
Besma Ben Dkhil
Institut Superieur Agronomique Chott-Mariem Sousse, 4042, Tunisie
Mounir Denden
Institut Superieur Agronomique Chott-Mariem Sousse, 4042, Tunisie
American Journal of Plant Physiology
Year: 2012 | Volume: 7 | Issue: 4 | Page No.: 174-183







ABSTRACT
To study the response of okra (Abelmoschus esculentus cv. Marsaouia) to salinity, the effect on seedling growth, water content, anthocyanins content and chlorophyll fluorescence was investigated. Okra seeds were germinated in Petri dishes and sown in peat culture under controlled conditions and submitted during 2 weeks to saline stress ranging from 0 (distilled water) to 20, 40, 60, 80 and 100 mmol of NaCl. Results showed that salt has no significant effect on aerial part dry weight and it showed no changes in chlorophyll fluorescence parameters (Fo and Fv/Fm). By contrast, root dry weight decreased with increasing NaCl concentration with a subsequent increase in root water content and cotyledons+hypocotyls/root ratio. In addition, anthocyanins content and electrolyte leakage were highly increased by exposure to high levels of salt especially 80 and 100 mmol NaCl.
PDF Abstract XML References Citation
Received: November 04, 2011;
Accepted: February 15, 2012;
Published: May 16, 2012
How to cite this article
Besma Ben Dkhil and Mounir Denden, 2012. Effect of Salt Stress on Growth, Anthocyanins, Membrane Permeability and Chlorophyll Fluorescence of Okra (Abelmoschus esculentus L.) Seedlings. American Journal of Plant Physiology, 7: 174-183.
DOI: 10.3923/ajpp.2012.174.183
URL: https://scialert.net/abstract/?doi=ajpp.2012.174.183
DOI: 10.3923/ajpp.2012.174.183
URL: https://scialert.net/abstract/?doi=ajpp.2012.174.183
INTRODUCTION
Among abiotic stresses, salinity is one of the most severe problems in worldwide agricultural production. Salinity changed plant morphological traits (Zadeh and Naeini, 2007) decreased plant dry matter, leaf area (Amirjani, 2010) and crop yield (Bybordi and Ebrahimian, 2011). Around 1/5 of the 2.8 billion hectares of arable land on earth are affected by salt and the problem is becoming an increasing threat to agriculture globally (Pessarakli and Szabolcs, 1999). Soil salinity, one of the most serious problems on planting areas, has the mast obstructive impact on crop production in the world. This crisis problem attracts many scientists to overcome this obstruction by improving salt-tolerant lines (Gholizadeh and Navabpour, 2011). In addition, there is an increasing trend for using saline water in agriculture (Alem et al., 2002) because of water scarcity, especially for vegetable crop production in many Mediterranean countries including Tunisia.
Most of cultivated plants are sensitive to salt stress, in which NaCl-salinity involves modification of morphological, physiological and biochemical processes and anatomical changes (Tester and Davenport, 2003). These changes usually result in reduction of shoot (Misra et al., 1995) and restricted rooting (Lopez and Satti, 1996). It has also been reported that the effect of excess salinity reduced transpiration rates (Massai et al., 2004), photosynthesis and pigment contents (Renault et al., 2001; Stoeva and Kaymakanova, 2008). Anthocyanins are a large class of water soluble pigments in the flavonoid group. Many works relate anthocyanin synthesis to the induction of some type of stress in plants but only few reports dealt with the increases of anthocyanins as a result of response to salt stress (Kaliamoorthy and Rao, 1994; Chalker-Scott, 1999).
The conductivity test, based on solute leakage, has been proposed as a good indicator of salt tolerance in plants (Ghoulam et al., 2002). The cell membrane often suffered injury associated with the increases in permeability and loss of integrity (Blokhina et al., 2003). The increase in electrolyte leakage was due to the loss of ability to reorganize cellular membranes rapidly and completely (McDonald, 1980).
