed the polymorphism among Mediterranean basin Pistacia species and accessions within the same species using RAPD and AFLP markers. Similar results were obtained by the two techniques. The RAPD and AFLP dendrograms showed two major groups with one group containing all the P. lentiscus accessions and the other group containing all other accessions. The latter group subdivided into two subgroups, one consisting of P. palaestina and P. terebinthus, the other consisting of P. atlantica, P. khinjuk and P. vera.
Yi and Wen (2005, personnel communication) have performed a 5-gene analysis of Pistacia to study its phylogeny and biogeography using both nuclear and chloroplast genes (NIA-i3 region, ITS4 and ITS5, trnC-trnD region, ndhF gene and trnL-F regions). They sampled all species of the genus and are preparing the manuscript with two additional collaborators. However, their study did not address the taxonomic relationships within the genus. The objectives of the present study were to investigate and clarify the taxonomic relationships among Pistacia species.
MATERIALS AND METHODS
Plant materials and DNA extraction:The following four Pistacia species were included in this study: P. khinjuk, P. lentiscus, P. terebinthus and P. vera. The seeds of only these four species were available. Seeds of these species were obtained from USDA, National Clonal Germplasm Repository (Davis, CA). Rhus ovata was used as an out group, the seeds being obtained from the Royal Botanic Gardens, Kew. DNA was extracted from the young leaves of greenhouse planted seedlings (4 weeks old) using a DNeasy Plant Mini kit (QIAGEN, Valencia, CA).
RAPD Amplification: Twenty-three random primers (10-mer) (Operon Technologies, Alameda, CA) of arbitrary sequence were used in this study (Table 1).
RAPD reactions were done in a total volume of 25 μL containing 60 ng of primer; 200 μM each of dATP, dCTP, dGTP and dTTP, 100 ng of DNA template, 2 mM MgCL2 and 1.5 u of Taq DNA polymerase in 1X PCR buffer. DNA amplification was performed in a Biometra thermal cycler (Goettingen, Germany) in 0.2 mL PCR tubes programmed for initial denaturation at 95°C for 2 min followed by 43 cycles for 1 min at 95°C, at an annealing temperature of 37°C for 1 min and at 72°C for 2 min as an extension step. The final extension step was done for 5 min at 72°C and the reactions were kept at 4°C.
| Table 1: | Random primers used in this study to screen RAPD markers |
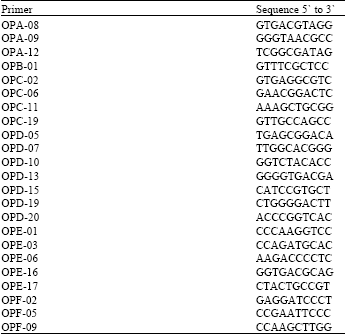 | |
The RAPD-PCR amplified products were analyzed by gel electrophoresis in 1.5% ultrapure agarose in 0.5X TBE buffer stained with ethidium bromide (0.5 μg mL-1) at 125 volts for 5 h using a horizantal gel electrophoresis apparatus (Sigma Chemical Co. St. Louis, MO). The amplified products were visualized under UV light and photographed with a Gel documentary system. A 1kb ladder was used as a DNA standard to estimate the molecular weights of the amplified products.
Band scoring and phylogenetic analysis: For each individual primer, PCR amplified products were designated. Data were scored on the basis of the presence or absence of the amplified products. If the product is present in a genotype, it was scored as 1, if absent, it was designated as 0. Only the clearest and strongest bands were used for phylogenetic analysis. Reproducibility of the patterns was tested by running the reactions in duplicates or in triplicates. Neighbor joining and parsimony analyses were performed on the RAPD data using PAUP 4.0 b10 (Swofford, 2003). A heuristic search was performed by RANDOM stepwise addition (10,000 replicates) with tree bisection-reconstruction (TBR) branch swapping to find the most parsimonious tree. From the resulting trees, the majority rule consensus was calculated. Robustness and topology of the trees were evaluated using the Consistency Index (CI) to measure the level of homoplasy in the data (how cleanly the data fit the tree). The Retention Index (RI) was measured to indicate the amount of synapomorphy on the tree. Robustness and reliability of the resulting phylogenies were measured by bootstrap (1,000 replicates with full heuristic searches). The step wise genetic distance matrix which measures the relatedness between the species (Kimura, 1980) was computed between all species. The data were analyzed with a neighbor joining approach.
RESULTS
The RAPD technique was used to characterize the four Pistacia species and clarify the relationships between them. A total of 248 fragments was generated by 23 arbitrary sequence primers and 139 bands out of these were polymorphic at the inter-specific level. Figure 1 indicates a representive example of RAPD markers detected in the four Pistacia species using primers OPA-09 and OPB-01.
