
Ismanizan Ismail
School of Biosciences and Biotechnology, Faculty of Science and Technology, Universiti Kebangsaan Malaysia, 43600, Bangi, Selangor, Malaysia
Norazreen Abdul Rahman
Rubber Research Institute, Malaysia Rubber Board, 4700, Sungai Buloh, Selangor Darul Ehsan, Malaysia
Chan Kok Fei
School of Biosciences and Biotechnology, Faculty of Science and Technology, Universiti Kebangsaan Malaysia, 43600, Bangi, Selangor, Malaysia
Zamri Zainal
School of Biosciences and Biotechnology, Faculty of Science and Technology, Universiti Kebangsaan Malaysia, 43600, Bangi, Selangor, Malaysia
Nik Marzuki Sidik
School of Biosciences and Biotechnology, Faculty of Science and Technology, Universiti Kebangsaan Malaysia, 43600, Bangi, Selangor, Malaysia
Che Radziah Che Mohd Zain
School of Biosciences and Biotechnology, Faculty of Science and Technology, Universiti Kebangsaan Malaysia, 43600, Bangi, Selangor, Malaysia
American Journal of Plant Physiology
Year: 2009 | Volume: 4 | Issue: 1 | Page No.: 24-37







ABSTRACT
This research aimed to evaluate the specificity of sesquiterpene synthase promoter (SesqPro) activity in the oil palm tissues and tomato hairy roots and to determine the functional region of the promoter. The effect of jasmonic acid (JA) on the promoter activation and gene expression was also analyzed. A series of 5’ sequence deletions on the full-length SesqPro were generated and individually cloned into the pCAMBIA 1301 vector. Functional analysis was carried out on leaves, mesocarp slices and Immature Embryos (IE) of oil palm and tomato hairy roots that had been transformed with full-length SesqPro (PSPr-VF6). GUS expression was found in all the tissues and a higher activity was detected in IE and mesocarp slices. All the constructed derivatives of SesqPro were transformed into IE and mesocarp slices in order to determine the promoter regions which are responsible for gene expression. The reduction of GUS activity was found to be related to the removal of DNA sequences within the promoter region. The promoter was induced by the elicitor molecule JA, thus suggesting the presence of JA responsive elements within the promoter. Incubation with 100 μM of JA showed higher GUS activity in IE and mesocarp slices that had been transformed with PSPr-VF4 to PSPr-VF6. Nevertheless, the GUS activity was drastically reduced in IE and mesocarp slices containing the PSPr-VF3 promoter, suggesting that the presence of the G/A hybrid box located at -622 to -617 act as a specific element in response to elicitors. This study has shown that the action of SesqPro is non-specific and was influenced by JA induction.
PDF Abstract XML References Citation
How to cite this article
Ismanizan Ismail, Norazreen Abdul Rahman, Chan Kok Fei, Zamri Zainal, Nik Marzuki Sidik and Che Radziah Che Mohd Zain, 2009. Functional Analysis of the Elaeis oleifera Sesquiterpene SynthasePromoter Reveals Non-Specific Tissue Expression and Activation under Stress Condition. American Journal of Plant Physiology, 4: 24-37.
DOI: 10.3923/ajpp.2009.24.37
URL: https://scialert.net/abstract/?doi=ajpp.2009.24.37
DOI: 10.3923/ajpp.2009.24.37
URL: https://scialert.net/abstract/?doi=ajpp.2009.24.37
INTRODUCTION
Plants produce a variety of secondary metabolites which are not specifically used for plant growth and development and instead play an important role in ecological functions for communication and defense. Terpenoids contribute to multiple plant biochemical functions, such as the formation of quinine; the electron transport chain, as components of membranes (prenillipid in archaeabacteria and sterol in eubacteria and eukaryotes); regulation and subcellular targeting (protein prenilation); photosynthesis, as a pigment component (carotenoids, chlorophyll side chains); hormones (gibberellins, brassinosteroid, abscisic acid); and as plant defense compounds (monoterpene, sesquiterpene, diterpene) (Lange et al., 2001). Terpenoid biosynthesis occurs in two kinds of metabolic pathways: the mevalonate pathway and the non-mevalonate pathways. Isopentenyl diphosphate (IPP) is produced indirectly via the mevalonate pathway that accumulates in the cytoplasm whereas dimethyl diphosphate is an IPP isomer which accumulates inside the plastid via a non-mevalonate pathway.
Interestingly, sesquiterpene is only found in plants, microbes and insects because this compound plays an important role as the intermediate for the interaction between organisms and their environment (Kessler and Baldwin, 2001). New research has shown that sesquiterpene is important as a chemo-preventive agent in colon and skin cancers (Picaud et al., 2006). The primary step in sesquiterpene biosynthesis involves a group of enzymes which are known as sesquiterpene synthases. Sesquiterpene synthases are a protein family which are expressed ubiquitously and are able to convert the acyclic universal precursor farnesyl diphosphate (FPP) into more than 300 different sesquiterpene skeletons (Picaud et al., 2006).
