
Maria A. Perez-Fernandez
Not Available
M. Lopez-Martin
Not Available
R. Flores-Vargas
Not Available
E. Calvo-Magro
Not Available
G. O`Hara
Not Available
Asian Journal of Plant Sciences
Year: 2004 | Volume: 3 | Issue: 4 | Page No.: 532-538







ABSTRACT
Studies oriented to understanding the colonising potential of plant species rarely focus on the relationships between plants and other living organisms in the soil and in the rhizobacteria. This work presents part of the microbial diversity identified in eleven annual plant species common in crops under arid and semi arid climatic conditions (Hymenocarpos hispanicus Lassen, Trifolium arvense L., Trifolium clusii, Gardon and Gren., Ornithopus compressus L., Avena sterilis L., Briza maxima L., Bromus tectorum L., Holcus lanatus L., Lolium rigidum Gaudin, Poa bulbosa L. and Rumex bucephalophorus (Steinh)Reich.fil.). The effect of rhizobacteria on the germination and growth of two annual plant species, annual ryegrass (Lolium rigidum Gaudin) and wild radish (Raphanus raphanistrum L) was studied showing complex relations of competition and mutualism. From 20 bacterial isolates applied to seeds and seedlings of both species, five promoted seed germination and enhanced vegetative growth; seven isolates showed competitive effect inhibiting the germination and delaying plant growth; eight isolates had no significant effect neither in the germination nor in the total biomass production. These relations, besides contributing to the knowledge of vegetative dynamics present a potential tool of biological control of weeds, with the advantage of not introducing in the soil foreign (exotic) organisms of difficult control.
PDF Abstract XML References Citation
How to cite this article
Maria A. Perez-Fernandez, M. Lopez-Martin, R. Flores-Vargas, E. Calvo-Magro and G. O`Hara, 2004. Screening of Soil Micro-organisms and Their Influence in the Establishment of Annual Herbaceous Species. Asian Journal of Plant Sciences, 3: 532-538.
DOI: 10.3923/ajps.2004.532.538
URL: https://scialert.net/abstract/?doi=ajps.2004.532.538
DOI: 10.3923/ajps.2004.532.538
URL: https://scialert.net/abstract/?doi=ajps.2004.532.538
INTRODUCTION
Dynamics of Ecosystems are driven, among other factors by the number of species included and the interactions established between these species and between them and the environment. Most of the investigations have been primarily focused on the importance of species[1]. Recently, studies on diversity have taken a new dimension, by recognizing that numerous aspects of the structure of the community co-vary with species richness and simultaneously can have a greater control on the ecological processes than that exerted by the number of species per se[2]. The interactions in the ecosystems become more complicated when micro and macro-organisms are involved, like interactions involving plants and micro-organisms such as fungi and bacteria. The better-known interactions are derived from the positive micorrysation and symbioses between nitrogen fixing bacteria and their host plants. Nevertheless, the degree of interactions is much more complex, implying less well-known relations and not even described yet[3,4].
It is well known that in the plant rhizosphere, bacterial flora is abundant[5]. The positive and negative balance of the interactions between bacteria and their hosts will determine the colonisation capacity of a plant species[8]. Thus, the aim of the present study was to evaluate the effect of bacterial strains isolated from eleven plant species exerted on germination and early growth of annual ryegrass (Lolium rigidum Gaudin) and wild radish (Raphanus raphanistrum L.). These species were selected because of their wide range of distribution in numerous plant communities in the world and for being considered noxious weeds in crops. The interactions between bacteria and plants species were analysed and interpreted from two points of view (I) the effects on the colonisation capacity of the plant species and (ii) the potential of the deleterious rhizobacteria as biological control agents.
MATERIALS AND METHODS
Adult plants of the species were collected from natural populations in the central-western region of Spain (Table 1).
| Table 1: | Scientific names of the plant species from which bacterial isolates were extracted. Isolates names follow plant species' scientific names (the two first letters refer to the genus and the three following ones to the species name) |
 | |
Three soil samples from the rhizosphere (Soil A, Soil B and Soil C) were also collected. Plants and soil samples were stored in sterile plastic bags and stored at 4°C until they were processed.
