
C.C. Onunogbo
Department of Biochemistry, National Root Crops Research Institute, Umudike, Umuahia, Abia State, Nigeria
O.C. Ohaeri
Department of Biochemistry, Michael Okpara University of Agriculture, Umudike, Umuahia, Abia State, Nigeria
C.O. Eleazu
Department of Biochemistry, National Root Crops Research Institute, Umudike, Umuahia, Abia State, Nigeria
LiveDNA: 234.3349
American Journal of Biochemistry and Molecular Biology
Year: 2013 | Volume: 3 | Issue: 1 | Page No.: 143-150







ABSTRACT
The effect of aqueous extract of mistletoe on the blood glucose, electrolytes, transaminases and phosphatase activities of alloxan induced diabetic rats was investigated using their respective kits. Twenty male rats, weighing between 90 and 120 g kg-1 b.wt. were used. Alloxan was injected into the experimental animals at a dosage of 100 mg kg-1 b.wt. The rats were divided into 5 groups: Group 1: non-diabetic received normal rat feeds comprising of clean water and growers mash. Group 2: diabetic control received the same treatment as group 1. Group 3: diabetic received 20% of the extract, Group 4: diabetic, received 40% of the extract while group 5 diabetic rats received 60% of the extract. The duration of the experiment was four weeks. Results indicate that all concentrations of the extract administered reduced the elevated blood glucose levels of the diabetic animals although they could not statistically ameliorate it (p>0.05), administration of 20 and 40% of the extract was most effective in ameliorating the altered K+ concentration of the diabetic rats, while 40 and 60% of the extract proved to be statistically most effective in ameliorating the elevated bicarbonate levels of the diabetic rats (p<0.05). All concentrations administered also ameliorated the altered sodium and alanine transaminase activities of the diabetic animals while they slightly improved their chloride levels (p>0.05). In addition, 60% of the extract significantly ameliorated the altered phosphatase activities and body weights of the diabetic rats while 40 and 60% of the extract significantly ameliorated the altered aspartate transaminase activities of the diabetic animals (p<0.05). It can be deduced that mistletoe has anti-diabetic potentials, in addition to ameliorating the altered electrolyte balance arising from diabetes. Finally, the dietary supplement may protect liver cells from free radical damage and could be effective in treating hepatoctye damage in diabetics.
PDF Abstract XML References Citation
Received: March 31, 2012;
Accepted: April 28, 2012;
Published: October 04, 2012
How to cite this article
C.C. Onunogbo, O.C. Ohaeri and C.O. Eleazu, 2013. Effect of Mistletoe (Viscum album) Extract on the Blood Glucose, Liver Enzymes and Electrolyte Balance in Alloxan Induced Diabetic Rats. American Journal of Biochemistry and Molecular Biology, 3: 143-150.
DOI: 10.3923/ajbmb.2013.143.150
URL: https://scialert.net/abstract/?doi=ajbmb.2013.143.150
DOI: 10.3923/ajbmb.2013.143.150
URL: https://scialert.net/abstract/?doi=ajbmb.2013.143.150
Key words: Liver enzymes, electrolytes, alloxan, diabetes
INTRODUCTION
Diabetes mellitus, a chronic disease with complex etiologies (Robert et al., 2002; Champe and Harvey, 1997) is a disease that has created serious cause for concern all over the world. The World Health Organization (WHO, 2002) reported that the number of diabetic patients is expected to increase from 150 million (King et al., 1998) in the year 2000 to 300 million or more by the year 2025. This has necessitated the screening of plants for anti-diabetic potentials. The search for the cure for diabetes mellitus continues along traditional and alternative medicine because many herbal supplements that have been used for the control of the disease are either not easily reachable (Wild et al., 2004) or they lack scientific evidence to support their effectiveness (Morelli and Zoorob, 2000). The use of synthetic agents such as sulphonylureas, on the other hand, has its limitations as it has been associated with side effects such as weight gain, hypoglycaemia (Aldhahi and Hamdy, 2003), cost effectiveness as well as correcting the fundamental biochemical lesion and diabetic complications.
Mistletoe (Viscum album) is a plant that is indigenous to the South-Eastern part of Nigeria where its used by the traditional medicine practitioners for the management of many metabolic diseases such as hypertension and diabetes mellitus (Obatomi et al., 1994). Mistletoe teas have been used for the prevention and management of stokes in parts of Nigeria and it is also believed to improve the circulatory system and heart function in traditional medicine (Deeni and Sadiq, 2002).
Although, at present, there are no reports of toxicity arising from the usage of this plant in diabetics, there is need to document such data. This forms the basis of this study aimed at investigating the effect of mistletoe extract on the liver enzymes and electrolyte balance in alloxan induced diabetic rats.
MATERIALS AND METHODS
Preparation of plant materials: The mistletoe plant used for the experiment was harvested in 2010 and identified by a Taxanomist in Michael Okpara University of Agriculture, Umudike, Nigeria. The plant was pulverized and dried in an oven at 60oC. It was then ground to flour and stored in an air tight container for further analysis.
Chemicals: Alloxan, glucose oxidase, aspartate transaminase, alanine transaminase and electrolyte reagent kits used were obtained from Sigma and Aldrich Chemical company, Uk. All other chemicals used for the animal experiments were purchased locally from Associated Laboratories in Aba, Abia State, Nigeria and were of analytical grade.
