
Fatih Seyis
Not Available
Wolfgang Friedt
Not Available
Wilfried Luhs
Asian Journal of Plant Sciences
Year: 2005 | Volume: 4 | Issue: 1 | Page No.: 6-10







ABSTRACT
Brassica crop species have become one of the world wide most important source of vegetable and oils. This development was accomplished by substantial progress in breeding and biotechnology as well as by modernisation of cultivation practices. Brassica napus (2n=38, genome AACC) is a natural amphidiploid that originated from several independent spontaneous hybridisation events between the diploid species B. rapa (2n=20, AA) and B. oleracea (2n=18, CC). The limited geographic range of B. napus combined with intensive quality breeding has led to a narrow genetic basis in this species. In contrast, the original progenitors are both highly polymorphic and therefore, do offer a broad genetic variability that can be exploited for rapeseed improvement via wide hybridisation (resynthesis). Zero-erucic B. oleracea mutants (namely Kashirka and Ladozhskaya), which were identified as a novel source of a gene conferring low erucic acid content to Brassica seed oils, were crossed with the B. rapa cultivar `Asko` (0-Quality). Fatty acid analysis of seeds from individual hybrids revealed a zero-erucic acid phenotype as expected. This basic material can be used as a genetic resource for quality and yield improvement of oilseed rape.
PDF Abstract XML References Citation
How to cite this article
Fatih Seyis, Wolfgang Friedt and Wilfried Luhs, 2005. Development of Resynthesized Rapeseed (Brassica napus L.) Forms with Low Erucic Acid Content Through in ovulum Culture. Asian Journal of Plant Sciences, 4: 6-10.
DOI: 10.3923/ajps.2005.6.10
URL: https://scialert.net/abstract/?doi=ajps.2005.6.10
DOI: 10.3923/ajps.2005.6.10
URL: https://scialert.net/abstract/?doi=ajps.2005.6.10
INTRODUCTION
Developing synthetic Brassica napus forms has provided important basic germplasm for further improvements of seed yield, disease and pest resistance as well as relevant seed quality traits[1-3]. Regarding the later some examples of introgression breeding are documented, such as the resynthesis approach to create new genetic variation for low glucosinolate content[4] or the approaches developing of yellow-seeded rapeseed via interspecific crosses[5-8]. Different cauliflower cultivars (B. oleracea ssp. oleracea convar. botrytis Var. botrytis) and Yellow Sarson (B. rapa ssp. trilocularis) were used in an earlier study for wide hybridisation to create novel rapeseed with High Erucic Acid Content (HEAR) and which is now used as basic material for the improvement of industrial rapeseed[9-15]. Similar experiments were conducted[2,16-20] using interspecific Brassica hybridization in order to modify the fatty acid composition of oilseed rape.
In fact, current double-low breeding material seems to be closely related and intensive quality breeding-using ‘Liho‘/‘Oro‘ and ‘Bronowski‘ spring rapeseed as unique donors for quality improvement-has also contributed to narrow the genetic base of oilseed rape[21,22]. The possibility to create new B. napus forms from the interspecific crosses between B. rapa and B. oleracea, the so called resynthesized rapeseed forms are used for increasing the genetic variation in rapeseed (B. napus L.). In whole genebanks all over the world, there exists 20.106 accessions of B. oleracea and 18.224 accessions of B. rapa[23]. These high number of accessions in both progenitor species are describing the possibilities in creating new variation or new sources for B. napus.
In the course of studies dealing with the inheritance of erucic acid content in the seed oil of Brassica species[13,14] individual plants belonging to three accessions of B. oleracea conv. capitata, Kashirka 202, Ladozhskaya DS 8395 and Eisenkopf, were identified being very low in erucic acid content[24]. Because there was no information about the presence of cabbage genotypes conferring a low erucic acid content it was the first time that low-erucic acid mutants of B. oleracea have been described[24,25].
