
O.L. Adebayo
Department of Biochemistry, Olabisi Onabanjo University, Remo Campus, P.M.B. 2005, Ikenne, Ogun State, Nigeria
B. O. Adegbesan
Department of Biochemistry, Olabisi Onabanjo University, Remo Campus, P.M.B. 2005, Ikenne, Ogun State, Nigeria
G. A. Adenuga
Department of Biochemistry, Olabisi Onabanjo University, Remo Campus, P.M.B. 2005, Ikenne, Ogun State, Nigeria
Asian Journal of Biological Sciences
Year: 2009 | Volume: 2 | Issue: 1 | Page No.: 7-13







ABSTRACT
The comparative effect of Protein-Undernutrition (PU) and well-known neurotoxins on brain weight, lipid peroxidation and cellular antioxidants were investigated. The animals were randomly assigned to ten groups: A, B, C, D, E, F, G, H, I and J with five rats per group. Group A were placed on 16% casein diet while group B were placed on low protein diet (5% casein). Animals in groups C, E, G and I, received 200 ppm KCN, 80 mg (kg b.wt.)-1 PB, 75 mg (kg b.wt.)-1 DDT and 100 μ mole (Kg b.wt.)-1 Pb(NO3)2, respectively while groups D, F, H and J, served as their respective controls. Results show that KCN induced significant reduction in brain weight and SOD activity and significant increase (p<0.05) in CAT but no effect was observed in Lipid Peroxidation (LP) and GSH. DDT induced significant reduction (p<0.05) in CAT activity but did not affect other parameters investigated. Pb(NO3)2, however, caused significant increase in LP and GSH and significant reduction (p<0.05) in brain weight but has no effect on the antioxidants. PB induced significant reduction in brain weight and significant increase (p<0.05) in LP, CAT and GSH whereas it has no effect on SOD. PU induced significant reduction in brain weight and CAT while it induced significant increase (p<0.05) in LP but has no effect on GSH level. In conclusion, these results have demonstrated that PU and all the chemical neurotoxins investigated in this study except DDT induced brain damage (measured as reduced brain weight) in some cases as a consequence of oxidative stress and in some other cases by some unknown mechanisms. PU like some of these chemical neurotoxins induced brain damage presumably as a consequence of oxidative stress.
PDF Abstract XML References Citation
How to cite this article
O.L. Adebayo, B. O. Adegbesan and G. A. Adenuga, 2009. Comparison of the Effects of Low Protein Diet Versus Chemical Neurotoxins on Brain Weight, Brain Lipid Peroxidation and Antioxidant Status of Rats. Asian Journal of Biological Sciences, 2: 7-13.
URL: https://scialert.net/abstract/?doi=ajbs.2009.7.13
URL: https://scialert.net/abstract/?doi=ajbs.2009.7.13
INTRODUCTION
Neurotoxicants are chemical substances that adversely act on the structure or function of the nervous system and have serious impact on human health. They affect the nervous system in different ways; this depends on the molecular structure of the neurotoxic chemical substance and the specific function and structure of the nervous system tissue (Rodier, 2004). Neurotoxins that act on the Central Nervous System (CNS) impair neurotransmission in the brain and the spinal cord causing confusion, irritability, fatigue and other behavioral changes while neurotoxins that act on the Peripheral Nervous System (PNS) impair neurotransmission in the other biological systems causing weakness, prickling or tingling in the limbs and loss of motor control (Campbell, 1993).
Protein Undernutrition (PU), a wasting malnutrition and simple nutrient deficiency syndrome, is a problem that concerns about half the world’s children (Orbak et al., 1998). PU has been established to affect the brain and impairs its functions. Reports have it that malnutrition imposed during early life not only reduces growth of the brain but also leaves it permanently smaller in size (Levitsky and Strupp, 1995; Adebayo and Adenuga, 2007).
Despite numerous reports on the fact that PU impairs brain function and so is suspected to be neurotoxic, it is not known whether PU affects the brain by the same mechanism as those of well known chemical neurotoxins such as potassium cyanide, lead, phenobarbital and DDT.
Lead (Pb) is recognized as a serious threat to human health because it affects both the CNS and PNS . Its effects include acute encephalopathy and peripheral motor neuropathy (Donovick and Burright, 1992; El-Sokkary et al., 2003). Pb has direct neurotoxicological effects on the CNS; such as decreased synapse growth and density, impaired neuron growth, impaired neuronal differentiation, induced glial cell development and altered neurochemical signals (Minnema, 1992).