A sesquiterpene synthase will catalyze the conversion of sesquiterpene compounds in various plant species according to the demand and the environment. For instance, an acyclic sesquiterpenoid such as farnesene and farnesol is produced by (E)-β-farnesene synthase, whereas bisabolene synthase will catalyze the conversion of bisabolene sesquiterpenoid. Germacrene and aristolechene sesquiterpenoids are catalyzed by germacrene synthase and aristolechene synthase, respectively. Germacrene synthase consists of a few isomers that exist naturally and are known as germacrene A, B, C and D. The conserved amino acid sequences, intron organization and exon size, which are almost similar in more than three dozen monoterpene, diterpenes and sesquiterpene synthases from plants, suggest that these synthases come from the same evolutionary origin (Trapp and Croteau, 2001). Consequently, physical-based homology bioinformatics screening methods had successfully been used to identify new plant terpene synthase genes. Nevertheless, there are significant differences in the primary structures of the sesquiterpene synthases of plants and microbes, while the crystal structure of four sesquiterpene synthases/cyclases from different biological ancestors shows that it is highly conserved in its three dimensional structure (Caruthers et al., 2000).
There are many studies on the uniqueness and diversity of sesquiterpene synthase, including its genetic organization, gene regulation, biochemical properties, phylogenetics and phytochemical properties. The primary study on sesquiterpene synthase was conducted on aristolechene synthase and epi-aristolochene synthase from fungi (Penicillium roqueforti) and Nicotiana tabaccum respectively (Proctor and Hohn, 1993). Earlier molecular biology studies on sesquiterpene synthase only involved cDNA clone characterization which encodes for enzyme activities in certain sesquiterpene biosyntheses (Facchini and Chappell, 1992) and its role in plant genomes. All sesquiterpene synthases from plants will be channeled into the cytoplasm (Back and Chappell, 1995; Chen et al., 1996) because there is no specific plastid sequence (Chappell, 1995). Kappers et al. (2005) has proven that cytoplasm sesquiterpene could also be produced in mitochondria due to the availability of farnesyl diphosphate.
Induced sesquiterpenes can also form secondary chemical defenses towards herbivorous insects and pathogen fungi which differ from the primary defense compounds that were produced from induced monoterpenes (Bohlmann et al., 1998). Unlike the relationship between sesquiterpene and antimicrobe activities, this compound is also involved in plant reproductive systems by protecting against any attacks of insects, bacteria and fungi in vivo. In Arabidopsis, an exclusively expressed sesquiterpene synthase is found in its flower which catalyzes a wide variety of sesquiterpenoids in that organ (Dorothea et al., 2005). Zea mays also produces some volatile sesquiterpenoids when it is attacked by Spodoptera littoralis and herbivores (Schnee et al., 2002).
There are also many studies conducted on plant responses to pathogens and abiotic factors involving the accumulation of antimicrobes phytoalexins. Similar to phytoalexin classes in other plant families, sesquiterpene phytoalexins could not be detected in healthy or control plants and instead are accumulated in response to elicitors or pathogens (Yin et al., 1997). Phytoalexins will only accumulate in cell cultures of tobacco cells (Nicotiana tabaccum) and Capsicum annum when these cells are being challenged with elicitors from pathogens such as cellulose, fungi cell wall hydrolysates and cryptogein, which is an extracellular protein from Phytophthora cryptogea (Milat et al., 1991). Facchini and Chappell (1992) had reported that there are 12-15 copies of sesquiterpene synthase found in the tobacco genome. It is also possible that this gene exists in a complex of genes which are controlled differently during development or are regulated in response to the environment.
Sesquiterpene synthase of E. oleifera has been previously isolated (Cha, 2001) and its gene expression is very unique because it is expressed continuously in specific mesocarp tissues. Thus, the aims of the work was to study the promoter specificity and strength as well as to recognize which elements are essential for the regulation of sesquiterpene synthase gene expression. The construction of the promoter deletion series, its transformation into oil palm tissues and tomato hairy roots and its subsequent treatment with JA as reported in this paper will help to identify the promoter region involved in the activation of the sesquiterpene synthase gene towards this elicitor.
MATERIALS AND METHODS
Plant Material and Growth Condition
The experiments were carried at the Plant Biotechnology Center, Institute of System Biology, Universiti Kebangsaan Malaysia, Bangi, Selangor, Malaysia. The IE and mesocarp slices used in this experiment were taken from oil palm fruits aged between 12-15 WAA (week after anthesis) and zero frond leaves of oil palms (unopened). Oil palm tissues were pre-cultured on an N6 OPS medium (Chu et al., 1975) at 28°C in the dark for 7-12 days before being subjected to transformation. Tomato seeds were cultured on an MS medium (Murashige and Skoog, 1962) for 14 days at 28°C under a photoperiod of 16:8 h (light:dark) to harvest the cotyledons before being using these in A. tumefaciens- and A. rhizogenes-mediated transformation.
Generation of Promoter Deletion-GUS Constructs
The pCAMBIA-Sesq-Pro (PSP-VF) plasmids each containing the full length and deleted sesquirtepene promoters were extracted from Escherichia coli strain DH5α as described by Birnboim and Doly (1979). Each of these plasmids were then digested with NcoI dan HindIII to remove the 35S CaMV promoter sequence. The resulting PSP-VF plasmid constructs which now lack the 35S CaMV promoter then were purified using a Qiagen gel purification kit. The fragments were made blunt-ended using Klenow enzymes and were purified by phenol:chloroform extraction. The linearized plasmids were then ligated at 6°C overnight and transformed into E. coli strain DH5α where 50 mg L-1 of kanamycin was used as the selection antibiotic. These new promoter constructs were designated as PSPr-VF1 to PSPr-VF6 with varying lengths of sesquiterpene synthase promoter sequences as shown in Fig. 1. The analysis of the series of promoter deletions was carried out by confirming the size of each construct using PCR with specific primers and digestion with EcoRI and PstI (data not shown).