Standard microbiological methods were used to isolate rhizobacteria from plant rhizosphere. Roots of the weed plants were cut into pieces of 3 cm length and transferred into 250 mL conical flasks with 40 mL of 0.1% (w/v) peptone water (1:40) and a few drops of Tween 80. Samples were shaked for two hours and the suspensions (1 mL) were plated onto selective (Sands and Rovira medium) and non-selective (tryptic soy and nutrient agar) media. Isolates were purified in culture and stored cryogenically (at -80°C). Similar procedure was used for bacterial extraction from the soil samples. The solutions were plated as above and the Petri dishes were checked daily, until growth of bacterial colonies.
Seeds of ryegrass and wild radish were collected from the same natural populations of adult plants of the species indicated in Table 1. Inoculation of sterile seeds in Petri dishes was carried out according to Vargas and O’Hara[5] protocol. One-day old cultures of each rhizobacterial isolate, grown in glucose minimum salt medium, were centrifuged at 20,000 rpm for 10 min and 2 mL of supernatant was added to the agar plates 0.9% (w/v). Four replicates of 20 surface-sterilised seeds of each weed species were then placed on each plate and incubated in the darkness at 200°C for five days. Controls used 2 mL of sterile medium. After five days the seedlings were removed, germination recorded and root lengths measured. Isolates that inhibited the target weed plants under laboratory conditions were further tested for their effects on plants grown under glasshouse conditions.
Sterile seeds of the two target species were germinated in Petri dishes on 0.9% agar. Two-day old seedlings of each species were transplanted into 110 mm pots containing sterile sand. Each seedling was inoculated with 2 mL of the supernatant extracted by centrifugation from bacterial cultures grown for 24 h. Selected bacterial isolates were those that had produced significant effects, positive or negative in root growth from previous experiment. Five replicates of four seedlings each were prepared for each combination of bacterial inoculant and plant species. Pots were covered with sterile polyethylene beads to prevent from contamination. Seedlings were grown over six weeks in a glasshouse at constant temperature of 25°C and photoperiod of 14 h light/10 h darkness. Plants were watered every two days with a nutrient solution supplemented with KNO3 (0.3%). After harvesting, roots were thoroughly washed with running water to eliminate any rest of sand. Individual shoot and root length was measured and dry biomass was estimated.
One-way ANOVA followed by multiple range test (test-T) for multiple comparisons was done for mean comparisons of plant growth.
RESULTS
Figure 1 shows the average length of seedlings of annual ryegrass and wild radish germinated in petri dishes after inoculation 20 with bacterial extracts. Significant differences were detected in seedling growth as a function of the inoculants (Table 2). Six of the inoculants applied to annual ryegrass (Hyhis 1, Trarv 1, Brtec 1, Brtec 2, Holan 1 and Rubuc) did not induce significant differences in seedling growth from that observed in the control; similarly, differences among these inoculants were not observed. The inoculants from Hyhis 2, Lorig, Trdub 1 and Trdub 2, Holan 2, Avste and Soil B, significantly promoted seedling growth. Inoculants from the soil (Soil B) and Trdub 1 induced the lowest growth, although still higher than that of the control. Inoculi Lorig, Trdub 1, Holan 2 and Avste induced intermediate values of seedling length similar between them and different from the others. The remaining inoculi, significantly reduced growth.
