
Tewodros Kumssa
Department of Zoological Sciences, Addis Ababa University, P.O. Box 1176, Addis Ababa, Ethiopia
LiveDNA: 251.33319
Afework Bekele
Department of Zoological Sciences, Addis Ababa University, P.O. Box 1176, Addis Ababa, Ethiopia
Asian Journal of Biological Sciences
Year: 2014 | Volume: 7 | Issue: 2 | Page No.: 57-65







ABSTRACT
ASLNP wetlands are important feeding sites of lesser flamingos (Phoeniconaias minor). It is located in the Great Rift Valley. Flamingos constitute the major content of the bulk of lakes. However, studies to determine food preferences of flamingoes in Ethiopia using crop content analyses are lacking. The analysis of diets examined during 2011-13 at two major feeding sites of Lakes Abijata and Chitu. Bacillariophyceae (diatoms) and Cyanophyceae (blue green algae) constituted the main diet of lesser flamingos in Lake Abijata. Nine diatom genera were identified in the crop contents of the bird. Diatoms had the highest percentage abundance (90.1-95.5%). These were followed by sand particles and detritus (3.9-9.16%) and Cyanophytes (0.23-0.87%) and Navicula was always the most common genus and accounted for 94.17 % of the average total algal density. The analysis of diet from the two flamingos in Lake Chitu had clearly shown that only one species Arthrospira fusiformis was dominating. There were insignificant variations on morphometric features of birds within and between lake birds. The distribution and abundance of flamingos are related to food supply. Thus, changes in the numbers of flamingos at a particular lake during the year and between years may reflect fluctuations in the availability of food supply. Lesser flamingos are specialized feeding on diatoms in Lake Abijata and Spirulina, in Lake Chitu. Presently, there is no effective protection of flamingo feeding areas or enforcement of laws protecting the bird.
PDF Abstract XML References Citation
Received: December 22, 2013;
Accepted: March 08, 2014;
Published: May 21, 2014
How to cite this article
Tewodros Kumssa and Afework Bekele, 2014. Feeding Ecology of Lesser Flamingos (Phoeniconaias minor) in Abijata-Shalla Lakes National Park (ASLNP) with Special Reference to Lakes Abijata and Chitu, Ethiopia. Asian Journal of Biological Sciences, 7: 57-65.
URL: https://scialert.net/abstract/?doi=ajbs.2014.57.65
URL: https://scialert.net/abstract/?doi=ajbs.2014.57.65
INTRODUCTION
Food availability is one of the most important factors determining bird distributions (Evans and Dugan, 1984). Moreover, habitat structure influences water bird foraging behavior by means of food availability and the energetic constraints of obtaining it and associated with the adaptation potential of a species to a specific habitat type in terms of its morphology (Gatto et al., 2008). Foraging habitat use and feeding methods are therefore important factors involved in resource partitioning and can be used to assign foraging guilds to birds (Weller, 1999).
Flamingos are morphologically adapted to exploit the microscopic food resources from these ecosystems via filter feeding (Vareschi, 1978). They separate food items from water and unwanted particles using a large, fatty, highly sensitive tongue with numerous fleshy spines complemented by a keeled, lamellate bill (Jenkin, 1957). Water flow is produced by the large, powerful tongue which forces water into and out of the bill and ingest the food using the typical inertial throw-and-catch behavior of most birds (Zweers et al., 1995).
Flamingos are the principal primary consumers of the prolific production in the alkaline lakes (Vareschi, 1978; Tuite, 1979). Thses lakes are characterized by simple community structures where few tolerant and adapted species attain high population densities (Vareschi and Jacobs, 1985). The two flamingo species differ in their food preference. The greater flamingo is a generalist that consumes small planktonic and benthic animals such as molluscs and arthropods, as well as mud and algae, the lesser flamingo is highly specialized as on cyanobacteria and algae (Jenkin, 1957; Brown, 1973).
