
Li-Juan Xiong
Triticeae Research Institute, Sichuan Agricultural University, Yaan, Sichuan 625014, China
Wei Li
Triticeae Research Institute, Sichuan Agricultural University, Yaan, Sichuan 625014, China
Yu-Ming Wei
Triticeae Research Institute, Sichuan Agricultural University, Yaan, Sichuan 625014, China
You-Liang Zheng
Triticeae Research Institute, Sichuan Agricultural University, Yaan, Sichuan 625014, China
Asian Journal of Biological Sciences
Year: 2008 | Volume: 1 | Issue: 1 | Page No.: 19-25







ABSTRACT
In order to exploit new genetic resources and provide fundamental materials for the breeding improvement of bread wheat quality, genetic variation of high-molecular-weight glutenin subunits and gliadin in 29 macha wheat accessions were observed by acidic polyacrylamide-gel electrophoresis and sodium dodecyl sulphate polyacrylamide-gel electrophoresis. Nine HMW-glutenin subunits alleles and 9 combinations were identified. Subunit null (82.8%), 7+8 (53.3%) and 2+12 (82.8%) scored the highest frequency at Glu-A1, Glu-B1 and Glu-D1 loci, respectively. In addition, subunits 7+9 (23.3%) was also found at higher frequency. A total of 49 gliadin bands and 28 patterns were detected and the polymorph among all accessions was identified in most of bands (97.96%). Furthermore, all materials could be clustered into three major groups based on genetic similarity coefficient. The results indicated that the variations of gliadin among macha accessions were not associated with their geographic origins.
PDF Abstract XML References Citation
How to cite this article
Li-Juan Xiong, Wei Li, Yu-Ming Wei and You-Liang Zheng, 2008. Variation of High-Molecular-Weight Glutenin Subunits and Gliadin in T.aestivum ssp. macha. Asian Journal of Biological Sciences, 1: 19-25.
URL: https://scialert.net/abstract/?doi=ajbs.2008.19.25
URL: https://scialert.net/abstract/?doi=ajbs.2008.19.25
INTRODUCTION
In the endosperm of wheat (Triticum aestivum L.), the main storage protein classes are glutenin and gliadin. The wheat baking quality is controlled by the content and composition of wheat endosperm proteins. Specific HMW-glutenin subunits have been closely associated with bread making quality (Payne, 1987; He et al., 1999; Barro et al., 2003; Edwards et al., 2007; Lefebvre and Mahmoudi, 2007). Glutenin and gliadin are generally considered to contribute to the viscosity and extensibility of gluten. Glutenins were composed of high-molecular-weight (HMW) subunits and low-molecular-weight (LMW) subunits. HMW-glutenin subunits are controlled by three gene loci, located on the long arm of chromosomes 1A, 1B and 1D and identified as Glu-A1, Glu-B1 and Glu-D1 (Payne, 1987). Gliadins on the other hand are encoded by the complex gene family, which located on the short arms of group 1 and 6 chromosomes (Lafiandra et al., 1984; Hassani et al., 2007).
In order to improve wheat quality breeding, it is essential to exploit the valuable gene resource from wheat relative species. One of the oldest hexaploid cultivated species, T. aestivum ssp. macha (2n = 6x = 42, AABBDD), that was grown mainly in the west of Gelugia in the past. It possessed stronger tillering ability, resist smut and moisture tolerance. Because it has the same genome structure as the common wheat, the useful genes of T. aestivum ssp. macha could be easy transferred into common wheat. Datiashvili (1983) found that there was better ratio between gliadin and glutenin in T. aestivum ssp. macha. Fang et al. (1997) suggested that moisture tolerance of macha wheat was controlled by a pair of dominant gene and further scored a SSR marker Xgwm148 (Fang and Cai, 2003). Zhang et al. (2003) further demonstrated that these genes are located on 5A chromosome based on RAPD marker. Xiong et al. (2006) also reported that macha wheat had higher plant height, strong tillering ability and more spikelets. However, little information for macha was investigated. This confined the utility of T. aestivum ssp. macha in wheat improvement breeding. In the present study, were report on genetic diversity of HMW-glutenin subunits and gliadin as evaluated in 29 T. aestivum ssp. macha accessions from 11 countries.