On the other hand, chlorophyll fluorescence has been used by physiologists to evaluate response of various crop species to determine the influences of abiotic stresses at various stages of plant growth. It has proved particularly useful in salinity tolerance screening programs (Jimenez et al., 1997; Pak et al., 2009). Analysis of fluorescence characteristics such as the nature and intensity of the emission bands, quantum yield and induction kinetics, reflects the properties of the chlorophyll molecules and their interaction with the external environment and also with associated physiological processes (Hall and Rao, 1999). The photochemical yield Fv/Fm (where, Fv: variable fluorescence and Fm: maximal fluorescence) which reflects potential quantum yield of PSII indicates the physiological state of the photosynthetic apparatus in intact plant leaves (Pereira et al., 2004). Pospisil et al. (1998) mentioned that environmental stresses affect PSII efficiency and lower the Fv/Fm values.
Germination and seedling establishment constitute the most critical periods in the life cycle of plants (Sosebee and Wan, 1987). There are many criteria for determining the levels of salt tolerance of varieties and species. Ideally, they should be simple to measure and permit the identification of salinity tolerance during seed germination or at the seedling stage (Akinci et al., 2004). Adverse effects of salinity on seed germination and seedling growth as well some physiological activities of cultivated plant species have been extensively investigated (Bayuelo-Jimenez et al., 2002; Zafar et al., 2005, Hameed et al., 2008, Kaymakanova et al., 2009).
Okra belongs to family Malvaceae, originated in tropical Africa and was grown in Mediterranean region. In many areas of Tunisia, the productivity of this crop is limited because of the water and soil salinization. The improvement of this productivity requires understanding of the mechanisms of tolerance of this species to salinity. On the basis of previous experiments, Ben Dkhil and Denden (2010) have reported that okra seeds were less sensitive to salinity (below 100 mmol NaCl) at germination stage. Salt tolerance of plants varies during their successive growth stages; the present investigation was carried out to evaluate the morphological and physiological behavior of okra seedlings in saline conditions. For this purpose, we compiled the measurements of seedling growth with measurements of chlorophyll fluorescence, anthocyanins content and membrane permeability. Besides investigating the possible tolerance mechanisms during seedling stage under conditions of moderate and severe salt stress (20, 40, 60, 80 and 100 mmol NaCl).
MATERIALS AND METHODS
Seed material and germination: Seeds were collected from local plants of okra (Abelmoschus esculentus L., cv. Marsaouia); they have been provided by Baddar Company (Tunisia). Marsaouia is the most cultivated cultivar in Tunisia. Seeds were sterilized with 15% sodium hypochlorite for 15 min and washed thoroughly with distilled water. They were then placed to germinate in Petri dishes (9 mm diameter) containing two sheets of Whatman No.1 filter paper, saturated with distilled water (control) or NaCl solutions (20, 40, 60, 80 and 100 mmol NaCl) at 25°C in the dark.
Growth conditions: The emergence of radicle from seed was taken as an index of germination. Thus, germinated seeds in distilled water and in saline solutions were transferred in a plastic disposable containers filed with peat/perlite mixture (2:1 v/v) to emerge. Containers, where incubated in a growth chamber with controlled temperature adjusted at 25°C day/night (25°C was determined by Ben Dkhil and Denden (2010) as optimal temperature for germination of okra seeds under saline condition) and relative humidity maintained between 75-80% with 12 h photoperiod. The containers were irrigated continuously with distilled water or salt solution during two weeks. Treatments were replicated 4 times in a factorial experiment laid out in a completely randomized design.
Growth parameters: On day 15, the seedling stage was achieved with cotyledonary expansion and data were recorded. Twenty seedlings, representing each treatment, were harvested and washed with deionised water. Washed seedlings were separated into root, hypocotyls and cotyledons for the determination of Fresh Weight (FW), Dry Weight (DW) after drying the samples to a constant weight in a 80°C oven. The (hypocotyls+cotyledons) dry weight/root dry weight ratio were calculated out of these data. Further, cotyledons and hypocotyls (aerial part) water content and root water content was obtained from the (FW-DW)/DW ratio (Prado et al., 2000).