Cyanide, another well-known neurotoxicant, affects the corporal striata, hippocampus, cortical gray matter and substantia nigra part of the brain (Odunuga et al., 1998). Its neurotoxic effect was also observed in the structure and function of neuronal cells (Ardelt et al., 1994).
Equally serious is 1,1,1- trichloro-2, 2 bis -(ρ-chlorophenyl) ethane (DDT), which was worldwide used as pesticide. It has been reported that neonatal exposure to DDT clearly affected the muscarinic cholinergic receptor of the cerebral cortex in new born mice, leading to permanent disturbances of the cholinergic system in the brain combined with disruption of learning capacity in adult life (Dorner and Plagemann, 2002).
Phenobarbital (PB), however, is a barbituric acid derivative that acts as a nonselective CNS system depressant. It promotes binding to inhibitory Gamma-Aminobutyric acid subtype receptors and modulates chloride currents through receptor channels. It also inhibits glutamate induced depolarization (Shaw and Finkle, 1994).
The objective of this study was to determine whether PU acts by the same or similar mechanism as those of chemical neurotoxins in affecting the brain.
MATERIALS AND METHODS
Animals and Chemicals
Fifty male weanling albino rats obtained from the animal house of the University of Agriculture, Abeokuta, Nigeria, were used for the experiment in 2007. The animals were housed and acclimatized for 2 weeks in wire cages before the commencement of the experiment. Casein and all reagents used were of analytical grade and obtained from Sigma Chemical Co., USA and BDH chemicals Ltd., Poole, England.
Experimental Protocol and Treatment
Animals were randomly assigned to ten groups, A, B, C, D, E, F, G, H, I and J with five rats per group. Animals in group A were placed on normal (16 % Casein) diet while group B were placed on low protein diet (5% Casein). Low Protein Diet (LPD) and Normal Protein Diet (NPD) were prepared according to Olowookere (1994). Animals in groups C-J were fed on Commercially Produced Diet (CPD). Animals in group C were given CPD (in powered form) mixed with potassium cyanide (KCN, 200 ppm) (Odunuga and Adenuga, 1997) while group D received no treatment and thus served as control for KCN-treated. Group E animals were given 80 mg (kg b.wt.)-1 PB (dissolved in 0.1N NaOH and pH adjusted to 9.0 with HCl) intraperitoneally for 4 days (Adenuga et al., 1992) while group F animals (control rats) were given at the same time appropriate vehicle (i.e., 0.1N NaOH, pH 9.0). Group G animals were also given a single dose of 75 mg (kg b.wt.)-1 DDT dissolved in oil and group H (control rats) received the same appropriate vehicle (Olive oil) (Odunuga and Adenuga, 1997). Group I animals were given a single dose of 100 μ mole (kg b.wt.)-1 Lead nitrate (Pb(NO3)2) intravenously, dissolved in 0.9% (w/v) normal saline (Adegbesan and Adenuga, 2007) while the last group, i.e., group J (given 0.9% saline only) served as the control.
Groups A and B were allowed to feed on their respective diets for 14 weeks in order to ensure the induction of protein undernutrition before they were sacrificed. Groups’ C animals (after KCN- treatment) and D were fasted overnight and allowed their diets and water for 48 h before the sacrificed. Group E (PB- treated) animals were sacrificed on the fifth day after treatment and group G (DDT- treated) was sacrificed on the eighth day with their controls respectively (i.e., Groups F and H). For groups I and J, the animals were sacrificed 72 h after group I treatment. Groups E-J were allowed their diets and water till they were sacrificed. At the end of each treatment period, the animals were sacrificed and brains were quickly removed and kept in the freezer at 4°C.
Preparation of Brain Homogenate
Two sets of brain homogenates were prepared. 0.1 g of each part of the brain of individual sample was homogenized in 1 mL 0.01 M phosphate buffer, pH 7.4, kept frozen in the freezer and used for Catalase, Glutathione and Lipid Peroxidation analysis while another 0.1 g of each of the brain sample was homogenized in 1 mL of isolation buffer (250 mM Sucrose, 5 mM Tris, 1 mM Mercaptoethanol and 0.5 mM Phenylsulphonyl-flouride (PMSF) pH 7.4) also kept in the freezer and used for superoxide dismutase analysis.
Biochemical Analyses
Lipid Peroxidation Assay
Brain Lipid Peroxidation was carried out by measuring the thiobarbituric acid-reactive (TBAR) products using the procedure of Vashney and Kale (1990). The method is based on formation of pink colored product when Malondialdehyde treated with 2- thiobarbituric acid which has a maximum absorbance at 531.87 nm.