Feeding in flocks is a widespread behavior among animals, especially birds like flamingo (McCulloch and Borello, 2000). How foraging birds may increase their fitness by joining flocks in spite of increased costs due to higher detectability and competition has stirred intense debates (Terborgh et al., 1990).
Flamingos wade (or swim if necessary) to sieve microorganisms from the water. Foraging mostly occurs in shallow waters near the banks of the lakes (Snow and Perrins, 1998). Foraging habitat depths range from 2-50 cm, according to the maximum length of a bird's leg (Mascitti and Castanera, 2006). The species forages both by day and night (Brown et al., 1982).
At present a new feeding ground for lesser flamingos was developed at Lake Oloidien, a former bay of Lake Naivasha in Kenya. The negative effects of overexploitation and degradation of Lake Naivasha freshwater ecosystem (Harper et al., 2011) has been accompanied by a positive development of a new flamingo lake. The ambivalent development characterized by increasing salinity and a switch in the phytoplankton composition created favorable conditions for the flamingos (Krienitz and Kotut, 2010). According to Brown (1973), an adult flamingo with 1.8 kg weight has a minimum daily food requirement of about 180 g Fresh Weight (FW) (= 21.2 g Dry Weight (DW).
Long-term monitoring of bird populations, together with detailed studies on individual species is very important. According to Edwards (1996), investigations on avian fauna in Ethiopia are negligible. Detailed studies have not been carried out. Only few researchers have conducted studies on the diversity and ecology of avian species in some parts of Ethiopia (Ash and Gullick, 1989). For instance, the feeding ecology of the flamingo in Ethiopia has not been well known. They occur in some wetland areas of the country. Published data are unavailable on the food and feeding habits of flamingos in Ethiopia which constitute the major content of the bulk of Lakes Abijata, Shalla and Chitu. Studies to determine food preferences of flamingoes using crop content analyses are lacking.
MATERIALS AND METHODS
Study area: Abijata-Shalla Lakes region was established as a National Park by the Ethiopian Wildlife Conservation Organization in 1970 with the aim of conserving the biodiversity of the spectacular number of aquatic birds (Hillman, 1993). The park is known as Abijata-Shalla Lakes National Park (ASLNP) by deriving its name from the two lakes Abijata and Shalla (Edwards 1996). ASLNP is one of Ethiopia’s National Parks, located in the Great Rift Valley comprising three lakes: Abijata, Shalla and Chitu (Fig. 1). The specific site lies between the coordinates of 7°15'-7°45'N and 38°30'-38°45'E; at about 207 km south from Addis Ababa. ASLNP comprises two types of ecosystems namely the water part and land together covering a total area of 887 km2 of which 405 km2 is land area while 482 km2 is water body (Senbeta and Tefera, 2001).
Among the three saline lakes the National Park has Lake Shalla is deepest (266 m deep) and Abijata is shallowest less than (7 m deep) (Ayenew, 2004). Lake Chitu is the smallest has an area of 0.8 km2 and a maximum depth of 21 m and is highly saline.
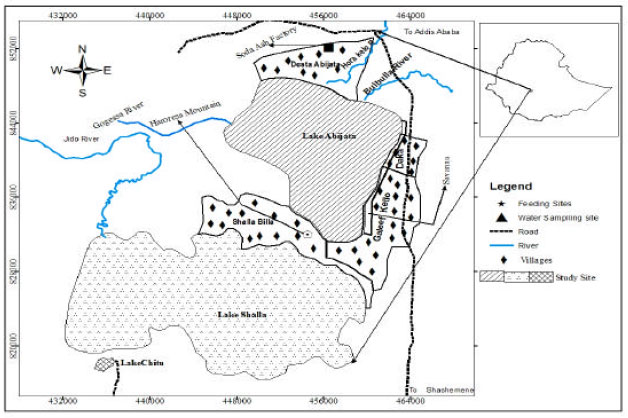 | |
| Fig. 1: | Map of the study area with study sites |
Due to its location in a rainfall deficit area of the Rift Valley, ASLNP receives an annual rainfall ranging between 500 and 700 mm and mean monthly temperature varies from 18.5°C-24.6°C with mean annual temperature of 21°C (Senbeta and Tefera, 2001). The habitat surrounding the lakes in the park is generally dominated with tree species of Acacia and open scrub rocky slopes (Senbeta and Tefera, 2001). A total of 453 bird species have been recorded in the park. The park has 6 endemic species to Ethiopia (Hillman, 1993) and it possessed at least 144 and 292 water-associated and terrestrial bird species, respectively (Senbeta and Tefera, 2001). The study was carried out from 2011-2013.