Anthocyanin determination: The method taken from Mancinelli (1990) was applied for the determination of anthocyanin content of okra seedlings. After being thoroughly extracted in 3 mL methanol-HCl (1% HCl, v/v), the samples were left at 4°C in the refrigerator for 2 days. Later on, the extract were vortexed, filtered and the total anthocyanin content was measured by an UV-visible spectrophotometer as the difference between the absorbance at 530 and 657 nm wavelength and placed in the A530-A657 formula to eliminate the chlorophyll content in the extract, defined quantitatively as OD 530 g-1 fresh weight.
Electrolyte leakage (membrane permeability): Electrolyte leakage was used to assess membrane permeability according to Lutts et al. (1995). Cotyledon samples were washed with distilled water to remove surface adhered electrolytes and cut into discs of uniform size. Cotyledon discs were put in closed test tubes containing 10 mL of deionized water and incubated at room temperature (25°C) for 24 h and subsequently electrical conductivity of the solution (EC1) was recorded. Samples were then autoclaved at 120°C for 20 min and the final electrical conductivity (EC2) was obtained after cooling the solution to room temperature. The Electrolyte Leakage (EL) was calculated as EC1/EC2 and expressed as percentage.
Chlorophyll fluorescence measurements: Chlorophyll fluorescence emission from the upper surface of the cotyledons of intact seedlings was measured by a portable fluorescence monitoring system (F.I.M, 1500, ADC, Fluorescence Induction Monitor 1550, Analytical Development Company Limited). Prior to fluorescence measurements, a circular surface of the upper face of the cotyledons were dark adapted for 20 min using the dark adaptation clips. The initial fluorescence (Fo), the maximum fluorescence value (Fm) were assessed. The variable fluorescence (Fv) was calculated as Fv = Fm-Fo and the maximum quantum efficiency of PSII photochemistry (Fv/Fm) was determined.
Statistical analysis: Data were analysed with SPSS for windows: version 13.0 (standard version) using one-way analysis of variance (ANOVA) procedure. Differences among the means were compared by Duncan’s test.
RESULTS
Seedling growth response: The physical growth parameters such as hypocotyls+cotyledons and root dry weights were measured under varying NaCl salinity stress levels (Table 1). Present results proved that aerial part (hypocotyls and cotyledons) dry weight did not vary whatever the NaCl treatments; it was in the range of 0.035 and 0.043 g, under (80 and 100 mmol NaCl) and (20, 40, 60 mmol NaCl), respectively. By contrast, the root dry weight was significantly reduced with increasing salt concentration, resulting in an increase on the shoot/root ratio. Indeed, salt treatment significantly affected shoot/root ratio (p<0.05), the 60 and 80 mmol treatments increased this ratio to 7.4 and 7.5, respectively. Moreover, the highest value of shoot/root ratio was registered in seedlings treated with 100 mmol NaCl, it was around 10. The magnitude of root growth reduction was highly dependent upon NaCl concentrations. The root dry weight was reduced by around 67% at the highest salt concentration (100 mmol NaCl) compared to control (Table 1).
In hypocotyls and cotyledons, water content gradually decreased with increasing salt concentration. As stress became more severe (60, 80 and 100 mmol NaCl) aerial part water content was considerably limited (respectively -35 and -39% with respect to control).
However, root water content significantly increased with increasing NaCl concentration, reaching values of 15.83 and 19.08 under 60 and 100 mmol NaCl, respectively (Table 2).
Anthocyanins content: Changes in anthocyanin amounts by NaCl concentrations of 15 days old seedlings are presented in Table 3. As can be shown by statistical analyses (p<0.05), anthocyanins content was significantly induced by salinity. With the increase in NaCl concentrations, there was a marked increase in anthocyanin amounts. For 100 mmol treatment, the value of anthocyanins content was 25 times higher than control one.
