
V. Asuzu Onyeka
Department of Animal Production and Health, Faculty of Agriculture, Federal University, Oye-Ekiti, Ekiti State, Nigeria
O. Nwaehujor Chinaka
Department of Biochemistry, Faculty of Basic Medical Sciences, University of Calabar, P.M.B. 1115, Calabar, Nigeria
LiveDNA: 234.22056
O. Ode Julius
Department of Veterinary Pharmacology and Toxicology, Faculty of Veterinary Medicine, University of Abuja, Nigeria
C. Nwobi Obichukwu
Department of Animal Production and Health, Faculty of Agriculture, Federal University, Oye-Ekiti, Ekiti State, Nigeria
American Journal of Drug Discovery and Development
Year: 2015 | Volume: 5 | Issue: 1 | Page No.: 24-33







ABSTRACT
This study was aimed at investigating the effects of chronic alcohol consumption on immunochemistry of mice experimentally infected with E. coli strain 0157:H7. The bacterial organisms used were collected from an already identified and characterized stock (ECO 6) maintained for research purposes in the Laboratory of the Department of Veterinary Microbiology and Pathology, Faculty of Veterinary Medicine, University of Nigeria, Nsukka. About 1.15x104 CFU was used to challenge mice which were placed on prior treatment with various concentrations (10, 20, 30 and 40%) of ethanol for 3 weeks in separate groups, intraperitoneally. Blood samples were collected 7 days post bacterial challenge from the experimental mice via the retrobulbar plexus in the median canthus of the eyes. The samples collected into Bijou bottles were subjected to immunochemistry assay (hemagglutination test and Hemagglutination Inhibition (HI) test). The mean antibody titer in 40% alcohol-treated mice (341.33±85.33) was found to be significantly (p<0.05) lower than the value induced with 30% alcohol (512.00±0.00). The titer values however, produced with 20% (1024.00±0.00) and 10% alcohol (1706.70±341.33) were not only elevated but highly significant (p<0.01) compared to the alcohol effects at 30 or 40%, respectively. There was concentration dependent decrease in antibody titer induced with alcohol in the experimental mice. Low antibody titer correlates with decreased immune capability. Chronic exposure to alcohol could have depressed immunity due impaired cell proliferation in the hematopoietic tissues.
PDF Abstract XML References Citation
Received: December 16, 2014;
Accepted: January 26, 2015;
Published: June 22, 2015
How to cite this article
V. Asuzu Onyeka, O. Nwaehujor Chinaka, O. Ode Julius and C. Nwobi Obichukwu, 2015. Chronic Alcohol Ingestion and its Effect on Immunochemistry in Albino Mice
Experimentally Challenged with Escherichia coli Strain 0157:H7. American Journal of Drug Discovery and Development, 5: 24-33.
DOI: 10.3923/ajdd.2015.24.33
URL: https://scialert.net/abstract/?doi=ajdd.2015.24.33
DOI: 10.3923/ajdd.2015.24.33
URL: https://scialert.net/abstract/?doi=ajdd.2015.24.33
INTRODUCTION
The immune system serves as the body’s defense against infections, damage caused by foreign substances and uncontrolled tumor growth (Cook, 1998). The immune system has two arms: The innate or non-specific immunity and acquired or specific immunity. The non-specific immunity exists in the body before exposure to a pathogen and it does not respond to specific antigen challenge (Sriskandan et al., 2007). Acquired immunity on the other hand, is developed post exposure to a pathogen and it responds only to a particular pathogen (Cook, 1998).
Alcohol abuse is considered to contribute to immune deficiencies (Walker et al., 2009). Alcoholics are immunodeficient but they often have elevated blood levels of autoimmune antibodies (Szabo, 1999). Diseases seen in alcoholics due to immunodeficiencies include cancers, bacterial pneumonia, septicemia, tuberculosis, hepatitis C, meningitis, lung abscess, diphtheria and cellulites (Cook, 1998). Alcoholics also respond poorly to treatment (Spitzer and Meadows, 1999). Immunodeficiencies occur as a result that alcohol inhibits the function of cells that ingest and destroy invading microorganisms (neutrophils, eosinophils and macrophages). Also, both acute and chronic alcohol exposure affects the production of signaling molecules that coordinate the immune response i.e., cytokines and this adversely affects the function of the cells that mediate the immune response against specific microorganisms and long term immunity i.e., T-cell and B-cells (Szabo, 1999).