Glutathione Assay
Assay for glutathione (GSH) was done by the method of Beutler et al. (1963). This method is based on the development of a stable yellow color when 2-nitrobenzoic acids is added to sulfhydryl compounds.
Catalase Assay
The catalase activity of each brain samples was determined by the method of Sinha (1972) but with a slight modification. About 0.1 mL of each brain part was mixed with 4.9 mL of distilled water. One milliliter of the mixture was added to H2O2 -phosphate buffer mixture. The principle is based on the formation of chromic acetate when hydrogen peroxide (H2O2) reacts with dichromate-glacial acetic mixture at 100°C. The decomposition of H2O2 when acted upon by catalase and reduction in the green coloration is measured spectrophotometrically at 570 nm.
Superoxide Dismutase (SOD) Assay
SOD activity was determined by the method of Del-Maestro et al. (1983). This assay is based on the ability of SOD to scavenge superoxide anion radical (O2), which, by shortening reaction chains, decreases the overall rate of pyrogallol autoxidation.
Statistical Analysis
The statistical analysis was carried out using independent t-tests (SPSS software).
RESULTS
The results of the present experiment, as presented in Table 1 shows that Potassium cyanide induced significant reduction (p<0.05) in brain weight and SOD activity and significant increase (p<0.05) in catalase activity but no change were observed in lipid peroxidation and GSH level. DDT, however, only induced significant reduction (p<0.05) in catalase activity while it does not affect brain weight, lipid peroxidation, SOD activity and GSH level (Table 2).
| Table 1: | Effect of KCN on rat brain lipid peroxidation, brain weight and antioxidants status |
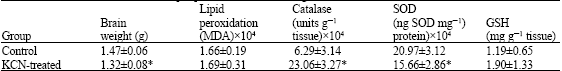 | |
| Treated rats were given diet containing 200 ppm KCN. Both treated and control groups were fasted overnight and allowed their diets and water for 48 h before sacrificed. Value are presented as Means±S.D, *p<0.05 | |
| Table 2: | Effect of DDT on rat brain lipid peroxidation, brain weight and antioxidants status |
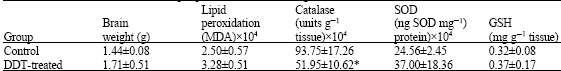 | |
| Treated rats were given a single dose of 75 mg kg-1 b. wt. of DDT and sacrificed on the eighth day. Value are presented as Means±S.D, *p<0.05 | |
| Table 3: | Effect of Pb(NO3)2 on rat brain lipid peroxidation, brain weight and antioxidants status |
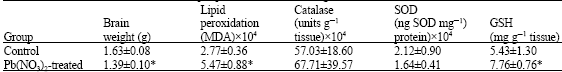 | |
| Rats treated with Pb(NO3)2 were given a single dose of 100 μ mole kg-1 b. wt. and allowed their diets and water for 48 h before sacrificed. Value are presented as Means±S.D, *p<0.05 | |
| Table 4: | Effect of PB on rat brain lipid peroxidation, brain weight and antioxidants status |
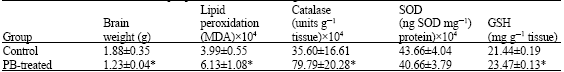 | |
| Rats treated with PB were given a single dose of 80 mg kg-1 b. wt. and allowed their diets and water. Rats were sacrificed five days later. Value are presented as Means±S.D, *p<0.05 | |
| Table 5: | Effect of PU on rat brain lipid peroxidation, brain weight and antioxidants status |
 | |
| Control rats were given diet containing 16% casein and PU treated rats were given diet containing 5% casein for 14 weeks. Value are presented as Means±S.D, *p<0.05, **p<0.001 | |
Lead nitrate caused significant increased (p<0.05) in both lipid peroxidation and GSH level but also induced significant reduction (p<0.05) in brain weight whereas it has no significant effect on both catalase and SOD activities as presented in Table 3. The result of Phenobarbital showed that it induced significant reduction (p<0.05) in brain weight and significant increase (p<0.05) in lipid peroxidatiion, catalase activity and GSH level whereas it has no effect on SOD activity as shown in Table 4. Finally, Protein undernutrition induced significant reduction (p<0.05) in brain weight, catalase activity while it induced significant increase (p<0.05) in lipid peroxidation whereas it has no effect on GSH level (Table 5).