Data were analyzed using the PAUP program, to obtain cladograms that depict the likely relationships among the species.
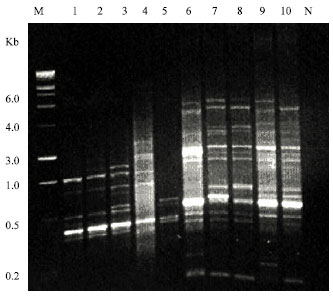 | |
| Fig. 1: | DNA amplification products of four Pistacia species. Lanes (1-5): RAPD patterns of DNA from P.vera, P. khinjuk, P. terebinthus, P. lentiscus and R. ovata, respectively using the primer OPA-09. Lanes (6-10): RAPD patterns of DNA from P. vera, P. khinjuk, P. terebinthus, P. lentiscus and R. ovata, respectively using the primer OPB-01. Lane M is a 1 Kbp molecular weight marker, lane N is a negative control (amplification product lacking DNA template sample) |
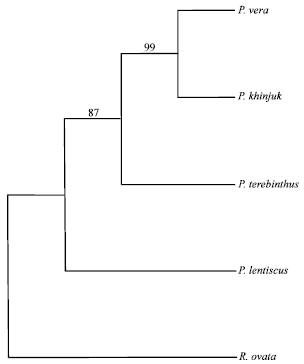 | |
| Fig. 2: | The majority rule consensus tree generated with RAPD data using parsimony analysis showing relationships of the Pistacia species with CI = 0.55 and RI = 0.67. Numbers above the branches indicate the bootstrap values |
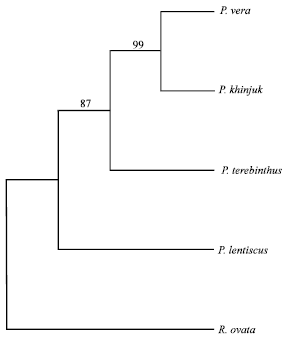 | |
| Fig. 3: | The majority rule consensus tree generated with RAPD data using neighbor joining analysis showing relationships of the Pistacia species. Numbers above the branches indicate the bootstrap values |
All the RAPD characters used were parsimony informative characters. Parsimony analysis produced one maximally parsimonious tree of 34 steps, with a Consistency Index (CI) of 0.55 and a Retention Index (RI) of 0.67 (Fig. 2). This tree was consistent with the tree produced by neighbor joining analysis (Fig. 3). There were insignificant differences between the bootstrap support values of the different branches of the two trees and the relationships among species within each clade. Pair-wise genetic distances between all the species were calculated by the same program.
According to these cladograms, the four Pistacia species are well separated from each other and the out group was well clustered from the Pistacia species. Two groups were resolved which were separated by a branch appearing in 87% of the bootstrap replicates in the parsimony analysis and 86% in the neighbor joining analysis. The first group includes P. khinjuk, P. terebinthus and P. vera while the second group comprises P. lentiscus. Within the first group, P. khinjuk and P. vera form a very close pair that is separated from P. terebinthus by a branch appearing in 99% of the bootstrap replicates in the parsimony analysis and 98% in the neighbor joining analysis.
DISCUSSION
The results of the current study support monophyly of the genus. Both analyses show that P. khinjuk and P. vera are very close species and P. khinjuk may be a descendant of P. vera. Two sections were revealed in the morphological study: section Pistacia which contains deciduous species and section Lentiscus which contains evergreen species. Two groups were obtained in the current study, the first group comprises P. khinjuk, P. terebinthus and P. vera (all species are deciduous trees and belong to section Pistacia) and the second group comprises one species (P. lentiscus) which is an evergreen shrub and belongs to section Lentiscus. The current study confirmed the placement of the three species of the first group in section Pistacia because they are genetically related and show a distant relationship with the other section represented by P. lentiscus.
Regarding the evolution of the four studied Pistacia species, Zohary (1952) hypothesized that P. vera is the most primitive species in the genus based on morphological characters (simple, odd-pinnate leaves, small number of leaflets per leaf, symmetrical leaflets, rounded leaflet apex, wingless petiole, highly branched panicles, deciduous character). Moreover, the very close relationship shown by the present study supports Zohary’s hypothesis that P. khinjuk may be a descendent of P. vera.