In humans and animals, the occurrence of the disease condition Colibacillosis is common. Colibacillosis is caused by the pathogen Escherichia coli normally abbreviated as E. coli. The harmless strains of this organism are normally part of the normal flora of the gut and can benefit their hosts by producing vitamin K2 (Bentley and Meganathan, 1982) and by preventing the establishment of pathogenic bacteria within the intestine (Hudault et al., 2001; Reid et al., 2001). However, sometimes these organisms leave the gut and go into other parts of the body, or in the gut, some strains develop traits that can be harmful to a host animal. These virulent strains typically cause a bout of diarrhoea that is unpleasant in healthy adults and is often lethal to children in the developing world (Nataro and Kaper, 1998). More virulent strains, such as O157:H7 cause serious illness or death in the elderly, the very young or the immunocompromised (Nataro and Kaper, 1998; Hudault et al., 2001).
In general, it is been shown that E. coli normally inhabits the gut area, but it is also known that alcohol has some effect on the mucosal lining of the gut and some adverse effect on the immune system. There is currently a dearth of information on the effects of alcohol consumption on E. coli infection either in human or animal models.
The objective of this study was to study the effects of chronic alcohol consumption on immunochemistry of mice experimentally infected with E. coli strain 0157:H7.
MATERIALS AND METHODS
Preparation, harvest and mice infection with E. coli strain 0157:H7: The bacteria were collected from an already identified and characterized stock (ECO 6) which has previously being maintained for research purposes in the Microbiology Laboratory of the Department of Veterinary Microbiology and Pathology, Faculty of Veterinary Medicine, University of Nigeria, Nsukka. With a sterile wire-loop, some bacteria were streaked into a petri-dish containing Mac Conkey agar. A colony from here was harvested and sub-cultured on an Eosin Methylene blue agar to confirm that the colony was Escherichia coli. A colony was further subcultured on sorbitol agar (10 g L-1 sorbitol instead of lactose) for isolation of the test organism. Nutrient broth (100 mL) was prepared and 10 mL of the broth was put into 10 test-tubes. These were clogged with cotton-wool and autoclaved at 121°C for 15 min. A colony of E. coli O157:H7 harvested from the growth on sorbitol agar growth was introduced into the first test-tube. This was properly mixed and 1 mL of the mixed solution was collected and transferred into the next test-tube. This process was used to achieve a serial dilution in all the 10 test-tubes. A pre-determined volume (20 mL) of Mac Conkey agar was poured into 10 universal bottles and was autoclaved. On cooling, 0.1 mL from each of the above serially diluted test-tubes was introduced into the separate universal bottles. Then after mild, yet thorough mixing, the content of each universal bottle was poured into one of already sterilized petri-dishes and incubated at 37°C for growth of colonies. Twenty-four hours later, the plates were examined and colony growth was counted. The concentration of known organisms per dilution extrapolated from the parent stock, was used to determine the volume for infecting the mice.
Animals and experimental design: Eight weeks old albino mice (26.6-35.3 g) of both sexes, sourced from the Laboratory Animal Unit of the Faculty of Veterinary Medicine, University of Nigeria, Nsukka (UNN) were used in the study. Animals were acclimatized for 14 days before commencement of the experiment. The mice were fed with commercial feed (Vital feed®, Grand Cereal Oil Mill Ltd., Nigeria) and provided with clean water ad libitum. The albino mice were divided into 6 groups of 12 mice each using stratified random selection method and were kept in clean cages in the Animal House of the Department of Veterinary Anatomy, Faculty of Veterinary Medicine, UNN. Group 1 was given 10% ethanol (V/V) in their drinking water. Group 2 received 20% of ethanol solution. The third group received 30% of ethanol while group 4 and 5 received 40% of alcohol ad-libitum, respectively. Group 6 served as normal control and received only water. The alcoholic groups received ethanol for 3 weeks to establish a chronic state of alcoholism before groups 1-4 were then challenged with E. coli strain 0157:H7. Group 5 was not challenged with E. coli. Seven days after the challenge, the mice in each group were humanely euthanized. Ethical conditions governing the conduct of experiments with life animals were strictly observed (Ward and Elsea, 1977; Zimmermann, 1983; Anonymous, 1996).