DISCUSSION
Reduced brain weight has been associated with the effects of many neurotoxins (Minnema, 1992; El-Sokkary et al., 2003). The results of these experiments revealed that all investigated neurotoxins excepts DDT induced a reduction in brain weight. It seems that the dosage of DDT used in this study originally used for studies in the liver as reported by Adenuga et al. (1992) is not strong enough to induce a reduction in brain weight. Protein undernutrition, like the chemical neurotoxins attenuated the brain weight of rats, suggesting that one of the consequences of PU is brain damage.
In order to understand the mechanism of the neurotoxic effect of PU and that the severity of PU-induced brain damage may be comparable to those of the aforesaid chemical neurotoxins, an investigation on its effect on brain lipid peroxidation was conducted and compared to those of chemical neurotoxins. Generally, all the chemical neurotoxins used in this investigation except KCN and DDT induced an increase in brain lipid peroxidation, suggesting that the mechanism of neurotoxic effects of KCN and DDT are different from those of the other neurotoxins investigated. It is well known that KCN inhibits the terminal reaction of the oxidase reaction of the respiratory chain (Lonesome and Eaton, 1990). While lead acetate and Phenobarbital have been associated with oxidative stress (Demir et al., 2001; Adegbesan and Adenuga, 2007), protein undernutrition also elicited an increased lipid peroxidation showing that the mechanism of action has to do with oxidative stress.
This study also show that the effects of the chemical neurotoxin on the antioxidant status of the brain varied widely. For example, while the level of catalase was increased in KCN and phenobarbital (PB) treated rats, it was reduced in DDT treated and PU rats whereas lead acetate has no significant effect on catalase. While the increase in catalase in KCN and PB could be attributed to increase in the expression of catalase and also as the consequence of an adaptive response to oxidative stress induced by these neurotoxins (Diez-Fernandez et al., 1998), the reduction in catalase activities in both DDT and PU treated rats suggest a repression of catalase synthesis and a different mechanism for the protection of brain against reactive oxygen species generated by these neurotoxins. Other antioxidant enzymes, for example, glutathione peroxidase (GPx) might be involved in the scavenging of these free radicals as GPx is known to have higher affinity for hydrogen peroxide (H2O2) than catalase (Czarniewska et al., 2003). GPx uses GSH as a hydrogen donor to eliminate H2O2 and to convert the organic hydroperoxide into alcohol. Present results showed that GSH was not significantly affected suggesting that there might be a mechanism for replenishing GSH. The mechanism for this could be that oxidized glutathione (GSSG) which is the product of GPx reaction is reduced by Glutathione Reductase (GR) in the presence of NADPH (reduced nicotinamide adenine dinucleotide phosphate) and in this way the GSH level is restored in the cell (Czarniewska et al., 2003). Regrettably, the levels of these enzymes were not determined in this study and so is an area for further studies.
KCN decreased SOD activity but other neurotoxins investigated have no effect on SOD activity. The decreased in SOD activity by KCN suggests also that SOD was not actively involved in the role of protecting the brain against the reactive oxygen species. The non effect of the other neurotoxins on the activity of SOD may be due to the dosage used for the study. For example, it has been reported that the activity of SOD increases with increase in dosage (Barros et al., 1994). It is unfortunate however, that the effect of PU on SOD was not determined. A significant increase (p<0.05) in total GSH was observed in lead acetate and PB treated rats while no change was observed in KCN, DDT and PU rats. The increase could be attributed almost exclusively to an increase in bound glutathione. For example, PB induces glutathione-binding proteins and increased the level of bound glutathione without affecting unbound glutathione. Thus, the concentration of the latter probably regulates glutathione synthesis (Kaplowitz et al., 1980).
In conclusion, the se results have demonstrated that PU and all the chemical neurotoxins investigated in this study except DDT induced brain damage (measured as reduced brain weight) in some cases as a consequence of oxidative stress and in some other cases by some unknown mechanisms. These results have demonstrated that PU like some chemical neurotoxins investigated in this study induced brain damage, presumably as a consequence of oxidative stress. It has been reported earlier that selenium alone without protein replenishment abolished PU induced brain damage in rats (Adebayo and Adenuga, 2007). The type(s) of prooxidants involved in PU induced brain damage remains to be established.
ACKNOWLEDGMENTS
We expressed our profound gratitude to Messrs Osifeso and Osineye for their technical assistance and also to Dr E.O Okegbile for his assistance in the procurement of Chemicals.
REFERENCES
- Adebayo, O.L. and G.A. Adenuga, 2007. Protective effect of selenium on protein-undernutrition-induced brain damage in rats. Biol. Trace Elem. Res., 116: 227-234.