MATERIALS AND METHODS
Plant Materials: A total of 29 accessions from different countries were kindly provided by Dr. Harold Bockelaman, USDA-ARS, National Small Grains Collection (Table 1). This study was conducted in 2007. Electrophoresis was performed at the Triticeae Research Institute, Sichuan Agriculture University.
A-PAGE: According the standard protocol of ISTA acidic polyacrylamide-gel electrophoresis (pH 3.1) (A-PAGE) (Draper, 1987), the gliadin electrophoresis was carried out. Common wheat cultivar Chinese Spring was used as reference.
SDS-PAGE: According to the procedures of Ng and Bushuk (1987), the HMW-glutenin subunits were separated by sodium dodecyl sulphate polyacrylamide-gel electrophoresis (SDS-PAGE). HMW-glutenin subunits were identified according to Payne and Lawrence (1983). Common wheat cultivar Chinese Spring (null/7+8/2+12) and Chuanyu12 (1/7+8/5+10) were used as reference.
Statistic Analysis: The presence or absence of gliadin bands was carefully recorded. The present bands were recorded as 1 and the bands absent as 0. Based on the gliadin bands data, the Genetic Similarity (GS) was calculated according to Nei`s (1973) method as GS = 2Nij/(Ni+ Nj), where Ni is the number of the band in the material i; Nj is the number of the gliadin band in the material j; Nij is the number of the gliadin band shared by material i and j.
| Table 1: | A list of accessions and their country of origin |
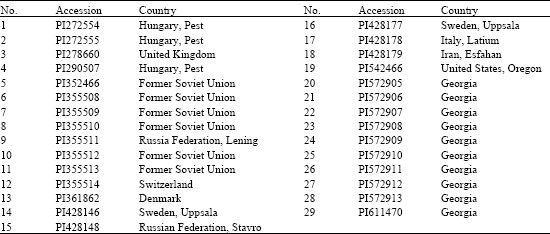 | |
Using the unweighted pair-group method, a clustering analysis was carried out based on GS index under NTSYS-pc (version 2.1) software.
RESULTS
Gliadin Variations: A total of 49 gliadin polymorphic bands and 28 patterns were detected in 29 T. aestivum ssp. macha accessions. The number of bands ranged from 19 to 34 per accession, with the mean of 25. Ten bands were detected in α-, β- and γ-region, respectively and 19 bands in ω-region. Twenty-two, nineteen, twenty-two and twenty-six gliadin patterns were identified in α-, β-, γ- and ω-region, respectively (Fig. 1). The genetic similarity (GS) among T. aestivum ssp. macha ranged from 0.29 to 0.95, with a mean of 0.61. Based on the gliadin GS coefficient, the genetic relationships among macha wheat accessions were further investigated by UPGMA program. The result indicated that all accessions could be divided into three groups at the 0.58 level of GS (Fig. 2). Group 1 contained 19 accessions, including all accessions from Hungary, Iran, Italy, United Kingdom and most of accessions from the Former Soviet Union and Georgia. It could be further divided into two subgroups at the 0.63 level of GS. Group 3 only contained one accessions (PI355513) from Former Soviet Union. Group 2 contained 9 accessions, including 5 accessions from Switzerland, United States, Denmark, Sweden and Russian Federation, respectively and 4 accessions from Georgia. Group 2 also could be further divided into two subgroups. One accession (PI572909) from Georgia formed subgroup 2, whereas other three accessions from Georgia were clustered in another subgroup. Therefore, the clustering of macha wheat is not associated with their geographic origin. These results also indicate that the macha wheat accessions from Former Soviet Union and Georgia had higher genetic variation than material from other sources.