Mice vaccination: Animal groups used for the viral test were challenged with Lasota virus, which was obtained from the National Veterinary Research Institute (NVRI), Vom Nigeria. The vaccine was procured in a condensed form and was re-constituted with 3 mL diluent to obtain a solution for inoculation. From the vile, 0.1 mL of the vaccine was used to challenge the animals. Virus was administered through the intra-peritoneal route to each animal in the various groups. In this test, only 10, 20, 30 and 40% alcohol treatment was used to evaluate ethanol-induced concentration variance in antibody titre of the experimental animals. Blood samples were subsequently collected into Bijou bottles from the retrobulbar plexus via the median canthus of the eye of the challenged mice for immunochemistry assay as described previously.
Hemagglutination (HA) test: The test was carried out as described by Ochei and Kolhatkar (2009). To each well of the micro titer plate, 20 μL of PBS (pH 7.2) was carefully added. A serial dilution of the test antigen was made in each row of the wells. To each of the wells, 20 μL of washed chicken erythrocytes was also added. The plates were then covered and incubated at room temperature for 25 min. The control of the test was also carried out but on another row of wells, which was void of the test antigen.
Hemagglutination Inhibition (HI) test: Mouse sera, Newcastle antigen obtained from NVRI, Vom Nigeria, Erythrocytes (0.6% chicken erythrocyte) and diluent, Phosphate Buffered Saline (PBS) at a pH value of 7.2 were used for the study.
PBS (0.02 mL) was added to each row of the V-bottomed micro-titer plate. A two-fold serial dilution of 0.02 mL of the mice sera was made in each row of the well. To each well was added 0.02 mL of the antigen and this was allowed to incubate at room temperature for 45 min. The washed chicken RBC (erythrocytes) was then added to each of the wells and this was read after 25 min. All control protocols for the test were set up as required.
Statistical analysis: The data obtained were subjected to analysis of variance (ANOVA) and the means separated using Duncan’s multiple range tests (Duncan, 1955), differences at 95 and 99% confidence intervals were considered significant.
RESULTS
Preparation, harvest and infection of E. coli strain 0157:H7: The organisms (E. coli) produced pinkish colonies on Mac Conkey agar due to lactose fermentation but the test strain (E. coli O157:H7) was selectively isolated based on formation of straw colored colonies on sorbitol agar. From the result obtained after the colony count, it was deduced that the parent stock contained about 2.3x108 CFU mL-1. A pre-determined volume (0.5 mL) of the parent stock containing about 1.15x104 CFU was the concentration of organisms administered intraperitoneally to challenge each mouse.
Hemagglutination (HA) test: The HA titer was 128 which was divided by 4 to obtain the 4 HAU (Haemoagglutination unit) and thus making a total 32 haemoagglutination units.
Hemagglutination Inhibition (HI) test: The blood antibody titer in 40% alcohol-treated group (341.33±85.33) was found to be significantly (p<0.05) lower than that of 30% alcohol-treated rat group (512.00±0.00) but at the same time, the titer value with 40% alcohol treatment became highly significantly (p<0.01) reduced compared to values in rats treated with 20 or 10% alcohol. Rats in 10% alcohol-treated group had a significantly (p<0.01) higher elevated mean titer value of 1706.70±341.33 relative to that of 20% alcohol-treated rats with 1024.00±0.00. There was an observed concentration dependent decrease in antibody titer with respect to the effect of the alcohol in the experimental rats (Fig. 1).