Plant Transformation and Growth Condition
The serial deletions of promoter/GUS constructs were introduced into A. tumefaciens strain LBA 4404 and A. rhizogenes strain A4 and subsequently were allowed to grow on LB-Glucose (5 mM) supplemented with kanamycin (50 mg L-1) and streptomycin (100 mg L-1) and LB-Glucose (5 mM) supplemented with kanamycin (50 mg L-1), streptomycin (500 mg L-1) and rifampicin (100 mg L-1), respectively. Both strains were cultured for 2 days at 28°C and were then recultured to reach an OD600 reading of 0.50.
For the transformation of tomato seedlings using A. rhizogenes, the cotyledons were cut into 1 cm2 pieces and were then pre-cultured for 2 days. Subsequently, they were incubated into a solution containing an A. rhizogenes culture for 30 min and shaken gently before being transferred to a sterile filter paper to remove excess bacteria.
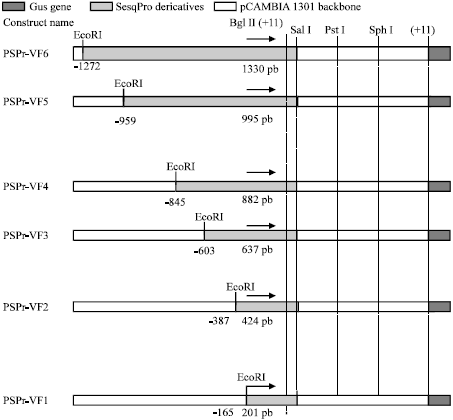 | |
| Fig. 1: | Restriction map of constructs series (pCAMBIA1301, SesqPro, r-EcoRI recombinant and VF1-6 represent insertions of deletion derivatives). SesqPro derivatives were cloned at EcoRI and SalI sites in the pCAMBIA 1301 vector. Arrows indicate the orientation of the promoter. Restriction enzyme SalI, PstI, SphI sites are located downstream of the promoter. The size for deletion derivatives of SesqPro are indicated in the map. SesqPro contains one site of BglII at position +11. The underlined number represents the actual size of the deletion derivatives of the SesqPro sequence |
After 2 days of co-cultivation at 28°C in the dark, the infected explants were transferred to an MS medium supplemented with 400 mg L-1 of cefotaxime (to inhibit bacterial growth). The transformed explants were incubated at 28°C with a photoperiod of 18:6 h (light:dark). Hairy roots that emerged from the cotyledons were subjected to selection by being transferring onto a new MS medium supplemented with 100 μg mL-1 of kanamycin.
The pre-cultured oil palm tissues (IE, mesocarp slices, leaf slices) were transformed using A. tumefaciens and were shaken gently for 30 min in an N6 6 medium (Chu et al., 1975) before being transferred to a sterile filter paper to remove excess bacteria for a 3 day co-cultivation in the dark at 28°C. The transformed tissues were then transferred to an N6 OPS medium and cultured at 28°C in the dark. These tissues were used for transient analysis after 10-15 days of culturing.
Treatment with Exogenous Jasmonic Acid
To determine the promoter strength under environmental stress, the transformed oil palm (mesocarp slices and IE) tissues were treated with 100 μM JA overnight in an N60 (modified Chu et al., 1975) medium at 28°C. The treated tissues were harvested and analyzed using GUS histochemical and fluorometric assays.
GUS Assays
GUS histochemical assays were performed 2 weeks after co-cultivation according to the method described by Jefferson et al. (1987). Transgenic explants were incubated in the GUS reaction buffer at 37°C for 18-24 h. To remove chlorophylls and pigments, the explants were treated with formalin-aceto-alcohol (FAA) solution (42.5% ethanol, 42.5% glacial acetic acid, 8.5% formalin). GUS expression was studied under a Nikon light stereomicroscope.
For fluorometric assay, FluoroAce (BioRad) was used. Various tissues of hairy roots and oil palm were homogenized in the GUS extraction buffer (50 mM natrium phosphate, 10 mM Dithiothreitol [DTT], 1 mM EDTA, 0.1% (w/v) sodium lauryl sarcosine and 0.1% (v/v) triton X-100). The homogenate was then centrifuged for 10 min at 10,000 rpm at 4°C and the GUS activity of the supernatant was assessed as described by Jefferson et al. (1987). Aliquots of extracts (30 μg) were added to 500 μL of assay buffer (5 mM MUG (methylumbelliferyl-β-D-glucuronide) in GUS extraction buffer ), pre-warmed and incubated at 37°C. After 30 min of incubation, the reaction mixtures were removed and placed in 1 mL of stop buffer (0.2 M Na2CO3, pH 11.2). Fluorescence was measured using a FluoroAce (BioRad). The protein concentration of the samples was determined using the procedure of Bradford (1976).