The behaviour of wild radish plants treated with the 20 bacterial inoculants was similar to that described for annual ryegrass although the bacterial inoculants that induced similar behaviour to the control (Fig. 1 and Table 2) were Brtec 1, Brtec 2, Trclu 1, Trclu 2 and Soil A. The isolates that significantly increased seedling growth were Trarv 1, Soil B, Lorig, Hyhis 1, Brmax 1, Holan 1 and Pobul.
| Table 2: | Results of the one-way ANOVAs. comparing length of seedlings germinated in petri dishes after inoculation with bacterial extracts from 20 different bacteria |
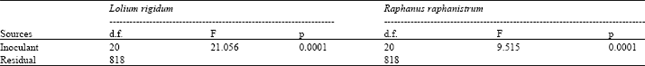 | |
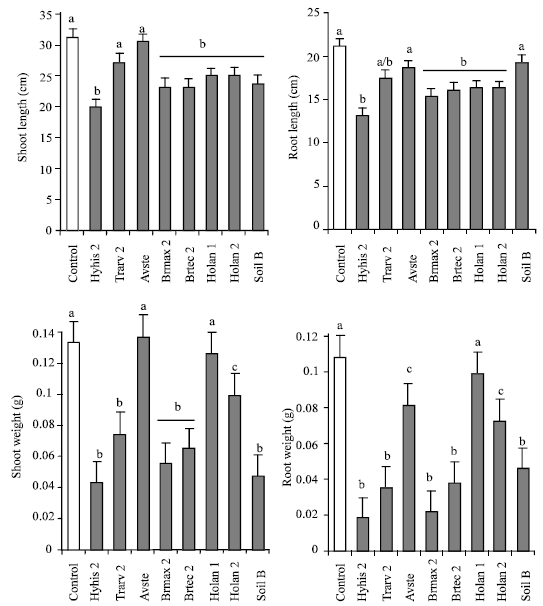
| Table 3: | Results of the one-way ANOVAs comparing shoot and root length and weight of seedlings of Raphanus raphanistrum grown under controlled conditions in the glasshouse after inoculation with the bacterial isolates from eight different bacteria |
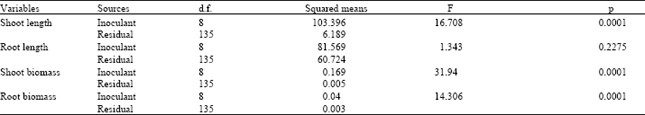 | |
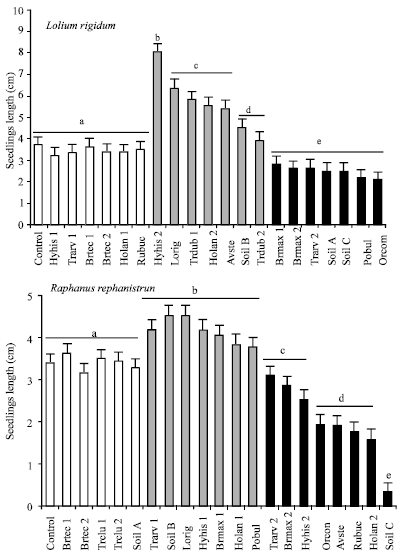 | |
| Fig. 1: | Average length of Lolium rigidum and Raphanus raphanistrum seedlings, germinated in petri dishes and inoculated with metabolites produced by 20 bacterial strains extracted from Hymenocarpos hispanicus (Hyhis), Ornithopus compressus (Orcom), Trifolium dubium (Trcdub), Trifolium arvense (Trarv), Avena sterilis (Avste), Briza maxima (Brmax), Bromus tectorum (Brtec), Lolium rigidum (Lorig), Holcus lanatus (Holan), Poa bulbosa (Pobul), Rumex bucephalophorus, as well as with three inoculants from soil samples(Soil A, Soil B and Soil C). Different letters on top of the columns indicate significant differences after ANOVA and T-test. Note different scales |
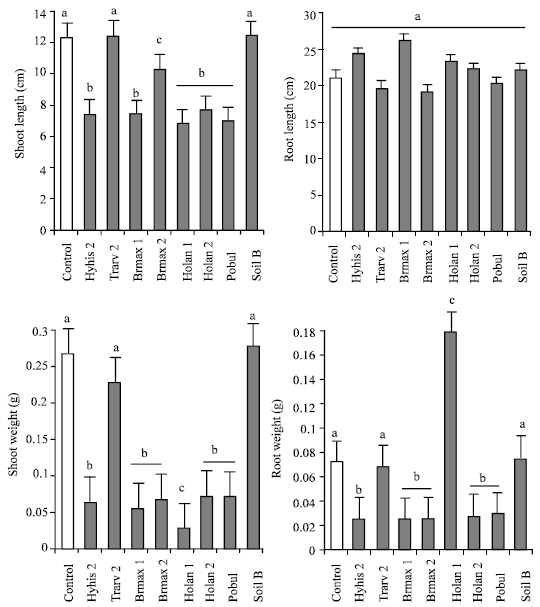 | |
| Fig. 2: | Average shoot and root length and weight of R. raphanistrum seedlings grown in sterile sand and inoculated with metabolites from eight bacterial strains extracted from Hymenocarpos hispanicus (Hyhis), Trifolium arvense (Trarv), Briza maxima (Brmax), Holcus lanatus (Holan), Poa bulbosa (Pobul), as well as one inoculant from a soil sample (Soil B). Different letters on top of the columns indicate significant differences after ANOVA and T-test. Note different scales |
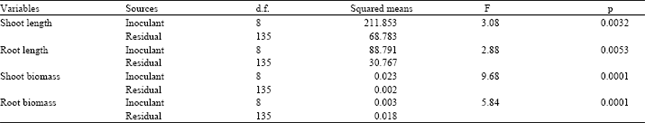
