
Ganiyu Oboh
Department of Biochemistry, Federal University of Technology, P.M.B. 704, Akure, Nigeria
Asian Journal of Clinical Nutrition
Year: 2009 | Volume: 1 | Issue: 1 | Page No.: 40-49







ABSTRACT
This study seeks to access the protective effect of sour tea (Hibiscus sabdariffa, Calyx) and green tea (Camellia sinensis) on some pro-oxidants [Fe (II), sodium nitroprusside, quinolinic acid] induced lipid peroxidation in rat`s brain-in vitro. Aqueous extracts of both teas were prepared (1 g tea in 100 mL of hot water). Thereafter, the ability of the extracts to prevent 25 μM FeSO4, 7 μM sodium nitroprusside and 1 mM quinolinic acid induced lipid peroxidation in isolated rat`s brain tissues preparation was determined in vitro. Subsequently, the total phenol content, reducing power, Fe (II) chelating and OH radical scavenging ability were determined. The results of the study revealed that both teas significantly (p<0.05) inhibited lipid peroxidation in basal and pro-oxidants induced lipid peroxidation in the rats brain homogenates in a dose dependent manner. Also, the teas had high total phenol content [sour (13.3 mg g-1; green (24.5 mg g-1)], reducing power, Fe (II) chelating and OH radical scavenging ability (except sour tea). However, green tea had a significantly higher (p<0.05) ability to inhibits lipid peroxidation in both the basal and pro-oxidants induced lipid peroxidation in rat`s brain homogenates in vitro. Therefore, it is very obvious from the study that both teas had high antioxidant and neuroprotective potentials, however, green tea had higher neuroprotective potentials, which may probably be due to its high total phenol content, reducing power, Fe2+ chelating ability, OH* and NO* scavenging ability, as well as inhibition of over-stimulation of NMDA receptor.
PDF Abstract XML References Citation
How to cite this article
Ganiyu Oboh, 2009. The Neuroprotective Potentials of Sour (Hibiscus sabdariffa,Calyx) and Green (Camellia sinensis) Teas on Some Pro-Oxidants Induced Oxidative Stress in Brain. Asian Journal of Clinical Nutrition, 1: 40-49.
DOI: 10.3923/ajcn.2009.40.49
URL: https://scialert.net/abstract/?doi=ajcn.2009.40.49
DOI: 10.3923/ajcn.2009.40.49
URL: https://scialert.net/abstract/?doi=ajcn.2009.40.49
INTRODUCTION
During the last decade, evidences abound to show that the free radical- mediated lipid peroxidation plays a crucial and causative role in the pathogenesis of many neurodegenerative diseases and atherosclerosis. In living organisms, various Reactive Oxygen Species (ROS) can be form in different ways. Normal aerobic respiration could stimulate polymorphonuclear leukocytes and macrophages and peroxisomes appear to be the main endogenous sources of most of the oxidants produced by cells. Exogenous sources of ROS include tobacco smoke, certain pollutants, organic solvents and pesticides (Oboh et al., 2007). ROS can cause lipid peroxidation in foods, which leads to the deterioration of the food (Gulcin et al., 2003). It can induce oxidative damage on biomolecules like lipids, nucleic acids, proteins and carbohydrates; these damage causes ageing, cancer and many other diseases (Gulcin et al., 2003). ROS have been implicated in more than 100 diseases, including malaria, acquired immunodeficiency syndrome, heart disease, stroke, arteriosclerosis, diabetes and cancer (Gulcin et al., 2003).
The brain and nervous system are vulnerable to oxidative stress due to limited antioxidant capacity (Vega-Naredo et al., 2005). Neurons can not synthesize glutathione, a fundamental component of aerobic cell antioxidant machinery, but instead rely on surrounding astrocyte cells to provide useable glutathione precursors (Peng et al., 2007). Because the brain has limited access to the bulk of antioxidants produced by the body, neurons are the first cells to be affected by a shortage of antioxidants and are most susceptible to oxidative stress (Peng et al., 2007; Oboh et al., 2007). The human body is equipped with an antioxidant defense system that deactivates these highly reactive free radicals, this includes antioxidants enzymes (made in the body) and antioxidant nutrients (found in foods), that soaks up all the excess reactivity that these free radicals have, turning them to harmless particles that can be get rid of. However, recent studies have revealed that one of the practical way through which the activity of free radicals could be managed in the body is through dietary means. Dietary antioxidants may play an important role in protecting the cell against damage caused by free radicals, by acting as radical scavengers, reducing agents, forming complexes with pro-oxidant metals and quenchers of singlet oxygen formation (Prior, 2003; Oboh, 2005; Oboh et al., 2007).