Direct Link - Adegbesan, B.O. and G.A. Adenuga, 2007. Effect of lead exposure on liver lipid peroxidative and antioxidant defense systems of protein-undernourished rats. Biol. Trace Elem. Res., 116: 219-225.
Direct Link - Adenuga, G.A., E.A. Bababunmi and R.G. Hendrickse, 1992. Depression of Ca2+-ATPase activity of the rat liver endoplasmic reticulum by the liver tumour promoters 1,1,1-trichloro-2,2-bis(ρ-chlorophenyl)-ethane and Phenobarbital. Toxicology, 71: 1-6.
PubMedDirect Link - Ardelt, B.K., J.L. Borowitz, E.U. Maduh, S.L. Swain and G.E. Isom, 1994. Cyanide-induced lipid peroxidation in different organs: subcellular distribution and hydroperoxide generation in neuronal cells. Toxicology, 89: 127-137.
PubMedDirect Link - Barros, S.B., R. Pimente, K. Simizu, L.A. Azzalis, I.S. Costa and V.B. Junqueira, 1994. Dose-dependent study of liver lipid peroxidation related parameters in rats treated with pp’-DDT. Toxicol. Lett., 70: 33-38.
PubMedDirect Link - Beutler, E., O. Duron and B.M. Kelly, 1963. Improved method for the determination of blood glutathione. J. Lab. Clin. Med., 61: 882-888.
PubMedDirect Link - Czarniewska, E., A. Kasprzyk and K. Ziemnicki, 2003. Effect of paraquat and metoxychlor on antioxidant enzymes in frog Rana esculenta L. liver. Biol. Lett., 40: 125-133.
Direct Link - Del-Maestro, R.F., W. McDonald and R. Anderson, 1983. Superoxide Dismutase, Catalase and Glutathione Peroxidase in Experimental and Human Brain Tumour in Oxyradicals and Their Scavenger Systems. In: Oxidative Enzymes in Tissue Homogenate, Greenwald, R. and Cohen, G. (Eds.). Vol. 2, Elsevier, New York, USA., pp: 291-296.
- Demir, O., E. Yazar, V. Altunok, M. Elmas and V. Ozdemir, 2001. Effect of Phenobarbital on antoxidant enzyme activities and blood gas parameters in Balb/C mice. Rev. Med. Vet., 152: 723-726.
Direct Link - Diez-Fernandez, C., N. Sanz, A.M. Alvarez, A. Wolf and M. Cascales, 1998. The effect of non-genotoxic carcinogens, Phenobarbital and clofibrate, on the relationship between reactive oxygen species, antioxidant enzyme expression and apoptosis. Cacinogenesis, 19: 1715-1722.
PubMedDirect Link - Dorner, G. and A. Plagemann, 2002. DDT in human milk and mental capacities in children at schoolage: An additional view in PISA 2000. Neuroendocrinol. Lett., 23: 427-431.
Direct Link - El-Sokkary, G.H., E.S. Kamel and R.J. Reiter, 2003. Prophylactic effect of melatonin in reducing lead-induced neurotoxicity in the rat. Cell. Mol. Lett., 8: 461-470.
Direct Link - Kaplowitz, N., J. Kuhlenkamp, L. Goldstein and J. Reeve, 1980. Effect of salicylates and phenobarbital on hepartic glutathione in the rat. J. Pharmacol. Exp. Ther., 212: 240-245.
PubMedDirect Link - Levitsky, D.A. and B.J. Strupp, 1995. Malnutrition and the brain: Changing concepts, changing concerns. J. Nutr., 125: 2212S-2220S.
Direct Link - Odunuga, O.O. and G.A. Adenuga, 1997. Sodium Nitrite alone protects the brain microsomal Ca2+-ATPase against potassium cyanide-induced neurotoxicity in rats. Biosci. Rep., 17: 543-546.
PubMedDirect Link - Orbak, Z., Y. Akin, E. Varoglu and H. Tan, 1998. Serum thyroid hormone and thyroid gland weight measurements in protein-energy malnutrition. J. Pediatr. Endocrinol. Metab., 11: 719-724.
PubMedDirect Link - Rodier, P.M., 2004. Environmental causes of central nervous system maldevelopment. Pediatrics, 113: 1076-1083.
PubMedDirect Link - Sinha, A.K., 1972. Colorimetric assay of catalase. Anal. Biochem., 47: 389-394.
CrossRefPubMedDirect Link - Varshney, R. and R.K. Kale, 1990. Effects of calmodulin antagonists on radiation-induced lipid peroxidation in microsomes. Int. J. Radiat. Biol., 58: 733-743.
CrossRefDirect Link