RESULTS AND DISCUSSION
Sesquiterpene Synthase Promoter (SesqPro) and Its Deletion Derivatives
The sesquiterpene synthase promoter (SesqPro) and its promoter deletion derivatives was isolated, analyzed and were subsequently constructed and inserted into the pCAMBIA 1301 vector (Cha, 2001). The 3’ end and the 5’ end primers were designed with Sal I and EcoR1 respectively for cloning purpose. As a result, there were six constructs which differ in size and which each carry several important putative transcription elements. The CaMV 35S promoter was removed by digestion with Hind III and Nco 1 and the plasmid was religated using klenow enzyme to form pCAMBIA-Sesq-Pro (PSP-VF) construct.
The new constructs were labeled as PSPr-VF6 which is ~1336 bp in length and carries the longest SesqPro promoter, PSPr-VF5, which is ~1001 bp in length, PSPr-VF4, which is ~888 bp and PSPr-VF3 which is ~643 bp in length. PSPr-VF2 is ~430 bp in length, while the PSPr-VF1 is ~270 bp long. The presence of the promoter deletion derivatives were confirmed using PCR analysis.
Schematic diagrams of all deletion derivatives are shown in Fig. 1 which explains the location of the promoter SesqPro and its deletion series in the pCAMBIA 1301 vector backbone. The sizes of the deletion derivatives and the important putative transcription element which were present in the different promoter constructs are summarized in Table 1.
As for the SesqPro, the putative transcription initiation site, designated as +1 was located at 33 bp upstream of the ATG translation start codon. A putative TATA box and CAAT box, which are the general signals for eukaryotic gene expression, were observed at positions -25 and -352, respectively and a putative GC box was located at position -237. Both the distance of the consensus sequence, CAAT and GC boxes in SesqPro were comparable to those found in the genome of other plants. Usually, the eukaryotic GC box is located at position -100 (Klug and Cummings, 1994), whereas the CAAT box is located between -80 to -300 bp from +1 (Tasanen et al., 1992).
| Table 1: | Putative elements of transcription factors present in the specified deletion derivates of SesqPro |
 | |
The CAAT box is a common element found in 80% of the eukaryotic promoters and can function in both forward and backward directions. It also plays an important role in TATA-less promoters (Mantovani, 1999). Other motifs present in SesqPro were the (GA)10 element located at -479 and three copies of GAGA boxes at positions -478, -677 and -695. The function of the GAGA box/ factor in plant promoters has not been previously reported, but the GAGA binding factor from Drosophila has been analyzed. The GAGA factor in Drosophila promoter functions as an anti-repressor which could inactivate the repressor protein from inactivating RNA polymerase II during the transcription process (Kerrigan et al., 1991). Two Dof binding factors, located at positions -411 and -450, were also present in the SesqPro promoter. The Dof binding factor has two functions, either activating transcription and being expressed constitutively and ubiquitously or, inactivating transcription and being highly expressed in the stem and root, but being expressed at a lower level in green leaves (Yanagisawa and Sheen, 1998).
Another important putative transcription factor in SesqPro is the G/A hybrid, commonly known as the bZIP element, which is located at position -617 and has ACGT as its core element. The CACGTA element is a homolog to the G box (CACGTG) (Scmidt et al., 1992). An interesting characteristic of the ACGT element is its ability to respond to elicitor induction. Therefore, the G/A hybrid box presented in SesqPro could possibly function as a specific element towards the elicitor induction. This is related to the native role of SesqPro, which controls the expression of the sesquiterpene synthase gene that is involved in the biosynthesis of phytoalexin compounds in anti-fungus and anti-microbe infections in plant (Droge-Lase et al., 1997). SesqPro also contains two copies of short AT-rich elements, AT1 located at position -960, AT2 located at position -1040 and another longer AT-rich element located at position -1180. Studies have shown that short AT elements (~20-55 bp) could enhance the level of gene expression by 6-13 folds either in normal or reverse orientation (Sandhu et al., 1998). The presence of three copies of AT elements in SesqPro may correlate with its high expression of sesquiterpene synthase gene in E. oleifera mesocarp (Cha, 2001). A (CA)n element in SesqPro which is located at position -329 is commonly found in seed storage protein promoters such as the napin gene promoter (Ellerstrom et al., 1996) and β-phaseolin (Burrow et al., 1992). Ellerstrom et al. (1996) suggested that the (CA)n element could function as an activator or repressor with different transcription factors in various tissues.
Analysis on Sesquiterpene Synthase Promoter Specificity Using Tomato Hairy Roots and Oil Palm Tissues
To evaluate the specificity of the sesquiterpene synthase promoter in different tissues, tomato hairy roots were generated from cotyledons via A. rhizogenes-mediated transformation. Conversely, different tissues from oil palm, including the IE, mesocarp slices and leaves had been transformed by A. tumefaciens strain LBA 4404. In this study, PSPr-VF6, the full-length sesquiterpene synthase promoter, was transformed into the selected tissues. Hairy roots and oil palm transformed tissues carrying the CaMV 35S promoter were made as the positive control. Two independent lines were analyzed; one for each tomato hairy root transformed with either a construct containing CaMV 35S promoter or full-length SesqPro promoter.