DISCUSSION
In the present study, 40% alcohol consumption induced a five-fold depression in antibody level equating to a titer value of 341.33±85.33 relative to 1706.70±341.33 produced with 10% alcohol treatment in mice. There was a corresponding increase in antibody titer with a measured reduction in the concentration alcohol treatment (Fig. 1). This implies a reduction in antibody generation with increased concentration of alcohol in mice.
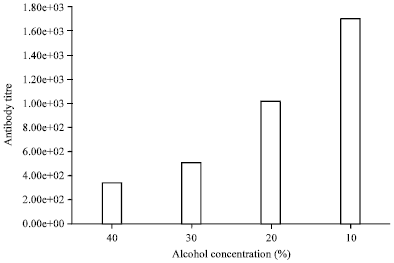 |
| Fig. 1: | Comparison of antibody titer of the different alcohol-treated rat groups |
This observation was supported by the results of earlier studies that, chronic consumption of alcohol induced a decrease in the number of B cells in the spleen (Zhang et al., 2012). Recent research findings also revealed a spectrum of alcohol induced immune dysfunction in humans and animal models. Chronic alcohol consumption impairs B-cells distribution, compromises circulation and interferes with some aspects of the B-cells functioning (Chang et al., 1990). In response to some antigens, B-cells require the assistance of cytokines secreted by the T-cells (T dependent responses), whereas in response to other antigens, cell activation by cytokines is not required (i.e., T independent responses) (Szabo, 1999). Alcohol appears to affect these responses differently because B-cells in the spleen of alcoholic animals showed impaired proliferation during the T-dependent responses and not in the T-independent responses (Hussain, 2010). Chronic alcohol exposure in rodent or murine models impaired the T-dependent B-lymphocytes responses leading to antibody secretion (Szabo, 1999).
Immunodeficiencies result, when alcohol inhibits the function of phagocytes that ingest and destroy invading microorganisms (neutrophils, eosinophils and macrophages). Also, both acute and chronic alcohol exposure affects the production of signaling molecules that coordinate the immune response i.e., cytokines and this adversely affects the function of the cells that mediate the immune response against specific microorganisms and long term immunity that is, T-cell and B-cells (Szabo, 1999), thus reducing their ability to fight off introduced challenges to the system. It could be deduced that with an increase in the concentration of alcohol, the lymphatic systems are made less sensitive or less active, thus increasing the capability of the infecting agent to cause more damage in the body. This position is however at variance with the reasoning of some experts who suspect that alcohol exerts an "all-or-none" effect on immune response-that is, the presence or absence of alcohol, rather than its amount, dictates the immune response (McGill et al., 1995; Messingham et al., 2002).
Both acute and chronic alcohol exposure affects the production of signaling molecules (cytokines) that coordinate immune response against specific microorganisms and long term immunity i.e., T-cell and B-cells (Szabo, 1999). Previous studies reported that the number of B-cells in the spleen decreased (Zhang et al., 2012).
An overwhelming amount of evidence revealed that both acute and chronic alcohol exposure suppresses all branches of the immune system, including early responses to infection and the tumor surveillance system (Cook, 1998; Diaz et al., 2002; Nelson and Kolls, 2002; Messingham et al., 2002). For example, there is a decrease in the ability to recruit and activate germ-killing white blood cells (Deaciuc, 1997) and an increase in the incidence of breast cancer in people who consume alcohol (Smith-Warner et al., 1998; Zhang et al., 1999).
Some experts suspect that alcohol exerts an "all-or-none" effect on immune response-that is, the presence or absence of alcohol, rather than its amount, dictates the immune response (McGill et al., 1995; Messingham et al., 2002). Other researchers believe that low doses of alcohol-the amount equivalent to a glass of wine-can confer health benefits, including protection against damage to the cardiovascular (Holman et al., 1996) and immune systems (Mendenhall et al., 1997). Such benefits, if they are present, may be attributable to antioxidants in alcoholic beverages such as red wine. In any case, health experts agree that the beneficial effects of antioxidants in some alcoholic beverages are lost if the level of alcohol consumption is elevated (Hanna et al., 1992).