Green and sour teas are popularly consumed in Asia and Africa and some western countries because of the numerous medicinal attributes ascribed to them. Hibiscus sabdariffa Calyx, called karkade in Switzerland, a name not too different from the Arabic is used in jams, jellies, sauces and wines. In the West Indies and elsewhere in the tropics the fleshy calyces are used fresh for making roselle wine, jelly, syrup, gelatin, refreshing beverages, pudding and cakes and dried roselle is used for tea, jelly, marmalade, ices, ice-cream, sherbets, butter, pies, sauces, tarts and other desserts. Oboh and Elusiyan (2004) reported the usefulness of roselle in arteriosclerosis and as an intestinal antiseptic. The drink made by placing, the calyx in water, is said to be a folk remedy for cancer. While green tea is made from unfermented leaves of Camellia sinensis and is reputed to contain the highest concentration of polyphenols, chemicals that act as powerful antioxidants. Green tea has been use in the management of many diseases such as atherosclerosis, breast cancer, colorectal cancer, Crohn`s diseases, diabetes mellitus, hypercholesterolemia, lung cancer, prostate cancer, ulcerative colitis and wounds.
Recently, it was reported that intact anthocyanins (a phenolic pigment abundant in sour and green tea) reached the brain within minutes from their introduction into the stomach (Passamonti et al., 2005). However, there are dearths of information on the ability of these teas to prevent oxidative stress associated with the following neurotoxins: Fe (II), sodium nitroprusside and quinolinic acid in the brain. The present study therefore sought to compare the ability of those teas to prevent the various pro-oxidants induced lipid peroxidation in isolated rat`s brain in vitro.
MATERIALS AND METHODS
Materials:
Green (Camellia sinensis) and sour (Hibiscus sabdariffa, Calyx) teas were purchased from a supermarket in Camobi, Santa Maria RS, Brazil. They were authenticated in Departamento de Biologia, Universidade Federal de Santa Maria, Santa Maria RS, Brazil and the study was carried out from March to December, 2005. All the chemicals used were analytical grade, while the water was glass distilled. The handling and the use of the animal were in accordance with NIH Guide for the care and use of laboratory animals. In this experiment Wistar strain albino rats weighing 200-230 g were used, these were collected from the breeding colony of Departamento de Quimica, Universidade Federal de Santa Maria, Santa Maria RS, Brazil. They were maintained at 25°C, on a 12 h light/12 h dark cycle, with free access to food and water.
Aqueous Extract Preparation
The aqueous extract of the green and sour teas were prepared in hot water, briefly about 1 g of each of the tea were soaked in 100 mL hot water for 5 min. Thereafter the mixtures were centrifuged at 2000 rpm for 10 min. The supernatant was used for the determination of total phenol content, antioxidant activity (reducing power, Fe (II) chelating ability and OH radical scavenging ability) and lipid peroxidation bioassay.
Total Phenol Determination
The total phenol content was determined by adding 0.5 mL extract to 2.5 mL 10% Folin-Ciocalteu`s reagent (v/v) and 2.0 mL 7.5% sodium carbonate was added. The mixture was incubated at 45°C for 40 min and the absorbance was measured at 765 nm in a spectrophotometer using gallic acid as a standard (Singleton et al., 1999).
Fe2+ Chelation Assay
The ability of the tea to chelate Fe2+ was determined using the method reported by Puntel et al. (2005). Freshly prepared 500 μM FeSO4 (150 μL) was added to a reaction mixture containing 168 μL 0.1M Tris-HCl (pH 7.4), 218 μL saline and the aqueous extract of the teas (0-25 μL). The reaction mixture was incubated for 5 min, before the addition of 13 μL 0.25% 1, 10-phenanthroline (w/v). The absorbance was subsequently measured at 510 nm in a spectrophotometer.