| Table 4: | Results of the one-way ANOVAs comparing shoot and root length and weight of seedlings of Lolium rigidum grown under controlled conditions in a glass house after inoculation with the bacterial isolates from eight different bacteria |
 | |
 | |
| Fig. 3: | Average shoot and root length and weight of Lolium rigidum seedlings grown in sterile sand and inoculated with metabolites from eight bacterial strains extracted from Hymenocarpos hispanicus (Hyhis), Trifolium arvense (Trarv), Avena sterilis (Avste), Briza maxima (Brmax), Bromus tectorum (Brtec), Holcus lanatus (Holan), Poa bulbosa (Pobul), as well as one inoculant from a soil sample (Soil B). Different letters on top of the columns indicate significant differences after ANOVA and T-test. Note different scales |
Finally, the isolates that significantly reduced seedling growth were Trarv 2, Brmax 2, Hyhis 2, Orcom, Avste, Rubuc, Holan 2 and Soil C.
None of the tested bacterial inoculants induced significant differences in root growth of wild radish (Table 3 and Fig. 2). However, significant differences were observed in shoot length and shoot and root biomass (Table 3). Both shoot length and biomass of wild radish were significantly inhibited by application of the inoculants Hyhis 2, Brmax 1, Brmax 2, Holan 1, Holan and Pobul, whereas the inoculants Trarv 2 and Soil B did not induce differences in plant growth with the control. Root biomass production in the presence of inoculants was similar to that described for shoot biomass, with the exception of inoculant Holan1.
As a rule, for annual ryegrass total seedling length (shoot+roots) and total shoot and root mass was significantly reduced in the presence of all bacterial inoculants (Table 4 and Fig. 3), with the exception of Trarv 2, Avste and Soil B. All inoculants significantly reduced seedling growth. The exceptions were Avste for shoot weight and Holan 1 for shoot and root weight (Table 4 and Fig. 3).
DISCUSSION
Soil and plant rhizospheres are colonised by bacterial strains whose metabolic exudates can exert positive and/or negative effects on the germination, growth and development of plant species[5]. Thus, together with the environmental factors, plant-micro-organisms interactions, like the ones described in this study, play a role in species germination and development.