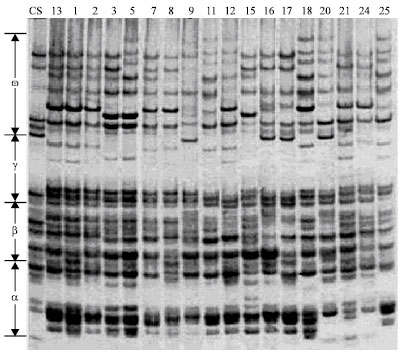 | |
| Fig. 1: | Gliadin patterns for 17 accessions of macha wheat in APAGE profile Note:The name of material was listed in Table 1, CS: Chinese spring, α, β, γ and ω refer to the corresponding gliadin region |
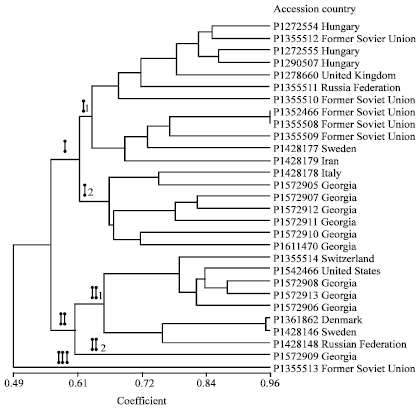 | |
| Fig. 2: | A dendrogram generated from gliadin |
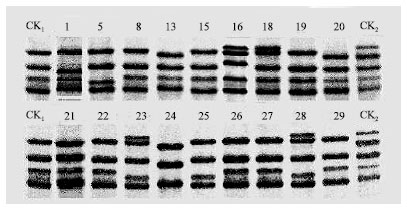 | |
| Fig. 3: | HMW-GS of macha wheat accessions after SDS-PAGE Note:The name of material was listed in Table 1, CK1: Chinese spring, CK2: Chuanyu12 |
HMW-GS Glutenin Variations: A total of 9 HMW-GS subunits were identified in all accessions, including 2, 5 and 2 subunits at Glu-A1, Glu-B1and Glu-D1 loci, respectively (Table 2) (Fig. 3). At the Glu-A1 locus, subunit null had the highest frequency (82.8%), whereas subunit 1 had low frequency (17.2%). At Glu-B1 locus, subunits 7+8 was the predominant frequency 55.2%, followed by subunit 7+9 (24.1%) and the subunits 8, 20 and 6+8 with low frequency were also detected. At Glu-D1 locus, only subunits 2+12 and 5+10 were identified and the former subunit had the highest frequency (82.8%). Nine HMW-GS patterns were identified among all accessions. The distribution of HMW-glutenin patterns are listed in Table 3.
| Table 2: | The distribution frequency of HMW-GS alleles in macha wheat accessions |
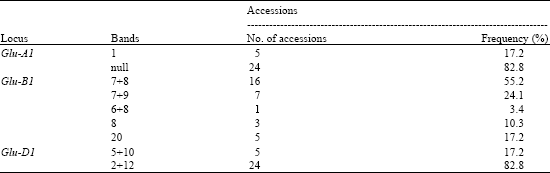 | |
| Table 3: | The combination and distribution frequency of HMW-GS in macha wheat accessions |
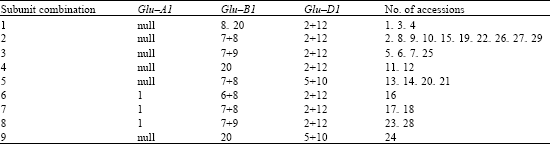 | |
The pattern null/7+8/2+12 was detected in 10 accessions and had the highest frequency. Four patterns, including null/7+9/2+12, null/7+8/5+10, 1/7+8/2+12 and 1/7+9/2+12, had higher frequency. The HMW-GS pattern 1/6+8/2+12 and null/20/2+12 were only observed at low frequency.