Membrane permeability: The extent of membrane damage was assessed indirectly by conductometric measurements of solute leakage from cells. The amount of electrolyte leakage increased under salt stress (Table 3).
| Table 1: | Variation of the aerial part (cotyledons+hypocotyls) dry weight (DW), root dry weight (RDW) and the aerial part/root ratio of okra variety subjected to different NaCl concentrations (0, 20, 40, 60, 80 and 100 mmol) during early seedling stage |
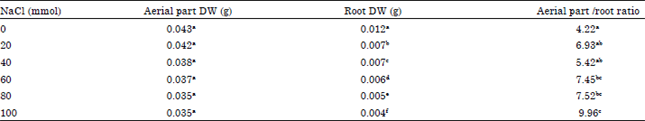 | |
| Values of each column followed by the same letter indicate no significant difference (p<0.05) according to Duncan’s test | |
| Table 2: | Variation of the water content in aerial part (cotyledons+hypocotyls) and root of okra variety subjected to different NaCl concentrations (0, 20, 40, 60, 80 and 100 mmol) during early seedling stage |
 | |
| Values of each column followed by the same letter indicate no significant difference (p<0.05) according to Duncan’s test | |
| Table 3: | Effects of NaCl treatments (0, 20, 40, 60, 80 and 100 mmol) on anthocyanins content (OD 530 g-1 fresh weight) and electrolyte leakage (%) at early seedling stage of okra variety |
 | |
| Values of each column followed by the same letter indicate no significant difference (p<0.05) according to Duncan’s test | |
| Table 4: | Effect of increasing NaCl concentration on fluorescence parameters in cotyledons of okra seedlings |
 | |
| Values of each column followed by the same letter indicate no significant difference (p<0.05) according to Duncan’s test | |
The magnitude of this increase was significant at 80 and 100 mmol NaCl. Electrolyte leakage was slightly modified by moderate salinity; it was ranged between 20.59 and 21.76% at 20 and 40 mmol NaCl, respectively.
Chlorophyll fluorescence: The effect of six levels of NaCl on chlorophyll fluorescence parameters were examined on dark-adapted cotyledons (Table 4). The initial level of fluorescence (Fo) didn’t change with NaCl level, it was not significantly affected by salinity stress in developing cotyledons. It was around 520 and 481, respectively at 0 and 60 mmol NaCl. Maximum level of fluorescence (Fm) was different in different salinity treatments than in control, 20 and 40 mmol NaCl, Fm being lower in salt treated seedlings with 60, 80 and 100 mmol NaCl. The Fv/Fm ratio was ranged between 0.79 and 0.82, demonstrating no significant effect of NaCl treatments on PSII photochemical activity.
DISCUSSION
In this work, salt stress inhibited significantly root growth, although, hypocotyl and cotyledons growth was not affected by the increasing NaCl concentrations. Our results were similar with the findings of Jamil et al. (2006) who suggested that salt stress inhibited the growth of root more than shoot in four vegetables species (cabbage, sugar beet, Paniculate amaranth and pak-choi). Similarly, Saha et al. (2010) found that roots of mungbean seedlings were more sensitive to salinity than shoots. Nevertheless, Maghsoudi and Maghsoudi (2008) reported that salt stress inhibited coleoptile growth more than root growth of wheat seedlings. Afzal et al. (2005) suggested that salinity affect seedling stage of wheat range from reduction in germination percentage, fresh and dry weight of shoots and roots to the uptake of various nutrient ions. Moreover, Cicek and Cakirlar (2002) reported that salinity reduced shoot length, fresh and dry weight of maize seedlings.
Reduced seedling growth under salt stress condition has also been reported by Huang and Redmann (1995) on barley, Foolad and Jones (1993) on tomato and Jeannette et al. (2002) on Phaseolus. In reviewing, the effect of salt stress on seedling growth found a reduction in root and shoot development that may be due to toxic effects of NaCl used as well as unbalanced nutrient uptake by the seedlings (Jamil et al., 2006). On the other hand, it was reported that decline in seedling dry weight in response to salinity is a consequence of decline in weight of mobilized seed reserve (Renault et al., 2001). Indeed, Jaleel et al. (2009) suggested that the reduction of plant growth under salt stress is the result of the alteration of many physiological activities in the plant, such as photosynthetic activity, mineral uptake and antioxidant activity. Amirjani (2010) reported that increasing salinity level to 100 and 200 mmol NaCl resulted in a reduction of plant height and fresh weight; an increase in proline content and diamine oxidase activity in soybean.