There are several mechanisms by which alcohol impedes immune function. First, alcohol impairs the ability of white blood cells, neutrophils to migrate to sites of injury and infection, a process called chemotaxis (Bautista, 2001). In addition, removing germ-fighting white blood cells (macrophages) and proteins that act as messengers between immune cells (cytokines) from an animal that has not been given alcohol and culturing them in the presence of alcohol, or isolating these cells from humans or animals after administering alcohol, has been shown to alter production of these macrophages and cytokines (Deaciuc, 1997; Szabo, 1998, 1999).
Rodent studies also show that animals are more vulnerable to infection after chronic or acute exposure to alcohol (Deaciuc, 1997; Cook, 1998; Messingham et al., 2002). This increase in susceptibility is equally dramatic in human patients who sustain traumatic injury (Smith and Kraus, 1988; Brezel et al., 1988). Those who have consumed alcohol prior to their injury are six times more likely to die than are alcohol-free patients with comparable injuries (McGill et al., 1995). The mechanisms responsible for this increased mortality are unknown, but it is thought that alcohol compromises the immune system's ability to quickly fight infection by unidentified invaders-a function of the innate immune system (Faunce et al., 1997; Cook, 1998).
To date, only a handful of studies have directly examined gender differences in the effects of alcohol on inflammatory and immune responses (Grossman et al., 1993; Spitzer and Zhang, 1996; Li et al., 1998; Spitzer, 1999). These studies were conducted in rodents and employed different methods, including varying the quantity and duration of alcohol exposure. The reports showed that in the absence of alcohol exposure, inflammatory and immune responses are stronger in females than in males (Grossman et al., 1993; Spitzer and Zhang, 1996; Spitzer and Meadows, 1999). However, the increased immunity in females is nullified by alcohol exposure. For example, in one study, proliferation of white blood cells was suppressed in alcohol-exposed female rats (Grossman et al., 1993); however, investigation also showed that alcohol induced an increase in antibody production. In two other studies, female rats were less able to fight infection when intoxicated (Spitzer and Zhang, 1996; Li et al., 1998). The mechanisms driving these effects remain uncertain. One possibility is that gender differences in inflammatory and immune responses following alcohol exposure stem from alcohol-induced changes in the production of gonadal steroid hormones, such as estrogen and testosterone.
In general, estrogen stimulates immune responses and testosterone is immunosuppressive (Grossman, 1989; Morell, 1995; Cannon and St Pierre, 1997; Verthelyi, 2001; Burger and Dayer, 2002). During their reproductive years, females have more vigorous cellular and humoral immune responses than do males. This heightened immunity in females is evidenced by a more developed thymus that is involved in the maturation of immune cells, higher antibody concentrations and a greater ability to reject tumors and transplanted tissues. Ironically, the enhanced immune function in women of reproductive age is associated with a higher prevalence of autoimmune disorders than is found in postmenopausal women or in men (although estrogen is present in males, its concentration is too low to affect immune response).
The effects of alcohol on production of the gonadal steroid hormones are well documented (Van Thiel et al., 1987; Gavaler and Van Thiel, 1992; Gavaler et al., 1993; Emanuele and Emanuele, 2001). In women, chronic alcohol exposure causes an initial increase in estrogen levels, followed by a marked decrease (Gavaler and Van Thiel, 1992; Gavaler et al., 1993). In men, chronic alcohol consumption causes a decrease in testosterone (Emanuele and Emanuele, 2001). The alcohol-induced decrease in testosterone levels is significant enough to cause atrophy of the testes, impotence and loss of secondary sex characteristics (Van Thiel et al., 1987).