Based on Fig. 2, two types of transformant tissues, the oil palm tissues with transient transformation and the tomato hairy roots with stable transformation, were used to evaluate the promoter specificity. Both indicated different systems which were homologous for oil palms and heterologous for tomato hairy roots. GUS histochemical assays for all tested tissues showed various positive GUS expressions with different patterns of distribution and intensity. Generally, both the CaMV 35S promoter and SesqPro (PSPr-VF6) were able to express the GUS gene in all tested tissues. From close observations, it appeared that the constitutive CaMV 35S promoter generally showed a higher expression of GUS in the transgenic hairy roots, mesocarp and IE compared to SesqPro. The CaMV 35S promoter is a well-studied and characterized constitutive promoter derived from the CaMV virus (Benfey et al., 1990) and has been shown to have a high expression in dicots compared to monocots (Yang et al., 2003).
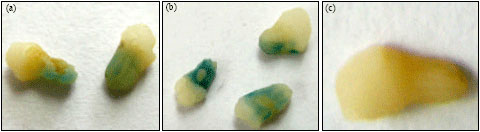 | |
| Fig. 2: | Distribution and intensity of gus gene expression in the transformed immature embryos (IE), mesocarp slices and leaf slices of oil palm (E. oleifera) (a): IE (CaMV 35) (b): IE (SesqPro); (c): IE untransformed (d): mesocarp negative control (untransformed).) (e): mesocarp (CaMV 35S) (f): (promoter SesqPro) (g): leaf slices (CaMV 35S) (h): leaf slices (SesqPro) (I): hairy roots (CaMV 35S) and (j): hairy roots (SesqPro). (k) GUS activity from fluorometric analysis used to evaluate sesquiterpene synthase promoter specificity and CaMV 35S promoter in transformed tissues HR: tomato hairy roots; M: mesocarp slices; IE: Immature embryos; L: Leaf |



SesqPro was isolated from the promoter region of sesquiterpene synthase gene in E. oleifera which was highly expressed in mesocarp tissues developmentally (Cha, 2001). Natively, only the sesquiterpene synthase gene from E. oleifera is highly expressed developmentally without induction by a pathogen. This is in contrast to other sesquiterpene synthase genes such as that in Elaies guineensis where its expression is only activated when it is elicited by the invasion of a pathogen.
Previously, Salwa (2006) conducted a GUS histochemical assay on two different sizes of sesquiterpene synthase promoter isolated from the mesocarp of Elaies guineensis. The shorter Sesyn0.5Pro promoter only contains a TATA box and the longer Sesyn1.2Pro contains similar putative transcription factors as in SesqPro. Both Sesyn0.5Pro and Sesyn1.2Pro showed GUS expression in IE and leaves but no expression was detected in the mesocarp. Sesyn0.5Pro gave a higher GUS expression compared to Sesyn1.2Pro. The absence of the (CA)n element in the Sesyn1.2Pro sequence may have contributed to a lower GUS expression. In contrast, SesqPro was able to drive the GUS expression in all tissues tested, including the heterologous tissues of tomato hairy roots.
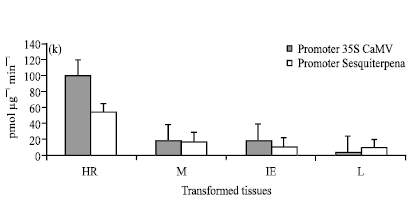
Figure 2k shows the GUS activity from the fluorometric analysis in evaluating the specificity of SesqPro and CaMV 35S promoters in selected transformed tissues. Fluorometric analysis for each sample was conducted in three replicates to increase accuracy. In comparison to different plant species, both monocots and dicots show an obvious quantitative difference in GUS activity. In this study, the level of GUS activity in the transgenic hairy roots was found to be higher than that of oil palm tissues. GUS activity driven by the SesqPro promoter in tomato hairy roots was higher compared to its activity in other oil palm tissues and this could be related to the accumulation of higher sesquiterpenoids in the roots (Maldonado-Bonilla et al., 2008). Besides, the stable transformation system occurring in hairy roots could give more accurate and consistent quantitative results compared to the transient expression in transformed oil palm tissues. The generation of transformed oil palm through tissue culture is time-consuming and therefore transient analysis is the best choice and is feasible for preliminary evaluations of promoter specificity and activity in oil palm tissues. In contrast, generation of stable transgenic lines for tomato hairy roots is relatively easy, as it could grow very fast in a plagiotropic manner (Nilsson and Olsson, 1997).
The level of GUS activity driven by SesqPro (PSPr-VF6) in the mesocarp and IE of oil palms was higher than that of leaves. SesqPro drives a higher GUS expression than the CaMV 35S promoter in leaves, but showed a lower expression in other oil palm tissues such as the mesocarp and IE. This is because SesqPro itself is isolated from the mesocarp and IE and it is also extracted from the endosperm in the mesocarp. Therefore, it is possible that both tissues have the same or similar transcription factors and communicate in line with the same trans factors and cis elements. Conversely, the GUS activity of the CaMV 35S promoter is lower in monocot compared to dicot plants (Basu et al., 2003). This might be due to the CaMV 35S promoter activity itself which involved the internal molecular interaction between cis and trans proteins since phytoalexin activity is also active in leaf tissues (Yedidia et al., 2003).