Facts like many plant species becoming invaders, as most weeds, or species with a world-wide distribution (i.e. Fabaceae family), can easily be explained by the kind of bacteria they establish associations with[6-9]. Whenever infection by bacteria that activate plant growth takes place, the plant species involved in the interaction experience a greater growth and development than that in the absence of these beneficial bacteria. The predominance of beneficial bacteria over the deletereous can be due to a higher affinity of the plants by the bacteria with which they establish positive symbiose, than that with those bacteria with which plants establish negative interactions. Thus, bacterial infection of plants is the final result of a balance between the infection by one or another type of bacteria, (i.e. the result of a balance between one or another type of interaction). Under field conditions, competition between bacteria takes place, being those bacteria with better competitive capacity the ones that accessed the roots of the plants[10,11]. This explanation accounts for the fact that in the field the most evident and abundant plant-bacteria interaction is the positive (i.e. that inducing plant growth) rather than that inhibiting germination and plant growth. However, when the bacteria are isolated and individually tested under sterile conditions, the true effects of each species on plants can be appraised, otherwise masked by effects other than interaction itself.
In their natural environment, many micro-organisms multiply when the environmental availability of nutrients and conditions are favourable. During the exponential phase of growth, they use the nutrients available to produce the essential metabolites needed for growth and reproduction. Additionally the micro-organisms produce a variety of secondary metabolites, those that do not play a determining role in the growth, maintenance or reproduction of the organism, at the end of the phase of exponential growth and during the stationary phase[12,13]. Production of metabolites by antagonistic soil-borne pathogens is regulated as much by environmental signals as by genetic regulators and it is also induce by particular vegetal substances[7]. Whereas some secondary metabolites seem not to exert effect in the organisms which generate such metabolites, they are deletereous for other living beings. Some metabolites, produced by the same micro-organism have been demonstrated to exert different effects in different plants[14]. Indeed, this fact explains that inoculants like Hyhis 1, Trarv 1, Holan 1 and Rubuc do not induce any kind of seedling growth in annual ryegrass but induce greater growth (or its inhibition) in wild radish. Inoculants like Hyhis 2, Holan 2, Avste activate seedling growth in annual ryegrass and inhibit it in wild radish.
On the other hand, some inoculants independently of the species to which they are applied, induce similar plant behaviour. Brtec 1 and 2 neither activated nor inhibited root growth; Lorig and Soil B activate root growth; Brmax 2, Trarv 2, Orcom and Soil C, inhibited root growth. Those bacteria that constantly induce vegetative growth could be used as natural fertilizers; bacteria that constantly induce retardation in germination and growth as well as minor biomass production could be used as agents for biological control of weeds. In fact, in all the species inoculated with deletereous bacteria and evaluated in pots in the glasshouse experiment, a grade of growth symptoms has been identified that vary from the generalised retardation of the growth to several types of foliage chlorosis. In the two species inoculated with deletereous bacteria a poor lateral root development was appraised. These results are similar to others[5,15,16] which guarantees the existence of bacteria able to exert a strong negative effect on the growth and vegetative development. The way of performance of these bacteria is still unknown. However, the fact that none of them induced damage at cellular level but only growth retardation and minor biomass production, lend us to think that action mechanism is based on the production of secondary metabolites. These metabolites are released into the environment when making contact with plants[10,17].
In order that the micro-organisms can be used like agents for biological control, either to induce plant growth or to control non-desired species, it is still precise to make a good selection of organisms adapted for each specific purpose (fertiliser or for weed control); to discern what are the suitable environmental conditions for their application in the field; and to devise formulations with potential commercial use.
ACKNOWLEDGMENTS
This work was supported by grants from ADICOMT through project 'Diseño de protocolos para el control biológico de malas hierbas de cultivo (viñedos, olivos e higueras)' and from Spanish Ministry of Science and Technology, Project Ren2001-0749/GLO.
REFERENCES
- Tilman, D., 1999. The ecological consequences of changes in biodiversity: A research for general principles. Ecology, 80: 1455-1474.
CrossRef - Belanger, R.R., N. Dufour, J. Caron and N. Benhamou, 1995. Chronological events associated with the antagonistic properties of Trichoderma harzianum against Botrytis cinerea-indirect evidence for sequential role of antibiosis and parasitism. Biocontrol Sci. Technol., 5: 41-54.
Direct Link