Like other stressors, alcohol stimulates a neuroendocrine network known as the Hypothalamic-Pituitary-Adrenal (HPA) axis, resulting in a dampening of the immune response (Eskandari and Sternberg, 2002). This process begins with activation of the hypothalamus which produces Corticotropin-Releasing Hormone (CRH). This triggers the pituitary gland to secrete adrenal corticotropic hormone (ACTH). Finally, ACTH stimulates the adrenal glands to release glucocorticoids (cortisol in humans and corticosterone in rodents). These steroid hormones, which direct the activity of many cell types, are transmitted throughout the body in the blood. At high levels, they suppress inflammatory and immune responses (Goulding and Guyre, 1993; Da Silva, 1999). Several studies have documented that under resting conditions and in response to stress, females have higher levels of glucocorticoids than do men (Kant et al., 1983; Chisari et al., 1995). Furthermore, estrogen stimulates glucocorticoid production in females (Burgess and Handa, 1992; Carey et al., 1995), whereas testosterone suppresses its production in both male and female subjects (Carlstrom and Stege, 1990; De Weerdt and Gooren, 1992; Handa et al., 1994). Alcohol exposure stimulates glucocorticoid production in both males and females (Ogilvie et al., 1998; Eskandari and Steinberg, 2002).
CONCLUSION
The findings from the study suggest that chronic and high dose alcohol consumption depresses immune response to antigen challenge in experimental mice. There is a critical need for human population to guard against addiction or high dose intake of alcohol.
REFERENCES
- Chang, M.P., D.C. Norman and T. Makinodan, 1990. Immunotoxicity of alcohol in young and old mice. I. in vitro suppressive effects of ethanol on the activities of T and B immune cells of aging mice. Alcoholism: Clin. Exp. Res., 14: 210-215.
CrossRefDirect Link - Hussain, D., 2010. Stress, immunity and health: Research findings and implications. Int. J. Psychosocial Rehab., 15: 94-100.
Direct Link - Sriskandan, S., L. Faulkner and P. Hopkins, 2007. Streptococcus pyogenes: Insight into the function of the streptococcal superantigens. Int. J. Biochem. Cell Biol., 39: 12-19.
CrossRefDirect Link - Walker, Jr. J.E., A.R. Odden, S. Jeyaseelan, P. Zhang, G.J. Bagby, S. Nelson and K.I. Happel, 2009. Ethanol exposure impairs LPS‐induced pulmonary LIX expression: Alveolar epithelial cell dysfunction as a consequence of acute intoxication. Alcoholism: Clin. Exp. Res., 33: 357-365.
CrossRefDirect Link - Spitzer, J.H. and G.G. Meadows, 1999. Modulation of perforin, granzyme A and granzyme B in murine natural killer (NK), IL2 stimulated NK and lymphokine-activated killer cells by alcohol consumption. Cell. Immunol., 194: 205-212.
CrossRefDirect Link - Zhang, H., Z. Zhu and G.G. Meadows, 2012. Chronic alcohol consumption impairs distribution and compromises circulation of B cells in B16BL6 melanoma-bearing mice. J. Immunol., 189: 1340-1340.
Direct Link - Bentley, R. and R. Meganathan, 1982. Biosynthesis of vitamin K (Menaquinone) in bacteria. Microbiol. Rev., 46: 241-280.
Direct Link - Brezel, B.S., J.M. Kassenbrock and J.M. Stein, 1988. Burns in substance abusers and in neurologically and mentally impaired patients. J. Burn Care Res., 9: 169-171.
Direct Link - Burger, D. and J.M. Dayer, 2002. Cytokines, acute‐phase proteins and hormones: IL-1 and TNF-α production in contact-mediated activation of monocytes by T lymphocytes. Ann. N. Y. Acad. Sci., 966: 464-473.
CrossRef - Burgess, L.H. and R.J. Handa, 1992. Chronic estrogen-induced alterations in adrenocorticotropin and corticosterone secretion and glucocorticoid receptor-mediated functions in female rats. Endocrinology, 131: 1261-1269.
CrossRefDirect Link - Cannon, J.G. and B.A. St Pierre, 1997. Gender differences in host defense mechanisms. J. Psychiatr. Res., 31: 99-113.
CrossRefDirect Link - Carey, M.P., C.H. Deterd, J. De Koning, F. Helmerhorst and E.R. De Kloet, 1995. The influence of ovarian steroids on hypothalamic-pituitary-adrenal regulation in the female rat. J. Endocrinol., 144: 311-321.
CrossRefDirect Link - Carlstrom, K. and R. Stege, 1990. Adrenocortical function in prostatic cancer patients: Effects of orchidectomy or different modes of estrogen treatment on basal steroid levels and on the response to exogenous adrenocorticotropic hormone. Urol. Int., 45: 160-163.