DISCUSSION
The gliadin patterns with a great diversity of variations have been used as fingerprints, genotype identification, quality markers and so on (Yan et al., 1992; Liu et al., 1999; Lang et al., 2001). Zhang et al. (1995) suggested that A-PAGE could provide an available method for identifying wheat germplasm. In this study, the higher genetic diversity of gliadin was found, a total of 28 gliadin patterns were identified in 29 T. aestivum ssp. macha. This indicates that gliadin could provide a valuable and exact diversity marker for T. aestivum ssp. macha accessions. These data is supported by the results of Khachidze (1989) who also observed a great diversity of variations in T. aestivum ssp. macha and found some bands in the patterns that is species specific. Yakobashvili and Naskidashvili (1988) reported that the greatest polymorphism (11 variants of gliadin blocks) was found for the gliadin locus on chromosome 6A. One of its variants was identical to block Gld 6A20 of bread wheat. Six variants of blocks associated with chromosome 1B, 3 were identical to blocks in bread wheat (Gld-1B4, Gld-1B8 and Gld-1B10). Metakovsky and Iakobashvili (1990) further reported an additional component mapped between Glu-B1 and Gli1B in the gliadin spectrum of T. aestivum ssp. macha. The analyses of seed storage protein components of the gliadin and glutenin in T. aestivum ssp. compactum, T. aestivum ssp. sphaerococcum, T. aestivum ssp. macha, T. aestivum ssp. spelta had revealed limited variation at the tightly linked coding loci Gli-D1/Glu-D3 and Glu-D1, located respectively on the short and long arm of chromosome 1D and at the Gli-D2 locus, positioned on the short arm of chromosome 6D (Lafiandra et al., 1992).
In this study, a great diversity of variations on gliadin loci in T. aestivum ssp. macha accessions, while the relationship between genetic variety and other properties traits need further study. Meanwhile, two accessions with the same gliadin bands had no diversity in all properties, but had further close relative relations, such as PI352466 and PI355508 used in this study. So the genetic varieties between accessions need other methods, like by molecular marker technology to detect and analyses.
In the tree of the cluster analysis, accessions from Georgia were clustered together, but only few accessions were grouped with other counties accessions. From the clustering dendrogram, the Georgia accessions could be divided into two groups. Accessions PI572908, PI572913, PI572906 and PI572909 formed a group, while others were another group. This indicates that there were two allelic variety types on gliadin locus in Georgia accessions. In this study, the clustering analysis indicated that the gliadin variations among T. aestivum ssp. macha are not associated with their geographic origin. High level of HMW-GS polymorphism was observed in the 29 T. aestivum ssp. macha by SDS-PAGE method. The main HMW-glutenin patterns was null (Glu-A1), In all ten Georgia accessions, the pattern (null, 7+8 and 2+12) had the highest frequency, the pattern (null, 7+9, 2+12) had the similar performance in Former Soviet Union accessions. The consistent efforts have been made to improve wheat quality by accumulating good HMW-glutenin subunits, such as 1, 5+10. Yakobashvili`s et al. (1988) indicated that the species included T. aestivum ssp. macha, T. aestivum ssp. spelta and T. aestivum ssp. aestivum were closely related through comparison with HMW glutenins of 3 species by SDS-PAGE. The greatest polymorphism in T. aestivum ssp. macha and T. aestivum ssp. spelta was for Glu-1B, for which the greatest difference between them was also found suggesting that they may had different donors of genome B.
ACKNOWLEDGMENT
This research was supported by the National Natural Science Foundation of China (No. 30300219 and 30571163) and A Foundation for the Author of National Excellent Doctoral Dissertation of PR China (No. 200357 and 200458). Y.-L. Zheng was supported by the Program for Changjiang Scholars and Innovative Research Teams in University of China (IRT0453).