Collections were made under authorization of the Ethiopian Wildlife Conservation Authority (scientific collection permit number WA31/202/2011). Birds were trapped with indigenous traps, i.e., noose traps made from nylon twine. Flamingos were caught by legs mostly with the noose which was operated during daytime in foraging areas. Potential trapping locations were identified by visiting sites a day before to know where flamingos regularly feed and rest during the day. Effort was made to finish placement of noose lines just before sunrise, when it was still dark, to keep the disturbance a minimum. The study of food habits of flamingos was carried out only on two of their main feeding lakes (Abijata and Chitu). All the trapping was done during the dry season since the area not suitable to set the trap in their feeding area of the bird in the wet season. Birds were trapped by placing the noose lines parallel and perpendicular to the shoreline at one or more locations along the shore where the flamingos walk and feed actively. The water should be sufficiently deep to obscure the loops from view. The specific depth varied depending upon where the flamingos have been walking and how high the loops float in the water in each particular situation. Markings were used on shores (e.g., arrows in sand) to indicate where the trap is located in the water, as flamingos are suspicious of markers in the water and avoid the areas around them (Childress et al., 2006). To analyze bird crop contents, 8 lesser flamingos were captured (6 from Lake Abijata and 2 from Lake Chitu). No flamingos were captured from Lake Shalla due to its depth. Representative individuals of birds in the habitat were captured while feeding and collected immediately for crop content analysis since for waders, a residence time in the crop is shorter (Dekinga and Piersma, 1993). Then the crop contents were immediately preserved in 5% formalin solution to minimize postmortem digestion and autolysis until further microscopic examination as adopted by Swanson and Bartonek (1970). The contents were brought to the Department of Zoological Sciences, Zoology Laboratory for diet analysis. To identify the crop contents qualitatively, a part of the contents was separated for further microscopic analysis. The separated samples for qualitative examinations were homogenised together with distilled water and were analyzed microscopically to identify and count all dietary taxa to the lowest taxonomic level possible (Peckarsky et al., 1990). The even distribution of samples was checked by the use of subsamples, two or more strips and repetitive subsamples from the same samples. Then a sub-sample was then taken using a teat pipette. This was placed in a Sedgewick Rafter cell which carries a volume of 1 ml. The food items were identified and enumerated under a compound microscope. A total of 40 grids were counted randomly and single cells and filaments were all considered as algal units (and not No. of cells). Photographs were taken by using microscope eye piece camera with universal attachment set. The algae count (algae ml-1) of their diet was calculated according to Hotzel and Croome (1999) and Wetzel and Likens (2000). The percentage abundance of each food item was also computed by multiplying the ratio of the number of a particular item in the crop to the total number of items in the crop by a hundred as adapted by Short et al. (1999).
Additionally, five common morphological measurements (mass, flattened wing length, total tarsus length, head length and culmen length) were measured in the field. Each bird was weighed by pasola spring balance to an accuracy of 1 g by tying the legs, to temporarily immobilizing the legs and wings and placing the wrapped bird into the scale. The length of wing, tarsus and culmen measured by ruler to the nearest of 1 mm. In the laboratory, total weight of the stomach contents was taken using a mettler 1600 balance to 0.01 g. All measurements were taken by the same observer, to minimize observer bias. To analyze the data stata version 12 software was used. ANOVA and Chi-square test were performed to find out statistically significant difference among various variables.