Based on the results obtained from both histochemical and fluorometric assays, we hypothesized that the SesqPro promoter could potentially be an active and constitutive promoter. This promoter can control and regulate gus gene expression in all tissues tested for its specificity, as can the constitutive CaMV 35S promoter, which is a well-characterized promoter.
Analysis on Sesquiterpene Synthase Promoter Strength in Mesocarp Slices and Immature Embryos
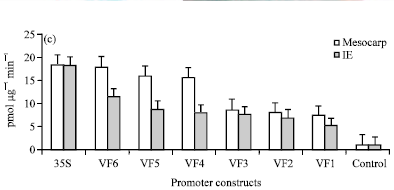
All different sizes of SesqPro deletion derivatives gave positive GUS expression in IE and mesocarp slices (Fig. 3a, b), but the level of GUS activity decreased in line with the removal of 5’ upstream sequences from the full-length to the shortest promoter deletion constructs series in transformed tissues (Fig. 3c).
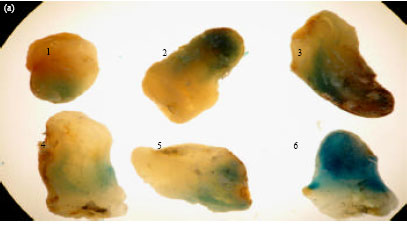 | |
| Fig. 3: | (a) shows results from histochemical assays conducted on IE with different distributions and intensities regarding its promoter deletion derivatives constructs transformed into IE, (b) shows results from histochemical assays conducted on mesocarps with different distributions and intensities with respect to its promoter deletion derivatives constructs transformed into mesocarp slices 1: PSPr-VF1; 2: PSPr-VF2; 3: PSPr-VF3; 4: PSPr-VF4; 5: PSPr-VF5; 6: PSPr-VF6. (c) GUS activity in mesocarp slices and IE of oil palm transformed with different lengths of sesquiterpene synthase promoter and CaMV 35S promoter |
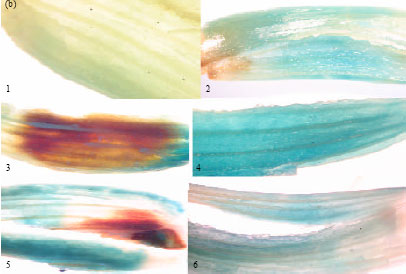
Higher GUS activity was observed in PSPr-VF6-transformed tissue compared to PSPr-VF5 which lack the AT elements in its SesqPro region. This observation may suggest that the AT elements play an important role in regulating the expression level of the sesquiterpene synthase gene.
GUS expression was observed to be lower in PSPr-VF4-transformed tissues compared to those transformed with PSPr-VF5 following the further deletion of both Silenser SBF and Un2 elements in the PSPr-VF4 construct. Deletion of another 200 bp sequence which contains the GAGA box and the hybrid G/A box from PSPr-VF4 gave rise to the PSPr-VF3 construct. The GUS expression observed in PSPr-VF3 was slightly lower in both IE and in the mesocarp compared to PSPr-VF4. The level of GUS expression in both PSPr-VF2 and PSPr-VF1 were still significant even though only the GC box, the CAAT box and (CA)n were left in PSPr-VF2 and TATA box in PSPr-VF1 construct.
Based on fluorometric assays shown in Figure 3c, the two transformed tissues gave similar patterns of GUS activity accordingly. GUS activity driven by CaMV 35S promoter was higher compared to that driven by SesqPro in both oil palm tissues. However, there was an insignificant difference of GUS activity driven by the CaMV 35S promoter in both the mesocarp and IE. Even though both oil palm tissues gave similar patterns of GUS activity, the activity was lower in IE compared to the mesocarp slices. One factor which could contribute to this observation is the original endogenous activity because the SesqPro was initially isolated from mesocarp tissues.
For the full-length SesqPro construct and its deletion derivatives, PSPr-VF6, which contains all of the SesqPro sequence in ~1336 bp, conferred a higher level of GUS activity in comparison to the other five deletion derivatives. As in this construct, all putative transcription motifs present and work accordingly as in its endogenous tissues, even though the GUS activity was slightly lower in IE. Overall, the GUS activity decreased according to its promoter deletion derivatives. As we can see, the GUS activity in PSPr-VF5 decreased insignificantly in both tissues. Following the deletion of a few elements from PSPr-VF5, GUS activity in PSPr-VF4 also decreased but the decrease was still insignificant compared to the other two longer promoter constructs, PSPr-VF6 and PSPr-VF5.
A significant reduction of GUS activity was detected in PSPr-VF3 followed by PSPr-VF2 and PSPr-VF1. This observation was related to the deletion of the GAGA box, the G/A hybrid box and also the Un4 element from the PSPr-VF4 construct. Therefore, these three putative elements could be threshold motifs for the SesqPro activity. However, PSPr-VF3, PSPr-VF2 and PSPr-VF1 could still confer expression based on the GUS activity measured. This was probably due to the presence of several basic but important transcription factors such as the TATA box, the CAAT box and the GC box for minimal transcription activity (Butler and Kadonaga, 2001).