OH Radical Scavenging Ability
The ability of the extract of the teas to prevent Fe2+/H2O2 induced decomposition of deoxyribose was carried out using the method of Halliwell and Gutteridge (1981). Briefly, freshly prepared aqueous extract (0-100 μL) was added to a reaction mixture containing 120 μL 20 mM deoxyribose, 400 μL 0.1M phosphate buffer, 40 μL 20 mM hydrogen peroxide and 40 μL, 500 μM FeSO4 and the volume were made to 800 μL with distilled water. The reaction mixture was incubated at 37°C for 30 min and the reaction was stop by the addition of 0.5 mL of 2.8% TCA (Trichloroacetic acid), this was followed by the addition of 0.4 mL of 0.6% TBA (Thiobarbituric acid) solution. The tubes were subsequently incubated in boiling water for 20 min. The absorbance was measured at measured at 532 nm in spectrophotometer.
Reducing Property
The reducing property of the teas were determined by assessing the ability of the extract to reduce a FeCl3 solution as described by Pulido et al. (2000), 2.5 mL aliquot was mixed with 2.5 mL 200 mM sodium phosphate buffer (pH 6.6) and 2.5 mL 1% potassium ferricyanide. The mixture was incubated at 50°C for 20 min. and then 2.5 mL 10% trichloroacetic acid was added. This was then centrifuged at 650 rpm for 10 min. Five milliliter of the supernatant was mixed with an equal volume of water and 1 mL 0.1% ferric chloride. The absorbance was measured at 700 nm and a higher absorbance indicates a greater reducing power.
Preparation of Brain Homogenates
The rats were decapitated under mild diethyl ether anaesthesia and the cerebral tissue (whole brain) was rapidly dissected and placed on ice and weighed. This tissue was subsequently homogenized in cold saline (1/10 w/v) with about 10-up and down strokes at approximately 1200 rev min-1 in a Teflon glass homogenizer. The homogenate was centrifuge for 10 min at 3000 x g to yield a pellet that was discarded and a low-speed supernatant (S1) containing mainly water, proteins, lipids (cholesterol, galactolipid, individual phospholipids, gangliosides), DNA and RNA that was kept for lipid peroxidation assay (Belle et al., 2004).
Lipid Peroxidation and Thiobarbibutric Acid Reactions
The lipid peroxidation assay was carried out using the modified method of Ohkawa et al. (1979) S1 fraction (100 μL) was mixed with a reaction mixture containing 30 μL of 0.1M Tris-HCl buffer (pH 7.4), tea extract (0-100 μL) and 30 μL of the pro-oxidant solution (250 μM freshly prepared FeSO4, 70 μM sodium nitroprusside and 1 mM quinolinic acid). The volume was made up to 300 μL with water before incubation at 37°C for 1 h. The colour reaction was developed by adding 300 μL 8.1% SDS (sodium dodecyl sulphate) to the reaction mixture containing S1, this was subsequently followed by the addition of 600 μL of acetic acid/HCl (pH 3.4) and 600 μL 0.8% TBA (Thiobarbituric acid). This mixture was incubated at 100°C for 1 h. TBARS (Thiobarbituric acid reactive species) produced were measured at 532 nm.
Analysis of Data
The results were expressed as mean±Standard Error (SE) and a student t-test was carried out (Zar, 1984). Significance was at p ≤ 0.05. The EC50 (extract concentration causing 50% inhibition of lipid peroxidation in the Rat`s brain) for the tea extracts was determined.