REFERENCES
- Barro, F., P. Barcelo, P.A. Lazzeri, P.R. Shewry, J. Ballesteros and N.A. Martin, 2003. Functional properties of flours from field grown transgenic wheat lines expressing the HMW glutenin subunit 1AX1 and 1DX5 genes. Mol. Breeding, 12: 223-229.
CrossRefDirect Link - Draper, S.R., 1987. ISTA and Variety committee: Report of the working group for biochemical tests for cultivar identification 1983-1986. Seed Sci. Tech., 15: 431-434.
Direct Link - Edwards, N.M., M.C. Gianibelli T.N. McCaig, J.M. Clarke, N.P. Ames, O.R. Larroque and J.E. Dexter, 2007. Relationships between dough strength, polymeric protein quantity and composition for diverse durum wheat genotypes. J. Cereal Sci., 45: 140-149.
CrossRef - Xianwen, F., C. Yang and C. Shibin, 1997. Genetic evaluation of waterlogging tolerance in Triticum macha. Jiangsu J. Agric. Sci., 13: 73-75.
Direct Link - Hassani, M.E., M.R. Shariflou, M.C. Gianibelli and P.J. Sharp, 2008. Characterisation of a ω-gliadin gene in Triticum tauschii. J. Cereal Sci., 47: 59-67.
CrossRef - He, G.Y., L. Rooke, S. Steele, F. Bekes and P. Gras et al., 1999. Transformation of pasta wheat (Triticum turgidum L. var. durum) with high-molecular-weight glutenin subunit genes and modification of dough functionality. Mol. Breeding, 5: 377-386.
CrossRef - Lafiandra, D., D.D. Kasarda and R. Morris, 1984. Chromosomal assignment of genes coding for the wheat gliadin protein components of the cultivars Cheyenne and Chinese Spring by two dimensional (two-pH) electrophoresis. Theor. Applied Genet., 68: 531-539.
CrossRef - Lafiandra, D., S. Masci, R. Dovidio, O.A. Tanzarella, E. Porceddu and B. Margiotta, 1992. Relationship between the D genome of hexaploid wheats (AABBDD) and Ae. Squarrosa as deduced by seed storage proteins and molecular marker analyses. Here. Landskrona, 116: 233-238.
CrossRef - Lefebvre, J. and N. Mahmoudi, 2007. The pattern of the linear viscoelastic behaviour of wheat flour dough as delineated from the effects of water content and high molecular weight glutenin subunits composition. J. Cereal Sci., 45: 49-58.
CrossRefDirect Link - Hua, L., W. Yusheng and Z. Hui, 1999. Preliminary construction and application of gliadin fingerprints database of Chinese wheat germplasm. Acta Agron. Sin., 25: 674-682.
Direct Link - Nei, M., 1973. Analysis of gene diversity in subdivided populations. Proc. Natl. Acad. Sci. USA., 70: 3321-3323.
PubMedDirect Link - Ng, P.K.W. and W. Bushuk, 1987. Glutenin of Marquis wheat as a reference for estimating molecular weights of glutenin subunits by sodium dodecyl sulfate-polyacrylamide gel electrophoresis. Cereal Chem., 64: 324-327.
Direct Link - Payne, P.I., 1987. Genetics of wheat storage proteins and the effect of allelic variation on breadmaking quality. Ann. Rev. Plant Physiol., 38: 141-153.
Direct Link - Zhang, X., X. Yang and Y.S. Dong, 1995. Genetic analysis of wheat germplasm by acid polyacrylamide gel electrophoresis of gliadins. Sci. Agric. Sin. A, 28: 25-32.
Direct Link - Zhang, J.L., S.B. Cai, G.X. Zhang, J.B. Wei, C.Q. Zhang and Z.Q. Ma, 2003. Genetic mapping of genes conferring waterlogging-tolerance in Triticum macha using RAPD markers. J. Nanjing Agric. Uni., 26: 7-10.
Direct Link