As a result, we presumed that SesqPro is still able to drive the gus gene expression with its basic transcription factors. Even though SesqPro contains a few negative transcription motifs or transcription repressor motifs, it also contains activator motifs/enhancers which could counteract the effect of the repressor activity. Therefore, SesqPro is capable of driving a higher GUS activity if the enhancer motifs are within its vicinity.
Activity of the Sesquiterpene Synthase Promoter in Response to Exogenous Jasmonic Acid
In order to investigate the promoter sequences of the sesquiterpene synthase gene which are responsible for the induction of GUS activity by exogenous JA, all the promoter constructs were tested with regard to the inducibility of GUS expression in 15 day old transformed tissues. Sesquiterpenoid is a defensive compound secreted in higher plants against fungal or microbial infections and is catalyzed by the enzyme sesquiterpene synthase. To study the transcription factor elements related to the defense activity, 100 μM of JA was added to the liquid culture medium of transformed tissues. JA is among the exogenous elicitors applied for the induction of defense mechanisms in plant tissue cultures (Van Etten et al., 1994). JA and its derivatives are believed to induce the production of secondary metabolites in plants (Sanz et al., 2000). JA and its derivative, methyl ester (methyl jasmonate, MJ) were proposed as the key signaling compounds in inducing the accumulation of multiple secondary metabolites (Szabo et al., 1999).
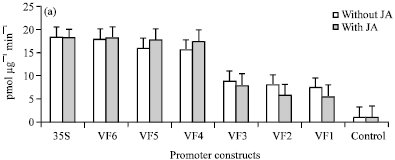 | |
| Fig. 4: | GUS activity induced by 100 μM jasmonic acid in (a) mesocarp slices and (b) IE of oil palm transformed with different lengths of the sesquiterpene synthase promoter and the CaMV 35S promoter |
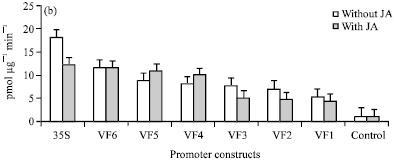
As shown in Fig. 4a and b, the same pattern of GUS activity was observed in transformed mesocarp slices and IE tissues when 100 μM of JA was applied. The GUS activity in both tissues increased upon JA induction in the transformed tissues carrying the PSPr-VF6 construct. This situation was similar to the other two constructs, PSPr-VF5 and PSPr-VF4. The difference in the level of GUS activity in these three constructs was insignificant. However, the GUS activity dropped significantly in PSPr-VF3, PSPr-VF2 and PSPr-VF1 upon induction by JA.
The G/A hybrid box were deleted from the PSPr-VF3 and the other two shorter SesqPro constructs. As discussed earlier, the presence of the G/A hybrid box in the SesqPro sequence will cause it to respond to elicitor inductions such as JA and will activate the biosynthesis of phytoalexins for plant defense mechanisms. The G/A hybrid box is categorized under the bZIP group which consists of the ACGT element and functions in response to environmental stresses and elicitor stimuli. This G/A hybrid box will interact with the accumulated exogenous JA and will activate the transcriptional machineries for plant defense mechanisms. Therefore, this will influence the transcription level of the gus gene and the proteins translated by this gene. The immediate reduction of GUS activity in PSPr-VF3 constructs was due to the loss of the G/A hybrid box in this construct. In the absence of this element, the plant defense mechanism conferred by this bZIP element is defective.
In summary, this preliminary study on the G/A hybrid box has shown that it is essential for SesqPro activation through environmental stress or elicitation by pathogen invasion. The findings reported in this study will serve as the underlying principles for future experiments further characterizing the sesquiterpene synthase promoter.
ACKNOWLEDGMENTS
This research is funded by Agriculture Biotechnology Institute Research Grant (UKM-ABI-NBD0010-2007), Ministry of Science, Technology and Innovation of Malaysia awarded to Dr. Ismanizan Ismail. We would like to thank the Malaysian Oil Palm Board (MPOB) for providing oil palm tissue for this study.
REFERENCES
- Benfey, P.N., L. Ren and N.H. Chua, 1990. Combinatorial and synergistic properties of CaMV 35S enhancer subdomains. EMBO J., 9: 1685-1696.
PubMed - Birnboim, H.C. and J. Doly, 1979. A rapid alkaline extraction procedure for screening recombinant plasmid DNA. Nucleic Acids Res., 7: 1513-1523.
CrossRefPubMedDirect Link - Bohlmann, J., G. Meyer-Gauen and R. Croteau, 1998. Plant terpenoid synthases: Molecular biology and phylogenetic analysis. Proc. Nat. Acad. Sci. USA., 5: 4126-4133.
CrossRefPubMedDirect Link - Bradford, M.M., 1976. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem., 72: 248-254.
CrossRefPubMedDirect Link - Burrow, M.D., P. Sen, C.A. Chlan and N. Murai, 1992. Developmental control of the β-phaseolin gene requires positive, negative, and temporal seed-specific transcriptional regulatory elements and a negative element for stem and root expression. Plant J., 2: 537-548.
CrossRef - Chappell, J., 1995. Biochemistry and molecular biology of the isoprenoid biosynthetic pathway in plants. Ann. Rev. Plant Physiol. Plant Mol. Biol., 46: 521-547.