CrossRefDirect Link - Chisari, A., M. Carino, M. Perone, R.C. Gaillard and E. Spinedi, 1995. Sex and strain variability in the rat Hypothalamo-Pituitary-Adrenal (HPA) axis function. J. Endocrinol. Invest., 18: 25-33.
PubMed - Cook, R.T., 1998. Alcohol abuse, alcoholism and damage to the immune system-a review. Alcoholism: Clin. Exp. Res., 22: 1927-1942.
CrossRef - Da Silva, J.A.P., 1999. Sex hormones and glucocorticoids: Interactions with the immune system. Ann. N. Y. Acad. Sci., 876: 102-118.
CrossRefDirect Link - De Weerdt, O. and L.J.G. Gooren, 1992. Patterns of serum cortisol levels in ovariectomized females with and without androgen administration. Horm. Metab. Res., 24: 82-84.
CrossRef - Emanuele, M.A. and N. Emanuele, 2001. Alcohol and the male reproductive system. Alcohol Res. Health, 25: 282-287.
Direct Link - Eskandari, F. and E.M. Sternberg, 2002. Neural-immune interactions in health and disease. Ann. N. Y. Acad. Sci., 966: 20-27.
CrossRefDirect Link - Faunce, D.E., M.S. Gregory and E.J. Kovacs, 1997. Effects of acute ethanol exposure on cellular immune responses in a murine model of thermal injury. J. Leukocyte Biol., 62: 733-740.
Direct Link - Gavaler, J.S. and D.H. Van Thiel, 1992. The association between moderate alcoholic beverage consumption and serum estradiol and testosterone levels in normal postmenopausal women: Relationship to the literature. Alcoholism: Clin. Exp. Res., 16: 87-92.
CrossRefDirect Link - Gavaler, J.S., S.R. Deal, D.H. Thiel, A. Arria and M.J. Allar, 1993. Alcohol and estrogen levels in postmenopausal women: The spectrum of effect. Alcoholism: Clin. Exp. Res., 17: 786-790.
CrossRefDirect Link - Grossman, C., 1989. Possible underlying mechanisms of sexual dimorphism in the immune response, fact and hypothesis. J. Steroid Biochem., 34: 241-251.
CrossRef - Grossman, C.J., M. Nienaber, C.L. Mendenhall, P. Hurtubise and G.A. Roselle et al., 1993. Sex differences and the effects of alcohol on immune response in male and female rats. Alcoholism: Clin. Exp. Res., 17: 832-840.
CrossRef - Goulding, N.J. and P.M. Guyre, 1993. Glucocorticoids, lipocortins and the immune response. Curr. Opin. Immunol., 5: 108-113.
CrossRefDirect Link - Handa, R.J., K.M. Nunley, S.A. Lorens, J.P. Louie, R.F. McGivern and M.R. Bollnow, 1994. Androgen regulation of adrenocorticotropin and corticosterone secretion in the male rat following novelty and foot shock stressors. Physiol. Behav., 55: 117-124.
CrossRefDirect Link - Hanna, E., M.C. Dufour, S. Elliott, F. Stinson and T.C. Harford, 1992. Dying to be equal: Women, alcohol and cardiovascular disease. Br. J. Addiction, 87: 1593-1597.
CrossRefDirect Link - Holman, C.D., D.R. English, E. Milne and M.G. Winter, 1996. Meta-analysis of alcohol and all-cause mortality: A validation of NHMRC recommendations. Med. J. Aust., 164: 141-145.
Direct Link - Hudault, S., J. Guignot and A.L. Servin, 2001. Escherichia coli strains colonising the gastrointestinal tract protect germfree mice against Salmonella typhimurium infection. Gut, 49: 47-55.
CrossRefDirect Link - Kant, G.J., R.H. Lenox, B.N. Bunnell, E.H. Mougey, L.L. Pennington and J.L. Meyerhoff, 1983. Comparison of stress response in male and female rats: Pituitary cyclic AMP and plasma prolactin, growth hormone and corticosterone. Psychoneuroendocrinology, 8: 421-428.