As indicated earlier in this study, Parfitt and Badenes (1997) performed a phylogenetic study of ten Pistacia species based on an analysis of the chloroplast genome. They found that the closest species to P. vera is P. khinjuk, followed by P. atlantica and P. terebinthus. They could not discriminate P. khinjuk from P. vera because the chloroplast genome is very conserved within the genus and they suggested that P. khinjuk and P. vera may be considered one species, despite the differences in morphology. Kafkas and Perl-Treves (2002) addressed the interspecific relationships in Pistacia based on RAPD fingerprinting. The following species were included: P. atlantica, P. eurycarpa, P. khinjuk, P. lentiscus, P. mexicana, P. palaestina, P. terebinthus and P. vera. Phylogenetic analysis yielded two main groups: P. atlantica, P. eurycarpa, P. khinjuk, P. integerrima and P. vera while P. lentiscus, P. mexicana, P. palaestina and P. terebinthus formed the second group. They showed again a close relationship between P. khinjuk and P. vera. The present study largely confirmed the relationships described by Parfitt and Badenes (1997) and Kafkas and Perl-Treves (2002).
ACKNOWLEDGMENTS
The authors gratefully acknowledge Sigma Xi, the Graduate Research Development Program at Virginia Polytechnic Institute and State University, the Southern Appalachian Botanical Society and the Virginia Academy of Science for funding this project.
REFERENCES
- Barazani, O., N. Dudai and A. Golan-Goldhirsh, 2003. Comparison of Mediterranean Pistacia lentiscus genotypes by random amplified polymorphic DNA, chemical and morphological analyses. J. Chem. Ecol., 29: 1939-1952.
Direct Link - Barone, E., L. Di Marco, F.P. Marra and M. Sidari, 1996. Isozymes and canonical discriminant analysis to identify pistachio (Pistacia vera L.) germplasm. Hortic. Sci., 31: 134-138.
Direct Link - Golan-Goldhirsh, A., O. Barazani, Z. Wang, D. Khadka, J. Saunders, V. Kostiukovsky and J. Rowland, 2004. Genetic relationships among Mediterranean Pistacia species evaluated by RAPD and AFLP markers. Plant Systematics Evol., 246: 9-18.
Direct Link - Hormaza, J.I., C. Dollo and V.S. Polito, 1994. Determination of relatedness and geographical movements of Pistacia vera (pistachio, Anacardiaceae) germplasm by RAPD analysis. Econ. Bot., 48: 349-358.
CrossRefDirect Link - Hormaza, J.I., K. Pinney and V.S. Polito, 1998. Genetic diversity of Pistachio (Pistacia vera, Anacardiaceae) germplasm based on randomly amplified polymorphic DNA (RAPD) markers. Econ. Bot., 52: 78-87.
CrossRefDirect Link - Kafkas, S. and R. Perl-Treves, 2001. Morphological and molecular phylogeny of Pistacia species in Turkey. Theor. Applied Gen., 102: 908-915.
CrossRef - Katsiotis, A., M. Hagidimitriou, A. Drossou, C. Pontikis and M. Loukas, 2003. Genetic relationships among species and cultivars of Pistacia using RAPDs and AFLPs. Euphytica, 132: 279-286.
Direct Link - Vos, P., R. Hogers, M. Bleeker, M. Reijans and T. van de Lee et al., 1995. AFLP: A new technique for DNA fingerprinting. Nucleic Acids Res., 23: 4407-4414.
CrossRefPubMedDirect Link - Warburton, M.L. and F.A. Bliss, 1996. Genetic diversity in peach (Prunus persica L. Batch) revealed by randomly amplified polymorphic DNA (RAPD) markers and compared to inbreeding coefficients. J. Am. Soc. Hortic. Sci., 121: 1012-1019.
Direct Link - Weining, S. and R.J. Henry, 1995. Molecular analysis of the DNA polymorphism of wild barley (Hordeum spontaneum) germplasm using the polymerase chain reaction. Genet. Res. Crop Evol., 42: 237-281.
CrossRef - Welsh, J. and M. McClelland, 1990. Fingerprinting genomes using PCR with arbitrary primers. Nucleic Acids Res., 18: 7213-7218.
CrossRefPubMedDirect Link - Williams, J.G.K., A.R. Kubelik, K.J. Livak, J.A. Rafalski and S.V. Tingey, 1990. DNA polymorphisms amplified by arbitrary primers are useful as genetic markers. Nucleic Acids Res., 18: 6531-6535.
CrossRefPubMedDirect Link - Parfitt, D.E. and M.L. Badenes, 1997. Phylogeny of the genus Pistacia as determined from analysis of the chloroplast genome. Proc. Natl. Acad. Sci. USA., 94: 7987-7992.
Direct Link - Kimura, M., 1980. A simple method for estimating evolutionary rates of base substitutions through comparative studies of nucleotide sequences. J. Mol. Evol., 16: 111-120.
CrossRefDirect Link