Salt stress also resulted in a significant increase in cotyledons+hypocotyls/root ratio which be considered as a direct effect of the observed roots dry mass reduction of treated seedlings (Table 1). These results contradict with Maghsoudi and Maghsoudi (2008) that salt stress inhibited coleoptile growth more than root growth. Rahman et al. (2008) reported that salinity depressed shoot growth more than root growth and increased root/shoot ratio.
According to the data obtained from the experiments, anthocyanin production increased markedly as salt stress level increased (Table 3). Anthocyanin accumulation that is known as a hallmark of plant stress was stimulated by increasing concentrations of NaCl in various parts of tomato and red cabbage seedlings (Eryilmaz, 2006). As reported by Winkel-Shirley (2002) anthocyanin synthesis is one of the subsequent production and its localization in root, stem and especially leaf tissues may allow the plant to develop resistance to a number of environmental stresses. Ali and Abbas (2003) found that saline (NaCl) stress in barley seedlings causes an increase in total phenolic compounds, flavonoids and enhancement of peroxidase and indoleacetic acid oxidase activities and consequent decrease in growth rate.
Therefore, increasing electrolyte leakage was used to assess membrane permeability. Addition of 100 mmol NaCl induced slightly membrane damage (Table 3). Kaya et al. (2001a) suggested that increases in membrane permeability at seedling stage were lower than at vegetative stage at high salinity and this also shows a strong link between time of exposure to high salinity and membrane permeability.
Salt stress induced electrolyte leakage has also been previously observed in maize (Hichem et al., 2009). Similar results were obtained in tomato by Kaya et al. (2001b) and in cucumber and pepper Kaya et al. (2001c).
Measurements of chlorophyll fluorescence parameters were attempted to evaluate the direct effects of salt stress on PSII photochemistry. Results showed that salinity level increased with no significant effect on Fv/Fm ratio, these results can be related to some findings in which it has been observed that salt stress has no effect on PSII photochemical activity, example in wheat (Raza et al., 2006). According to Zribi et al. (2009), the Fv/Fm ratio was not affected by any salt treatment of tomato seedlings (cv. Rio Grande). Similar results were reported by Jamil et al. (2007). They observed a lack of changes in the maximal efficiency of PSII photochemistry (Fv/Fm) in control and salt-stressed plants suggests that salt stress had no effects on PSII primary photochemistry in cabbage and sugar beet. It seems that the effects of salt stress on PSII photochemistry in plants depend on light intensity.
The results presented in this paper suggested that salt levels applied did not create high membrane damage and did not significantly affect the PSII activity. At seedling stage, NaCl levels ranged from 0 to 100 mmol might be low for okra. On the other hand, Ul-Haq (2009) reported that many genotypes of okra failed to germinate at 80 mmol NaCl, this concentration was found to be suitable for discriminating tolerant and non tolerant okra genotypes.
CONCLUSION
Some important conclusions can be drawn from the results achieved during this experiment. Okra is considered to be a salt tolerant species. Our results indicate its ability to maintain high physiological activities when subjected to relatively high levels of NaCl at early seedling stage. Therefore, comparisons of salinity tolerance mechanisms in different varieties will be fundamental to understand the regulatory points assigning physiological mechanisms that are likely to provide salinity tolerance.
REFERENCES
- Afzal, I., S.M.A. Basra, N. Ahmad and M. Farooq, 2005. Optimization of hormonal priming techniques for alleviation of salinity stress in wheat (Triticum aestivum L.). Caderno Pesquisa Serie Biologia, 17: 95-109.
Direct Link - Akinci, I.E. S. Akinci, K. Yilmaz and H. Dikici, 2004. Response of eggplant varieties (Solanum melongena) to salinity in germination and seedling stages. N. Z. J. Crop Hortic. Sci., 32: 193-200.
CrossRefDirect Link - Ali, R.M. and H.M. Abbas, 2003. Response of salt stressed barley seedlings to phenylurea. Plant Soil Environ., 49: 158-162.