RESULTS AND DISCUSSION
The results of the study revealed that both teas had high total phenol content [green (24.5 mg g-1), sour (13.3 mg g-1)]. It is worth noting that the total phenol content of these teas were generally higher than that of some commonly consumed green leafy vegetables in Nigeria such as Telfairia occidentalis, Struchium sparganophora, solanum macrocarpon, Vernonia amygdalina, Ocimum gratissimum, Amaranthus cruentus and Cnidoscolus acontifolus (Oboh, 2005) and those reported by Chu et al. (2002). Also it was found that the total phenols of these teas were higher than that of some commonly consumed fruits (Sun et al., 2002) and those of hot and sweet pepper (Oboh et al., 2007; Oboh and Rocha, 2007; Maran et al., 2004; Materska and Perucka, 2005). However, the total phenol content of both teas were lower than that of green tea and black tea earlier reported by Lee and Lee (Lee and Lee, 2002), the reason for the difference in the total phenol content cannot be categorically stated, however it will not far fetched from location difference in the cultivation of the green tea, as well as difference in the mode and the condition under which the tea extracts were prepared. Nevertheless, the relatively high total phenol content of these teas may have accounted for the use of these teas in the management of degenerative diseases such as cancer and hypertension. Nevertheless, green tea had a significantly higher (p<0.05) total phenol content (almost twice) than the sour.
The ability of the teas to prevent lipid peroxidation in basal rat brain tissue homogenates are shown in Fig. 1. The results revealed that both teas inhibited the production of malondialdehyde (MDA) in the cultured brain tissues in a dose-dependent manner. However, green tea caused a significantly (p<0.05) higher inhibition in the lipid peroxidation in the isolated brain tissues than the sour tea. The EC50 of the green tea (0.41 mg mL-1) was significantly (p<0.05) lower than that of sour tea (0.70 mg mL-1). Furthermore, it was obvious from the study that there was an agreement between the total phenol content and the inhibition of lipid peroxidation in the isolated rat brain by both teas (Fig. 1). Green tea with higher total phenol content caused higher inhibition of lipid peroxidation than sour tea. This goes a long way to confirm a possible correlation between the antioxidant activity of this teas and their total phenol content. It is also worth noting that the EC50 of both teas on isolated rat`s brain homogenates was far below that of aqueous extracts of hot peppers on isolated rat`s brain tissue in vitro (Oboh et al., 2007).
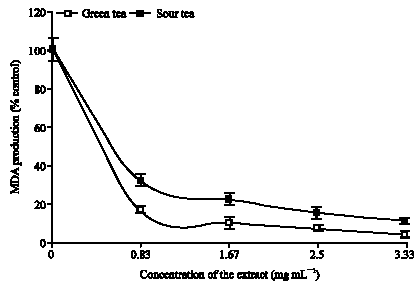 | |
| Fig. 1: | Inhibition of lipid peroxidation in Rat`s brain (in vitro) by green and sour tea |
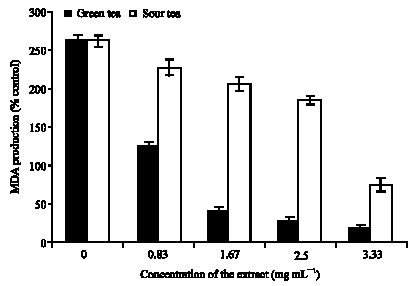 | |
| Fig. 2: | Inhibition of Fe (II) induced lipid peroxidation in Rat`s brain (in vitro) by green and sour tea |
As shown, incubation of the brain tissues in the presence of 25 μM Fe (II) increased the malondialdehyde (MDA) produced in the isolated brain tissue homogenates (260%) when compared with that of the basal brain tissue homogenates (100%) (Fig. 2). These findings agree with our earlier reports (Oboh et al., 2007; Oboh and Rocha, 2007); in that Fe (II) is a very potent pro-oxidant. The increased lipid peroxidation in the presence of Fe2+ could be attributed to the fact that, Fe2+ can catalyze one-electron transfer reactions that generate reactive oxygen species, such us the reactive OH*, which is formed from H2O2 through the Fenton reaction. Iron also decomposes lipid peroxides, thus generating peroxyl and alkoxyl radicals, which favors the propagation of lipid oxidation (Oboh et al., 2007). Nevertheless, both teas at the concentration range tested (0.83-3.33 mg mL-1) caused a significant decrease (p<0.05) in the MDA produced in the Fe (II) induced lipid peroxidation in the isolated rats brain homogenates in a dose-dependent manner [green tea (17.7-124.5%), sour tea (74.4-224.5%)]. The reason for the inhibition of the lipid peroxidation in the brain by both teas cannot be categorically stated; however, it will not be far fetch from the possibility that the phytochemical in the teas could have form complexes with the Fe (II) thereby preventing them from catalyzing the initiation of lipid peroxidation, or/and the possibility that the phytochemical (phenol) could have scavenge the OH* radical produced by the Fe (II) catalyzed reaction (Oboh et al., 2007).