CrossRef - Ellerstrom, M., K. Stalberg, I. Ezurra and L. Rask, 1996. Functional dissection of napin gene promoter: identification of promoter elements required for embryo and endosperm specific transcription. Plant Mol. Biol., 32: 1019-1027.
PubMed - Jefferson, R.A., T.A. Kavanagh and M.W. Bevan, 1987. GUS fusions: Beta-glucuronidase as a sensitive and versatile gene fusion marker in higher plants. EMBO J., 6: 3901-3907.
Direct Link - Kappers, I.F., A. Aharoni, T.W.J.M. van Herpen, L.L.P. Luckerhoff, M. Dicke and H.J. Bouwmeester, 2005. Genetic engineering of terpenoid metabolism attracts bodyguards to arabidopsis. Science, 309: 2070-2072.
CrossRefPubMedDirect Link - Kerrigan, L.A., G.E. Croston, L.M. Lira and J.T. Kadonaga, 1991. Sequence-specific transcriptional antirepression of the Drosophila kruppel gene by the GAGA factor. J. Biol. Chem., 266: 574-582.
PubMed - Kessler, A. and I.T. Baldwin, 2001. Defensive function of herbivore-induced plant volatile emissions in nature. Science, 291: 2141-2144.
Direct Link - Maldonado-Bonilla, L.D., M. Betancourt-Jimenez and E. Lozoya-Gloria, 2008. Local and systemic gene expression of sesquiterpene phytoalexin biosynthetic enzymes in plant leaves. Eur. J. Plant Pathol., 121: 439-449.
CrossRef - Milat, M.L., P. Ricci, P. Bonnet and J.P. Blein, 1991. Capsidiol and ethylene production by tobacco cells in response to cryptogein, an elicitor from Phytophthora cryptogea. Phytochemistry, 30: 2171-2173.
CrossRef - Murashige, T. and F. Skoog, 1962. A revised medium for rapid growth and bio assays with tobacco tissue cultures. Physiol. Plant., 15: 473-497.
CrossRefDirect Link - Nilsson, O. and O. Olsson, 1997. Getting to the root: the role of the Agrobacterium rhizogenes rol genes in the formation of hairy roots. Physiol. Plant, 100: 463-473.
CrossRef - Picaud, S., M.E. Olsson, M. Brodelius and P.E. Brodelius, 2006. Cloning, expression, purification and characterization of recombinant (+)-germacrene D synthase from Zingiber officinale. Arch. Biochem. Biophys., 452: 17-28.
PubMed - Proctor, R.H. and T.M. Hohn, 1993. Aristolochene synthase: Isolation, characterization and bacterial expression of sesquiterpenoid biosynthetic gene (Ari1) from Penicillium roqueforti. J. Biol. Chem., 268: 4543-4548.
PubMed - Sanz, M.K., X.E. Hernandez, C.E. Tonn and E. Guerreiro, 2000. Enhancement of tessaric acid production in Tessaria absintheoides cell suspension cultures. Plant Cell Rep., 19: 821-824.
CrossRef - Szabo, E., A. Thelen and M. Petersen, 1999. Fungal elicitor preparations and methyl jasmonate enhance rosmarinic acid accumulation in suspension cultures of Coleus blumei. Plant Cell Rep., 18: 485-489.
CrossRef - Tasanen, K., J. Oikarinen, K.I. Kivirikko and T. Pihlajaniemi, 1992. Promoter of the gene for the multifunctional protein disulfide isomerase polypeptide. Functional significance of the 6 CCAAT boxes and other promoter elements. J. Biol. Chem., 267: 11513-11519.
PubMed - Trapp, S.C. and R.B. Croteau, 2001. Genomic organization of plant terpene synthases and molecular evolutionary implications. Genetics, 158: 811-832.
Direct Link - Van Etten, H.D., J.W. Mansfield, J.A. Bailey and E.E. Farmer, 1994. Two classes of plant antibiotics: Phytoalexins versus phytoanticipins. Plant Cell, 6: 1191-1192.
CrossRefPubMedDirect Link - Yang, M., R. Bower, M.D. Burow, A.H. Paterson and T.E. Mirkov, 2003. Genomics, molecular genetics and biotechnology: A rapid and direct approach to identify promoters that confer high levels of gene expression in monocots. Crop Sci., 43: 1805-1813.
Direct Link - Yin, S., L. Mei, J. Newman, K. Back and J. Chappell, 1997. Regulation of sesquiterpene cyclase gene expression. Characterization of an elicitor-and pathogen-inducible promoter. Plant Physiol., 115: 437-451.
PubMed - Chu, C.C., C.C. Wang, C.S. Sun, K.C. Hsu, K.C. Yin, C.Y. Chu and F.Y. Bi, 1975. Establishment of an efficient medium for anther culture of rice through comparative experiments on the nitrogen sources. Scient. Sin., 18: 659-668.
Direct Link - Droge-Laser, W., A. Kaiser, W.P. Lindsay, B.A. Halkier, G.J. Loake, P. Doener, R.A. Dixon and C. Lamb, 1997. Rapid stimulation of a soybean protein-serine kinase that phosphorylate a novel bZIP DNA-binding protein, G/HBF1, during the induction of early transcription-dependent defenses. EMBO J., 16: 726-738.
CrossRef