Direct Link - Amirjani, M.R., 2010. Effect of salinity stress on growth, mineral composition, proline content, antioxidant enzymes of soybean. Am. J. Plant Physiol., 5: 350-360.
CrossRefDirect Link - Blokhina, O., E. Virolainen and K.V. Fagerstedt, 2003. Antioxidants, oxidative damage and oxygen deprivation stress: A review. Ann. Bot., 91: 179-194.
CrossRefPubMedDirect Link - Bybordi, A. and E. Ebrahimian, 2011. Effect of salinity stress on activity of enzymes involved in nitrogen and phosphorous metabolism case study: Canola (Brassica napus L.). Asian J. Agric. Res., 5: 208-214.
CrossRef - Chalker-Scott, L., 1999. Environmental significance of anthocyanins in plant stress responses. Photochem. Photobiol., 70: 1-9.
CrossRef - Cicek, N. and H. Cakirlar, 2002. The effect of salinity on some physiological parameters in two maize cultivars. BULG. J. Plant Physiol., 28: 66-74.
Direct Link - Eryilmaz, F., 2006. The relationships between salt stress and anthocyanin content in higher plants. Biotechnol. Biotechnol. Equip., 20: 47-52.
CrossRefDirect Link - Gholizadeh, F. and S. Navabpour, 2011. Effect of salinity on morphological and physiological characteristics in correlation to selection of salt tolerance in rice (Oryza sativa L.). Int. J. Agric. Res., 6: 780-788.
CrossRefDirect Link - Ghoulam, C., A. Foursy and K. Fares, 2002. Effects of salt stress on growth, inorganic ions and proline accumulation in relation to osmotic adjustment in five sugar beet cultivars. Environ. Exp. Bot., 47: 39-50.
CrossRefDirect Link - Hichem, H., D. Mounir and E. Naceur, 2009. Differential responses of two maize (Zea mays L.) varieties to salt stress: Changes on polyphenols composition of foliage and oxidative damages. Ind. Crop Prod., 30: 144-151.
CrossRef - Hameed, A., S. Naseer, T. Iqbal, H. Syed and M.A. Haq, 2008. Effects of NaCl salinity on seedling growth, senescence, catalase and protease activities in two wheat genotypes differing in salt tolerance. Pak. J. Bot., 40: 1043-1051.
Direct Link - Huang, J. and R.E. Redmann, 1995. Saft tolerance of Hordeum and Brassica species during germination and early seedling growth. Can. J. Plant Sci., 75: 815-819.
Direct Link - Jaleel, C.A., R. Gopi and R. Panneerselvam, 2009. Alterations in non-enzymatic antioxidant components of Catharanthus roseus exposed to paclobutrazol, gibberellic acid and Pseudomonas fluorescens. Plant Omics J., 2: 30-40.
Direct Link - Jamil, M., L. Deog Bae, J. Kwang Yong, M. Ashraf, L. Sheong Chun and R. Eui Shik, 2006. Effect of salt (NaCl) stress on germination and early seedling growth of four vegetables species. J. Cent. Eur. Agric., 7: 273-282.
Direct Link - Jimenez, M.S., A.M. Gonzalez-Rodriguez, D. Morales, M.C. Cid, A.R. Socorro and M. Caballero, 1997. Evaluation of chlorophyll fluorescence as a tool for salt stress detection in roses. Photosynthetica, 33: 291-301.
CrossRef - Kaya, C., H. Kirnak and D. Higgs, 2001. Enhancement of growth and normal growth parameters by foliar application of potassium and phosphorus in tomato cultivars grown at high (NaCl) salinity. J. Plant Nutr., 24: 357-367.
CrossRefDirect Link - Kaya, C., H. Kirnak and D. Higgs, 2001. Effects of supplementary potassium and phosphorus on physiological development and mineral nutrition of cucumber and pepper cultivars grown at high salinity (NaCl). J. Plant Nutr., 24: 1457-1471.
Direct Link - Lopez, M.V. and S.M.E. Satti, 1996. Calcium and potassium-enhanced growth and yield of tomato under sodium chloride stress. Plant Sci., 114: 19-27.