However, green tea caused a significantly (p<0.05) higher inhibition in the Fe (II) induced lipid peroxidation in the rat`s brain than sour tea; the reason for the higher inhibitory properties of the green tea may not be far fetch from that fact that, green tea has higher total phenol content than sour tea. Moreover polyphenol had been reported to be able to form complexes with metals and scavenge free radicals (Oboh et al., 2007; Oboh and Rocha, 2007). In order, to explain the mechanism through which the teas inhibits the Fe (II) induced lipid peroxidation, the Fe (II) chelating ability and OH* scavenging ability of both teas were determined.
As shown in Fig. 3, both teas significantly chelate Fe (II) at 0.25-1.00 mg mL-1 concentration of the teas, in a dose-dependent manner. This high ability of the teas to form complexes with Fe (II) may have caused a significant reduction in the available Fe (II) that will be required to catalyze the production of free radicals in order to initiates lipids peroxidation in the tissues. This high Fe (II) chelating ability may have contributed immensely to the ability of the extract to prevent Fe (II) induced lipid peroxidation in the isolated rat`s brain homogenates. However, green tea (53.3-82.0%) had a significantly (p<0.05) higher Fe (II) chelating ability than sour tea (34.2-52.4%), this higher Fe (II) chelating ability of the green tea may have accounted for its higher inhibition of Fe (II) induced lipid peroxidation in isolated rat`s brain homogenates. However, the higher Fe (II) chelating ability of the green tea may not be far fetch from its higher total phenol content. Phenol had been reported to form complexes with Fe (II), thereby making them unavailable for the initiation of lipid peroxidation (Oboh et al., 2007; Oboh and Rocha, 2007).
The ability of the aqueous extract to inhibits Fe (II)/H2O2-induced decomposition of deoxyribose as an index for hydroxyl radical (OH*) scavenging ability is shown in Fig. 4. The results revealed that both extracts had high OH* scavenging ability at the concentration of the extract tested (0.25 to 1.00 mg mL-1), the extracts were able to inhibit deoxyribose decomposition in a dose dependent manner. However, aqueous extract of green tea had a significantly higher (p<0.05) OH* scavenging ability than that of the sour tea. This higher hydroxyl radical scavenging of the green tea (in addition to high Fe (II) chelating ability) may have accounted for its higher inhibition of Fe (II) induced lipid peroxidation in isolated rats` brain when compared to the sour tea.
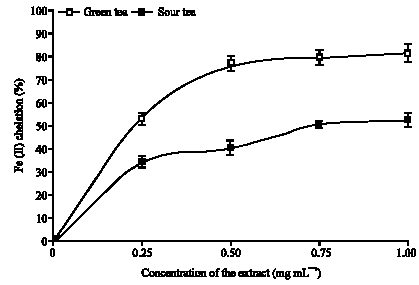 | |
| Fig. 3: | Fe (II) chelating ability of green and sour tea |
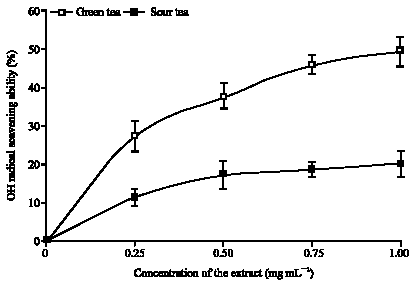 | |
| Fig. 4: | OH radical scavenging ability of green and sour teas |
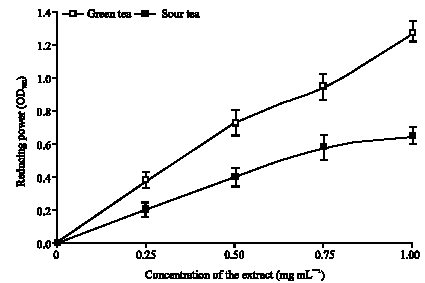 | |
| Fig. 5: | Reducing power of green and sour teas |
The results revealed that both teas were able to reduce Fe3+ to Fe2+ in a dose-dependent manner. However, green tea had a significantly higher (p<0.05) ability to reduce Fe3+ to Fe2+ than sour tea (Fig. 5). The high phenol content, reducing power, Fe (II) chelating and OH* radical scavenging ability may have accounted for the high ability of the teas to inhibit Fe (II) induced lipid peroxidation in the rats brain in vitro. However, the significantly higher total phenol, reducing power, Fe (II) chelating and OH* radical scavenging ability of green tea may have accounted for it significantly higher (p<0.05) ability to inhibits the production of MDA in isolated rat`s brain homogenates-in vitro than sour tea.