CrossRefDirect Link - Li, X., C.J. Grossman, C.L. Mendenhall, P. Hurtubise, S.D. Rouster, G.A. Roselle and P. Gartside, 1998. Host response to mycobacterial infection in the alcoholic rat: Male and female dimorphism. Alcohol, 16: 207-212.
CrossRefDirect Link - McGill, V., A. Kowal-Vern, S.G. Fisher, S. Kahn and R. Gamelli, 1995. The impact of substance use on mortality and morbidity from thermal injury. J. Trauma-Injury, Infect. Crit. Care, 38: 931-934.
Direct Link - Mendenhall, C.L., S.A. Theus, G.A. Roselle, C.J. Grossman and S.D. Rouster, 1997. Biphasic in vivo immune function after low-versus high-dose alcohol consumption. Alcohol, 14: 255-260.
CrossRef - Messingham, K.A.N., D.E. Faunce and E.J. Kovacs, 2002. Review article: Alcohol, injury and cellular immunity. Alcohol, 28: 137-149.
CrossRef - Morell, V., 1995. Zeroing in on how hormones affect the immune system. Science, 269: 773-775.
CrossRefDirect Link - Nataro, J.P. and J.B. Kaper, 1998. Diarrheagenic Escherichia coli. Clin. Microbiol. Rev., 11: 142-201.
Direct Link - Nelson, S. and J.K. Kolls, 2002. Alcohol, host defence and society. Nat. Rev. Immunol., 2: 205-209.
CrossRefDirect Link - Ogilvie, K., S. Lee, B. Weiss and C. Rivier, 1998. Mechanisms mediating the influence of alcohol on the hypothalamic‐pituitary‐adrenal axis responses to immune and nonimmune signals. Alcoholism: Clin. Exp. Res., 22: 243S-247S.
CrossRefDirect Link - Reid, G., J. Howard and B.S. Gan, 2001. Can bacterial interference prevent infection? Trends Microbiol., 9: 424-428.
CrossRef - Smith, G.S. and J.F. Kraus, 1988. Alcohol and residential, recreational and occupational injuries: A review of the epidemiologic evidence. Annu. Rev. Public Health, 9: 99-121.
CrossRefDirect Link - Spitzer, J.A. and P. Zhang, 1996. Gender differences in phagocytic responses in the blood and liver and the generation of cytokine‐induced neutrophil chemoattractant in the liver of acutely ethanol‐intoxicated rats. Alcoholism: Clin. Exp. Res., 20: 914-920.
CrossRefDirect Link - Szabo, G., 1998. Monocytes, alcohol use and altered immunity. Alcoholism: Clin. Exp. Res., 22: 216S-219S.
CrossRefDirect Link - Szabo, G., 1999. Consequences of alcohol consumption on host defence. Alcohol Alcoholism, 34: 830-841.
CrossRefDirect Link - Van Thiel, D.H., J.S. Gavaler, E. Rosenblum and P.K. Eagon, 1987. Effects of ethanol on endocrine cells: Testicular effects. Ann. N. Y. Acad. Sci., 492: 287-302.
CrossRefDirect Link - Verthelyi, D., 2001. Sex hormones as immunomodulators in health and disease. Int. Immunopharmacol., 1: 983-993.
CrossRefDirect Link - Smith-Warner, S.A., D. Spiegelman, S.S. Yaun, P.A. van den Brandt and A.R. Folsom et al., 1998. Alcohol and breast cancer in women: A pooled analysis of cohort studies. J. Am. Med. Assoc., 279: 535-540.
CrossRefDirect Link - Zhang, Y., B.E. Kreger, J.F. Dorgan, G.L. Splansky, L.A. Cupples and R.C. Ellison, 1999. Alcohol consumption and risk of breast cancer: The Framingham study revisited. Am. J. Epidemiol., 149: 93-101.
Direct Link - Zimmermann, M., 1983. Ethical guidelines for investigations of experimental pain in conscious animals. Pain, 16: 109-110.
CrossRefPubMedDirect Link