During the establishment of an appropriate in ovulum culture for producing interspecific rapeseed hybrids[26,27] a small number of interspecific hybrids with probably low erucic acid character were developed.
In this study zero-erucic B. oleracea mutants namely Kashirka and Eisenkopf were crossed with Asko (B. rapa) an 0+ quality type in order to develop resynthesized B. napus as novel genetic resource for both quality and yield improvement of oilseed rape.
MATERIALS AND METHODS
Plant material: The B. rapa cultivar Asko (0-Quality) and two cabbage accessions Kashirka 202 and Eisenkopf were used to develop resynthesized rapeseed forms with low erucic acid character. In every crossing combination the B. rapa genotype were used as pollinator. The fatty acid composition of the B. oleracea and B. rapa parents can be seen in Table 1. Unfortunately, only a small number of RS-lines were developed and evaluated phenotypically in the greenhouse.
Emasculation and pollination: For controlled pollination, buds which are about to open or will open the following day were selected for emasculation. Such flowers will not have been contaminated by self or foreign pollen. For emasculation of the flower buds we have used forceps. Because all the work of emasculation and pollination was done in the greenhouse sepals and petals should be removed together with anthers with no adverse effects. The pollination of the emasculated buds were done two days later, because the pollination and fertilization rate is more effective on this way in interspecific crosses. Pollen was collected from male plants and were transferred on the stigma of female plants and the emasculated flower buds were covered with plastic bags to prevent contamination with foreign pollen. 7-15 DAP (days after pollination) the immature pods were cutted at the moment when the ovules became obvious for preparing them using in in ovulum culture.
In ovulum culture: Immature pods were surface sterilized with NaHCl (5%) in 100 mL glasses. The washing procedure of these immature pods were done with sterile distilled water three times in the sterile bank. Then they were cutted with a scalpel during holding the pod with forceps under an binocular light microscope. The ovules were dissected and transferred to plastic petri dishes (7 cm in diameter) containing MS-medium with reduced sugar content (10 g L-1). The dishes were covered on the side with Nescofilm After germination of the ovules the plantlets were transferred on the same medium in Weck glasses.
After obtaining whole plants these were first transferred on M and S with normal sugar content (20 g L-1). At the time where its possible to make cuttings from these plants, they were transferred in Quick-Pots. Cuttings from these hybrids were treated with colchicine in order to obtain amphidiploid B. napus plants (°C) and artificially vernalised for 8 to 10 weeks at 5°C.
Fatty acid analysis: The fatty acid composition of the B. oleracea material was determined on the basis of half-seed analysis as described earlier[28,29] using GC. The preparation of half seeds was done according to the method described by Thies[30].
RESULTS
The presented data is the result of pre-experiments to develop RS-lines with quality characters. The parents Asko (Brassica campestris), Kashirka and Eisenkopf (Brassica oleracea) have nearly similar fatty acid amounts regarding oleic acid (C18:1), linoleic acid (C18:2) and erucic acid (C22:1) (Table 1).
The interspecific hybrids derived from the “Kashirka x Asko” cross are showing an range from 46.22 up to 63.11% regarding oleic acid (Table 1). Their erucic acid amounts are nearly zero. An ideal dietary plant oil contains approximately 60% oleic acid, high amounts of linoleic acid (>35%) and zero erucic acid[31]. The developed RS-lines (resynthesized rapeseed lines) show an fatty acid profile nearly to dietary oil regarding the given fatty acids.
The reason for the obtained limited seed number from the produced interspecific hybrids may be the self incompatibility character of these hybrids. İf self incompatible parents were used in interspecific crosses the produced hybrid possess the same character[32-34], which will give difficulties in obtaining seed with selfing.