The results of the interaction of the aqueous extract of both teas with sodium nitroprusside induced lipid peroxidation in rats brain homogenates are shown in Fig. 6. The results revealed that incubation of the isolated rats brain homogenates in the presence of 7 μM sodium nitroprusside caused a significant increase (p<0.05) in the malondialdehyde (MDA) (222.6%) content of the isolated brain compared to the unstressed brain (100%), this result agrees with earlier reports on the interaction of sodium nitroprusside with isolated rat`s brain (Oboh and Rocha, 2007). Sodium Nitroprusside (SNP) has been reported to cause cytotoxicity through the release of cyanide and/or Nitric Oxide (NO) (Belle et al., 2004; Posser et al., 2006; Oboh and Rocha, 2007). NO is a free radical with a short half-life (< 30 sec). Although NO acts independently, it also may cause neuronal damage in cooperation with other Reactive Oxygen Species (ROS) such as superoxide radical to form peroxynitrite radical (Belle et al., 2004; Posser et al., 2006; Oboh and Rocha, 2007). In addition, the Fe produced from the decomposition of the sodium nitroprusside could also sustain the lipid peroxidation, by initiating the production of OH radical through Fenton`s reaction (Belle et al., 2004; Puntel et al., 2005; Oboh and Rocha, 2007).
However, it is quite interesting to note that both teas significantly (p<0.05) inhibited 7 μM sodium nitroprusside induced lipid peroxidation in rat`s brain in a dose-dependent manner. This clearly indicates that both teas were able to scavenge the NO* produced by sodium nitropusrusside and chelate and/or scavenge the Fe and OH* produced as a result of the decomposition of the sodium nitroprusside. However, green tea had a significantly higher (p<0.05) ability to inhibits sodium nitroprusside induced lipid peroxidation in the isolated rat`s brain. This clearly indicates that green tea have higher NO* scavenging ability, in addition to its higher reducing power, Fe (II) chelating ability and OH* scavenging ability than the sour tea.
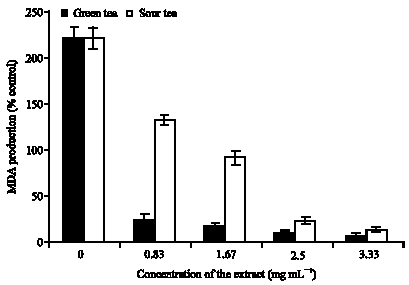 | |
| Fig. 6: | Inhibition of Sodium nitroprusside induced lipid peroxidation in Rat`s brain (in vitro) by green and sour tea |
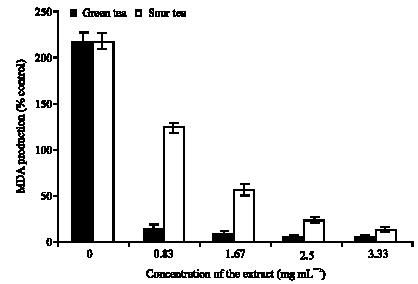 | |
| Fig. 7: | Inhibition of Quinolinic acid induced lipid peroxidation in Rat`s brain (in vitro) by green and sour tea |
Furthermore, incubation of the isolated rats brain in the presence of quinolinic acid caused a significant increase (p<0.05) in the MDA content of the rats brain (215%) when compared with the basal brain tissues (100%) (Fig. 7). Quinolinic Acid (QA) is a neuroactive metabolite of the tryptophan-kinurenine pathway, which can be produced by macrophages and microglia (Cammer, 2000). It is present in both the human and rat brain and it has been implicated in the pathogenesis of a variety of human neurological diseases (Belle et al., 2004; Oboh and Rocha, 2007). Quinolinic acid is recognized pharmacologically as an endogenous glutamate agonist with a relative selectivity for the NMDA receptor in the brain (Vega-Naredo et al., 2005). Since it is not readily metabolized in the synaptic cleft, it stimulates the NMDA receptor for prolonged periods. This sustained stimulation results in opening of calcium channels causing Ca2+ influx followed by Ca2+-dependent enhancement of free radical production leading to molecular damage and often to cell death (Cabrera et al., 2000).