RESULTS
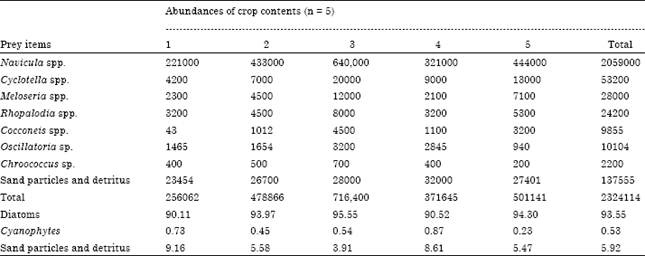
Information on diet of lesser flamingo was obtained from eight crop contents of adult lesser flamingos. Six were trapped from Lake Abijata and two from Lake Chitu. Among 6 dissected birds, one had empty crop content. Bacillariophyceae (diatoms) and Cyanophyceae (blue green algae) constituted the main diet of lesser flamingos in Lake Abijata. Nine diatom genera were identified in the crop contents of the bird. Among these, Rhoicosphenia sp., Thalassiosira sp., Cymbella spp. and Pinnlaria spp. were rare in their occurrence. The most common genera were represented mainly by Navicula spp., Cyclotella spp., Meloseria spp., Rhopalodia spp. and Cocconies spp. while blue green algae were represented by Oscillatoria sp. and Chroococcus sp. The percentage abundance of the major food items ingested by the birds is depicted in Table 1. Diatoms had the highest percentage abundance (90.1-95.5%). These were followed by sand particles and detritus (3.9-9.16%) and Cyanophytes (0.23-0.87%) and Navicula was always the most common genus and accounted for 94.17 % of the average total algal density. There were no zooplanktons recorded from all crops. Crop contents consisted varied diatom sizes with diatom of 55-60 μm length dominating.
| Table 1: | Composition of prey items ingested by lesser flamingos from lake abijata |
 | |
| Table 2: | Morphometric features of lesser flamingos (n = 8) |
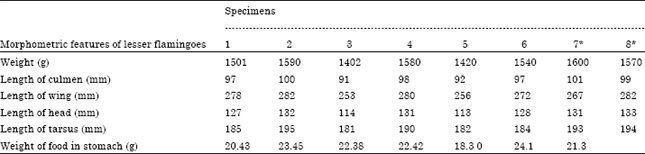 | |
| *Specimens from lake Chitu | |
There was a significant change (F416 = 18.11, p<0.05) in the proportion of diatom species in the crop contents during the investigation period. Navicula spp. and Cyclotella spp. were the dominant genera at all sampling periods. Species like Navicula is not only in high abundance in crop (Table 1) of the bird, they were also abundant in the identification of water samples.
Both Lake Chitu and Lake Abijata are the main feeding sites for lesser flamingos. Microscopic analysis of diet from the two flamingos in Lake Chitu had clearly shown that the species diversity of phytoplankton is very low compared to the flamingos of Lake Abijata, with only one species (Cyanophytes “spirulina”, Arthrospira fusiformis) with variable size dominating.
There were insignificant variations (F741 = 0.11, p>0.05) on morphometric features of birds within and between lake birds (Table 2). Flamingos are waders (long-legged birds that wade in water in search of food) and have webbed feet which produce discrete structures such as webbed footprints (~7 cm long, ~9 cm wide).
DISCUSSION
Analysis of crop contents during the study period showed the presence of two algal groups (diatoms and blue-green algae). The diatom and blue-green-algae taxa shown in (Fig. 2) were representative of the most common diet. Diatoms were the most preferable food of plant origin where they occurred in all of the examined birds in Lake Abijata. Lesser flamingoes were herbivorous since all their diet was phytoplankton. Benthic diatoms are the primary food choices.
 | |
| Fig. 2: | Microscopic photograph of the most common phytoplankton in diet of lesser flamingos in the two lakes |
Navicula was the dominant genus in their crop. The over-representation of this genus in the crop contents could reflect dietary preferences of certain algal species. Diatoms are thought to be a particularly important food in East Africa when Spirulina densities are unusually low (Tuite, 1981). Diatoms particularly Navicula are also taken in East Africa (Ridley et al., 1955) which was also found at Etosha.