Compared with other crop species rapeseed represents a relatively young crop plant. As a result the genetic variability in the gene pool of rapeseed seems to be intensively restricted with regard to important traits[29,35-38].
| Table 1: | Average fatty acid variation found in offspring lines (C1) derived from interspecific crosses between zero-erucic B. oleracea and B. rapa quality form (The description is corresponding for that every plant was developed from an different ovule)** |
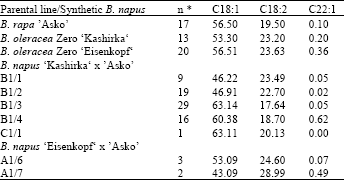 | |
| * Number of half seeds (parental genotypes) and seed samples (offspring lines) analysed, ** also B1/10 in the Kashirka x Asko cross and A1/1, A1/2 and A1/7 was developed, but because of no seed set of these hybrids they are not involved in Table 1. | |
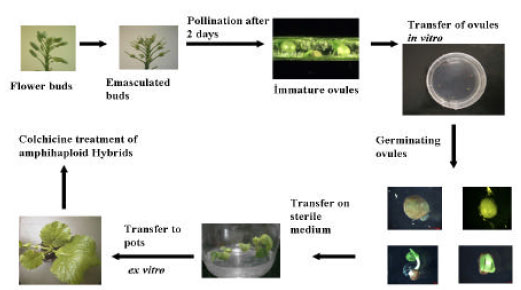 | |
| Fig. 1: | In ovulum method in producing interspecific rapeseed (Brassica napus) |
Resynthesized rapeseed forms are representing a very useful strategy to broaden the gene pool of rapeseed, because material produced on this way represents again a new gene pool compared with traditional rapeseed[29,37-41]. It’s very favourable, that for the generation of RS-forms a wide spectra of crossing parents are available in the diploid progenitor species, B. rapa and B. oleracea. The potential of resynthesizing B. napus is so far not totally exhausted, so that efforts, which are dealing with the maintaining and well directed use of genetic resources of the diploid parents will in future still became more important.
With the well directed selection of both the resynthesized rapeseed forms can be more or less predetermined. The examples Norde[42-47], Hakuran[48] and HEAR-rapeseed[29] are verifying this consideration. The newest development in the resynthesis of rapeseed are to develop them with regard to specific breeding objectives (for example disease resistance) or to use them in hybrid breeding.
The individual yield potential of resynthesized rapeseed is thereby low as expected. Therefore the use of generated RS-lines and consequently of the new generated variation lies with regard to the both character complexes quality and yield in the integration in high yielding breeding material[27,36]. This can be realized by developing so called semy-synthetic rapeseed forms, that is crossing resynthesized rapeseed with traditional rapeseed cultivars and lines, or with the development of test hybrids.
Therefore, with the discovery of low-erucic acid mutants[24-26,36] among the mentioned B. oleracea accessions and the Fig. 1 shown possible development of synthetic rapeseed forms via interspecific crosses with 00-quality B. rapa genotypes will open further the possibility to use RS-rapeseed material as a genetic resource for example for quality and yield improvement in Brassica napus, perhaps in developing RS-lines with 0- or 00-quality.
REFERENCES
- Seyis, F., W. Friedt, A. Voss and W. Luhs, 2004. Identification of individual Brassica oleracea plants with low erucic acid content. Asian J. Plant Sci., 3: 593-596.
CrossRefDirect Link - Seyis, F., R.J. Snowdon, W. Luhs and W. Friedt, 2003. Molecular characterization of novel resynthesized rapeseed (Brassica napus) lines and analysis of their genetic diversity in comparison with spring rapeseed cultivars. Plant Breed., 122: 473-478.
Direct Link - Luhs, W.W., A. Voss, J. Han, A. Grafin-zu-Munster and D. Weier et al., 1999. Genetic Modification of Erucic Acid Biosynthesis in Brassica napus. In: Genetics and Breeding for Crop Quality and Resistance Developments in Plant Breeding, Mugnozza, G.T.S., E. Porceddu and M.A. Pagnotta (Eds.). Vol. 8, Kluwer Academic Publ., Dordrecht, The Netherlands, pp: 323-330.