However, aqueous extracts of green and sour teas were able to inhibit quinolinic acid induced lipid peroxidation in rat`s brain in a dose dependent manner. The inhibition of quinolinic acid induced lipid peroxidation by both teas clearly indicates that, in addition to the fact those teas had free radical scavenging ability, they were able to prevent quinolinic acid induced over-stimulation of NMDA receptors that leads Ca2+ influx that results in free radical production that could cause eventual brain damage. Nevertheless, aqueous extract of green tea had a significantly higher (p<0.05) inhibitory effect on quinolinic acid induced lipid peroxidation in the rat`s brain than the sour tea. A possible explanation for the higher inhibitory effect of green tea is that, in addition to its higher radical scavenging ability, it may have had higher ability to prevent over-stimulation of NMDA receptor than sour tea.
CONCLUSION
In conclusion, aqueous extract of green and sour teas were able to inhibit various pro-oxidants (Fe2+, sodium nitroprusside and quinolinic acid) induced lipid peroxidation in isolated rat`s brain homogenates-in vitro. However, green tea had a significantly higher (p<0.05) ability to prevents the various pro-oxidants induced lipid peroxidation in rat`s brain. The higher ability of the green tea to protect the brain from oxidative stress could be attributed to its higher total phenol content, Fe2+ chelating ability, OH* and NO* scavenging ability, as well as inhibition of over-stimulation of NMDA receptor.
ACKNOWLEDGMENTS
The assistance from Prof. J.B.T. Rocha and Post-doctoral fellowship from Alexander von Humboldt (AvH) is graciously acknowledged during which period the manuscript was written.
REFERENCES
- Belle, N.A., G.D. Dalmolin, G. Fonini, M.A. Rubin and J.B. Rocha, 2004. Polyamines reduces lipid peroxidation induced by different pro-oxidant agents. Brain Res., 1008: 245-251.
CrossRefPubMedDirect Link - Cabrera, J., R.J. Reiter, D.X. Tan, W. Qi and R.M. Sainz et al., 2000. Melatonin reduces oxidative neurotoxicity due to quinolinic acid: In vitro and in vivo findings. Neuropharmacology, 39: 507-514.
CrossRefDirect Link - Chu, Y.F., J. Sun, X. Wu and R.H. Liu, 2002. Antioxidant and antiproliferative activities of common vegetables. J. Agric. Food Chem., 50: 6910-6916.
CrossRefPubMedDirect Link - Gulcin, I., M. Oktay, E. Kirecci and O.I. Kufrevioglu, 2003. Screening of antioxidant and antimicrobial activities of anise (Pimpinella anisum L.) seed extracts. Food Chem., 83: 371-382.
CrossRefDirect Link - Halliwell, B. and J.M.C. Gutteridge, 1981. Formation of a thiobarbituric-acid-reactive substance from deoxyribose in the presence of iron salts: The role of superoxide and hydroxyl radicals. FEBS Lett., 128: 347-352.
CrossRefPubMedDirect Link - Lee, K.W., H.J. Lee and C.Y. Lee, 2002. Antioxidant activity of black tea vs. green tea. J. Nutr., 132: 785-786.
PubMedDirect Link - Marin, A., F. Ferreres, F.A. Tomas-Barberan and M.I. Gil, 2004. Characterization and quantitation of antioxidant constituents of sweet pepper (Capsicum annuum L.). J. Agric. Food Chem., 52: 3861-3869.
CrossRefDirect Link - Materska, M. and I. Perucka, 2005. Antioxidant activity of the main phenolic compounds isolated from hot pepper fruit (Capsicum annuum L.). J. Agric. Food Chem., 53: 1750-1756.