CrossRefDirect Link - Lutts, S., J.M. Kinet and J. Bouharmont, 1995. Changes in plant response to NaCl during development of rice (Oryza sativa L.) varieties differing in salinity resistance. J. Exp. Bot., 46: 1843-1852.
Direct Link - Mancinelli, A.L., 1990. Interaction between light quality and light quantity in the photoregulation of anthocyanin production. Plant Physiol., 92: 1191-1195.
Direct Link - Misra, M., N. Das and A.N. Misra, 1995. Sodium chloride salt stress induced changes in protein content and protease activity in callus culture of pearl millet (Pennisetum glaucum L. R. br.). Acta Physiol. Plant, 17: 371-374.
Direct Link - Maghsoudi, A. and K. Maghsoudi, 2008. Salt stress effects on respiration and growth of germinated seeds of different wheat (Triticum aestivum L.) cultivars. World J. Agric. Sci., 4: 351-358.
Direct Link - Pospisil, S., J. Kopecky and V. Prikrylova, 1998. Derepression and altered feedback regulation of valine biosynthetic pathway in analogueresistant mutants of Streptomyces cinnamonensis resulting in 2-ketoisovalerate excretion. J. Applied Microbiol., 85: 9-16.
Direct Link - Prado, F.E., C. Boero, M. Gallardo and J.A. Gonzalez, 2000. Effect of NaCl on germination, growth and soluble sugar content in Chenopodium quinoa willd. seeds. Bot. Bull. Acad. Sin., 41: 27-34.
Direct Link - Mujeeb-ur-Rahman, U.A. Soomro, M. Zahoor-ul-Haq and S. Gul, 2008. Effects of NaCl salinity on wheat (Triticum aestivum L.) cultivars. World J. Agric. Sci., 4: 398-403.
Direct Link - Raza, S.H., H.R. Athar and M. Ashraf, 2006. Influence of exogenously applied glycinebetaine on the photosynthetic capacity of two differently adapted wheat cultivars under salt stress. Pak. J. Bot., 38: 341-351.
Direct Link - Renault, S., C. Croser, J.A. Franklin and J.J. Zwiazek, 2001. Effect of NaCl and Na2So4 on red-osier dogwood (Cornus stolonifera Michx). Plant Soil, 233: 261-268.
Direct Link - Stoeva, N. and M. Kaymakanova, 2008. Effect of salt stress on the growth and photosynthesis rate of bean plants (Phaseolus vulg L.). J. Central Eur. Agric., 9: 385-391.
Direct Link - Tester, M. and R. Davenport, 2003. Na+ tolerance and Na+ transport in higher plants. Ann. Bot., 91: 503-527.
CrossRefDirect Link - Winkel-Shirley, B., 2002. Biosynthesis of flavonoids and effects of stress. Curr. Opin. Plant Biol., 5: 218-233.
CrossRefDirect Link - Zadeh, H.M. and M.B. Naeini, 2007. Effects of salinity stress on the morphology and yield of two cultivars of canola (Brassica napus L.). J. Agron., 6: 409-414.
CrossRefDirect Link - Zafar, S., M.Y, Ashraf and M. Ashraf, 2005. Protease activity and associated changes during germination and early seedling stages of cotton grown under saline conditions. Int. J. Bot., 1: 103-107.
CrossRefDirect Link - Zribi, L., G. Fatma, R. Fatma, R. Salwa, N. Hassan and R.M. Nejib, 2009. Application of chlorophyll fluorescence for the diagnosis of salt stress in tomato Solanum lycopersicum (variety Rio Grande). Sci. Hortic., 120: 367-372.
CrossRef - Pak, V.A., M. Nabipour and M. Meskarbashee, 2009. Effect of salt stress on chlorophyll content, fluorescence, Na + and K + ions content in rape plants (Brassica napus L.). Asian J. Agric. Res., 3: 28-37.
CrossRefDirect Link - Foolad, M.R. and R.A. Jones, 1993. Mapping salt tolerance genes in tomato (Lycopersicon esculentum) using trait-based marker analysis. Theor. Applied. Gen., 87: 184-192.
CrossRef