Obtaining forage by filtering does not allow flamingos to actively select their prey. Selection is therefore made by size (Zweers et al., 1995). Prey selection primarily depends upon abundance, availability (distribution in the water column) and catchability (size) of prey (Madenjian and Gabray, 1995). The amount in the crop appears to be influenced by the relative abundance of food in the environment and the rate that the gizzard is able to process the food material consumed (Collins and Paggi, 1997). In Lake Abijata, the filtered diatoms have the size of 50 μm by 18 μm but quite frequently a diatom of 60 μm length is dominating. Analysis of the stomach contents of lesser flamingos revealed that the smallest particles filtered in large quantities were pinnate diatoms with size range from 15-70 μm (Jenkin, 1957). Andean flamingo feeds on benthic diatoms that range from 61-120 um (Mascitti and Kravetz, 2002). They are also known to consume mud from get nutrients. Examination of the diet of this species also showed that, there was high percentage of sand particles occurring in its crop which may facilitate digestion in the stomach.
The crop content analysis in Lake Chitu exclusively indicates a monospecific dominance of the Arthrospira fusiformes. The lamellae are equipped with rows of fringed platelets that have perfect size to collect the coiled filaments of Arthrospira fusiformes. Lesser flamingos are specialized consumers of Arthrospira fusiformes. Spirulina of Lake Chitu has an average size of 50-120 μm (length). The filtering device of lesser flamingo is very well adapted to the size of the Spirulina spirals which measure 40-200 μm by 20-50 μm. The platelets of the bill have an average distance of 120 μm, with 20 platelets per mm2 (Jenkin, 1957). Hence, the small size form of Arthrospira fusiformes from Chitu fits perfectly into the size range from practically trapped by filter apparatus of lesser flamingo.
Aggregations of phytoplankton in the stomach contents were observed. Many phytoplanktons were broken in the position of girdle bands. All food items except some few diatoms in the stomach were not sufficiently intact to insure accurate identification and enumeration. This may be due to the fast transfer of food items from the esophagus through the proventriculus to the gizzard. Most phytoplanktons broke down rapidly in the gizzard and made broken down beyond distinction.
In this study, the morphological components related to feeding (culmen length, head length and body weight), along with the components associated with movement (wing and tarsus length), showed insignificant variation among birds. Flamingos have the longest legs and necks relative to body size of any group of birds and are well known for their unusual bill shape (Del Hoyo et al., 1992). The bill is very large and sharply bent down near the middle of its length. Mean body weight of ASLNP flamingos are 1525 g. Bartholomew and Pennycuick (1973) obtained 1800 g. Brown (1973) stated an average weight of 1760 g. In comparison, flamingos of ASLNP were not in a very good nutritional state, 1525 g is considered slightly low. Besides the weight, the bill size is of special importance because it determines the pumping volume which in turn determines clearing and feeding rates. All of the measurements showed ASLNP flamingos lower than observed in Lake Nakuru by Vareschi, (1978). Flamingos have long legs and webbed feet help them to wade. They wade in water at depths between 2 and 50 cm depending on their leg length (Mascitti and Castanera, 2006).
CONCLUSION
Wetlands are very important natural areas. Millions of water birds depend on them. ASLNP lakes are one from the important wetland lakes were flamingos depend. The distribution and abundance of flamingos are related to food supply. Thus, changes in the numbers of flamingos at a particular lake during the year and between years may reflect fluctuations in the availability of food supply. Lesser flamingos are specialized feeding on diatoms in Lake Abijata and Spirulina, in Lake Chitu. The abundance and the quality of these algae at a particular site may attract large number of birds irrespective of the size of the lakes. At present, the dietary value of the different diatom species is still not well understood. So, studies should be conducted to identify the dietary value of on a range of diatom species in the lakes. Presently, there is no effective protection of flamingo feeding areas or enforcement of laws protecting the bird.
ACKNOWLEDGEMENT
We wish to thank Addis Ababa University and animal diversity research project for providing the required facilities for carrying out the above research study.