CrossRefPubMedDirect Link - Oboh, G. and C.A. Elusiyan, 2004. Nutrient composition and antimicrobial activity of Sorrel drinks (Soborodo). J. Med. Food, 7: 340-342.
CrossRefPubMedDirect Link - Oboh, G., 2005. Effect of blanching on the antioxidant property of some tropical green leafy vegetables. LWT-Food Sci. Technol., 38: 513-517.
CrossRefDirect Link - Oboh, G. and J.B.T. Rocha, 2007. Polyphenols in red pepper [Capsicum annuum var. aviculare (Tepin)] and their protective effect on some pro-oxidants induced lipid peroxidation in brain and liver. Eur. Food Res. Technol., 225: 239-247.
CrossRefDirect Link - Oboh, G., R.L. Puntel and J.B.T. Rocha, 2007. Hot pepper (Capsicum annuum, Tepin and Capsicum chinese, Habanero) Prevents Fe2+-induced lipid peroxidation in brain-in vitro. Food Chem., 102: 178-185.
CrossRefDirect Link - Ohkawa, H., N. Ohishi and K. Yagi, 1979. Assay for lipid peroxides in animal tissues by thiobarbituric acid reaction. Anal. Biochem., 95: 351-358.
CrossRefPubMedDirect Link - Passamonti, S., U. Vrhovsek, A. Vanzo and F. Mattivi, 2005. Fast access of some grape pigments to the brain J. Agric. Food Chem., 53: 7029-7034.
PubMedDirect Link - Peng, J., L. Peng, F.F. Stevenson, S.R. Doctrow and J.K. Andersen, 2007. Iron and paraquat as synergistic environmental risk factors in sporadic Parkinson's disease accelerate age-related neurodegeneration J. Neurosci., 27: 6914-6922.
CrossRefPubMedDirect Link - Posser, T., M.B. Moretto, A. Luiz Dafre, M. Farina and J.B.T. Rocha et al., 2006. Antioxidant effect of diphenyl diselenide against sodium nitroprusside (SNP) induced lipid peroxidation in human platelets and erythrocyte membranes: An in vitro evaluation. Chem. Biol. Interact., 164: 126-135.
CrossRefPubMedDirect Link - Prior, R.L., 2003. Fruits and vegetables in the prevention of cellular oxidative damage. Am. J. Clin. Nut., 78: 570S-578S.
Direct Link - Pulido, R., L. Bravo and F. Saura-Calixto, 2000. Antioxidant activity of dietary polyphenols as determined by a modified ferric reducing/antioxidant power assay. J. Agric. Food Chem., 48: 3396-3402.
CrossRefPubMedDirect Link - Puntel, R.L., C.W. Nogueira and J.B.T. Rocha, 2005. Krebs cycle intermediates modulate Thiobarbituric Acid Reactive Species (TBARS) production in rat brain in vitro. Neurochem. Res., 30: 225-235.
CrossRefPubMedDirect Link - Singleton, V.L., R. Orthofer and R.M. Lamuela-Raventos, 1999. Analysis of Total Phenols and Other Oxidation Substrates and Antioxidants by Means of Folin-Ciocalteu Reagent. In: Methods in Enzymology, Burslem, G.L. (Ed.), Academic Press, Cambridge, Massachusetts, ISBN: 9780121822002, pp: 152-178.
CrossRefDirect Link - Sun, J., Y.F. Chu, X. Wu and R.H. Liu, 2002. Antioxidant and antiproliferative activities of common fruits. J. Agric. Food Chem., 50: 7449-7454.
CrossRefPubMedDirect Link - Vega-Naredo, I., B. Poeggeler, V. Sierra-Sanchez, B. Caballero and C. Tomas-Zapico et al., 2005. Melatonin neutralizes neurotoxicity induced by quinolinic acid in brain tissue culture. J. Pineal Res., 39: 266-275.
CrossRefPubMedDirect Link - Zar, J.H., 1984. Biostatistical Analysis. 2nd Edn., Prentice-Hall Inc., Englewood Cliffs, New Jersey, USA, Pages: 718.
Direct Link