REFERENCES
- Bartholomew, G.A. and C.J. Pennycuick, 1973. The flamingo and pelican populations of the Rift Valley Lakes in 1968-69. Afr. J. Ecol., 11: 189-198.
CrossRef - Collins, P.A. and J.C. Paggi, 1997. Feeding ecology of Macrobrachium borelli (Nobili) (Decapoda: Palaemonidae) in the flood valley of the River Parana, Argentina. Hydrobiologia, 362: 21-30.
CrossRef - Dekinga, A. and T. Piersma, 1993. Reconstructing diet composition on the basis of faeces in a mollusc-eating wader, the knot Calidris canutus. Bird Study, 40: 144-156.
CrossRef - Gatto, A., F. Quintana and P. Yorio, 2008. Feeding behavior and habitat use in a waterbird assemblage at a marine wetland in coastal Patagonia, Argentina. Waterbirds, 31: 463-471.
CrossRef - Hillman, J.C., 1993. Ethiopia: Compendium of Wildlife: Conservation Information. New York Zoological Society (NYZS), Ethiopian Wildlife Conservation Organization, Addis Ababa, Ethiopia, Pages: 34.
Direct Link - Jenkin, P.M., 1957. The filter-feeding and food of flamingoes (Phoenicopteri). Philos. Trans. Roy. Soc. Lond. Ser. B Biol. Sci., 240: 401-493.
Direct Link - Krienitz, L. and K. Kotut, 2010. Fluctuating algal food populations and the occurrence of lesser flamingos (Phoeniconaias minor) in three kenyan rift valley lakes. J. Phycol., 46: 1088-1096.
CrossRef - Mascitti, V. and F.O. Kravetz, 2002. Bill morphology of South American flamingos. Condor, 104: 73-83.
CrossRefDirect Link - Mascitti, V. and M.B. Castanera, 2006. Foraging depth of flamingos in single-species and mixed-species flocks at Laguna de Pozuelos, Argentina. Waterbirds, 29: 328-334.
CrossRefDirect Link - McCulloch, G.P. and W. Borello, 2000. The importance of the Makgadikgadi salt pans in Botswana for flamingos in Africa. Waterbirds: Int. J. Waterbird Biol., 23: 64-68.
Direct Link - Short, J., M.C. Calver and D.A. Risbey, 1999. The impact of cats and foxes on the small vertebrate fauna of heirisson prong, Western Australia. I. Exploring potential impact using diet analysis. Wildl. Res., 26: 621-630.
CrossRefDirect Link - Swanson, G.A. and J.C. Bartonek, 1970. Bias associated with food analysis in gizzards of bluewinged teal. J. Wildl. Manage., 34: 739-746.
CrossRef - Ayenew, T., 2004. Environmental implications of changes in the levels of lakes in the Ethiopian Rift since 1970. Regional Environ. Change, 4: 192-204.
CrossRefDirect Link - Terborgh, J., S.K. Robinson, T.A. Parker, C.A. Munn and N. Pierpont, 1990. Structure and organization of an Amazonian forest bird community. Ecol. Monogr., 60: 213-238.
Direct Link - Tuite, C.H., 1979. Population size, distribution and biomass density of the lesser flamingo in the eastern Rift Valley, 1974-1976. J. Applied Ecol., 16: 765-775.
Direct Link - Tuite, C.H., 1981. Standing crop densities and distribution of Spirulina and benthic diatoms in East African alkaline saline lakes. Freshwater Boil., 11: 345-360.
CrossRef - Vareschi, E., 1978. The ecology of Lake Nakuru (Kenya). I. Abundance and feeding of the lesser flamingo. Oecologia, 32: 11-35.
Direct Link - Vareschi, E. and J. Jacobs, 1985. The ecology of Lake Nakuru (Kenya). VI. Synopsis of production and energy flow. Oecologia, 65: 412-424.
Direct Link - Zweers, G., F. De Jong, H. Berkhoudt and J.V. Berge, 1995. Filter feeding in flamingos (Phoenicopterus ruber). Condor, 97: 297-324.
Direct Link