Animal experiment
Selection of animals and their care: Twenty male albino white rats (Wistar strain) weighing between 90-120 g used for this experiment were purchased from the University of Nigeria, Nsukka in 2010 and were kept in the animal house in the Federal University of Technology Owerri, Imo State, Nigeria. The animals were acclimatized for a period of 7 days to the laboratory conditions prior to the experiment in line with the University’s ethics of animal experiments. Rats were housed in well ventilated colony cages with 2 rats per cage at room temperature (27-30°C) with 12 h of light and dark cycle and had free access to drinking water and their diets (ad libithium). The rats were fed with commercial rat feed, obtained from Chukwuma Ventures Ltd, Owerri, Imo State, Nigeria.
Induction of diabetes: The rats were fasted for 24 h before injection of a freshly prepared solution of alloxan intraperitonially at a dosage of 100 mg kg-1 b.wt. This single dose of alloxan produced Type I diabetes having fasting blood sugar level of 204.37±1.24 mg dL-1 after five days of injection of alloxan.
Experimental procedure: The rats were divided into five groups with four animals in each group:
Group 1: Rats served as the normal (control) and they received oral administration of 60% aqueous extracts of the commercial rat feed and water for a period of four weeks at a dose of 100 mg kg-1 b.wt.
Alloxan was injected into the animals of group 2 at a single dose of 100 mg kg-1 b.wt. Those with fasting blood glucose above 180 mg dL-1 were categorized into groups 2-5:
Group 2: Diabetic control: The animals of this group received oral administration of 60% aqueous extract of the commercial rat feed and water for a period of four weeks at a dosage of 100 mg kg-1 b.wt.
Group 3: Diabetic rats with 20% concentration of mistletoe flour extract: The animals of this group received oral administration of 20% concentration of aqueous extract of mistletoe flour for a period of four weeks at a dosage of 100 mg kg-1 b.wt.
Group 4: Diabetic rats with 40% concentration of mistletoe flour extract: The rats of this group received oral administration of 40% concentration of aqueous extract of mistletoe flour for a period of four weeks at a dosage of 100 mg kg-1 b.wt.
Group 5: Diabetic rats with 60% concentration of mistletoe flour extracts: The animals of this group received oral administration of 60% concentration of aqueous extract of mistletoe flour for a period of four weeks. At the end of four weeks, the animals were starved overnight, stunned by blow and killed by decapitation and their blood was collected intraveneously from the heart using a 10 mL syringe. Their fasting blood glucose, transaminase, electrolytes and phosphatase activities were determined using their respective kits while the changes in their body weights were recorded twice in a week throughout the duration of the experiment.
Similarly, the initial and final body weights were measured with an electronic weighing balance. From these, Fasting Blood Glucose (FBG %) reduction and % weight change were calculated using the formula:
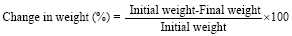 |
The percentage growth rate was calculated as:
The percentage change in fasting blood glucose was calculated as:
Determination of glucose: The serum glucose was determined using the glucose oxidase method as described by Cooper (1973). The principle was based on the fact that β-D-glucose is oxidized by glucose oxidase to produce D-glucoronic acid and hydrogen peroxide. The hydrogen peroxide is oxidatively coupled with 4-aminoantipyrine and phenol substitute, p-HBS, in the presence of peroxidase to yield a red quinone imine dye. The amount of colored complex formed is proportional to glucose concentration and can be read spectophotometrically at 500 nm against a reagent blank.
The concentration of glucose in the unknown samples was calculated from the equation:
Determination of aspartate transaminase activity (AST or SGOT): The method of Henry et al. (1960) was used in the analysis of the serum transaminase activity of the rats. This is measured by monitoring the concentration of oxaloacetate hydrazone formed with 2-4 dinitrophenyl hydrazine. The colored solution formed was measured spectrophotometrically at 546 nm against a reagent blank.
Determination of alanine transaminase (ALT or SGPT): Alanine transaminase was measured using the method of Henry et al. (1960). The principle is based on the reaction of α-ketoglutarate and L-alanine to form L-glutamate and pyruvate. The enzyme activity was measured by monitoring the concentration of pyruvate hydrazone formed with 2,-4 dinitrophenyl hydrazine using a UV spectrophotometer at 546 nm.
Determination of chlorides (Cl¯): The chlorides were analyzed using the method of (Tietz, 1976). The principle is based on the fact that chloride ions form a soluble non-ionized compound with mercuric ions and will displace thiocyanate ions from non-ionized mercuric thiocyanate. The released thiocyanate ions react with ferric ions to form a red color complex that absorbs light at 480 nm. The intensity of the color produced is directly proportional to the chloride concentration. The concentration of Cl¯ released was calculated from the equation:
Determination of sodium (Na+): The method of Henry (1974) was adopted for the assay. The principle is based on the fact that sodium is precipitated as the triple salt, sodium magnesium uranyl acetate with the excess uranium then being reacted with ferrocyanide, producing a chromophore whose absorbance varies inversely with the concentration of sodium in the test specimen. The absorbance of the released Na+ was read using a UV spectrophotometer at 550 nm and the concentration of Na+ in the sample was derived from the standard using the equation:
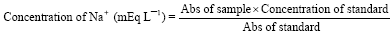 |
Determination of bicarbonates (HCO3¯): This was measured using the method of Henry (1974). The principle is based on the fact that phosphoenol pyruvate carboxylase catalyzes the reaction between phosphoenol pyruvate and carbon dioxide (bicarbonate) to form oxaloacetate and phosphate ion. Oxaloacetate is reduced to Malate with simultaneous oxidation of an equimolar amount of reduced nicotinamide adenine dinucleotide (NADH) to NAD+. This reaction is catalyzed by Malate dehydrogenase. This results in a decrease in absorbance at 340 nm that is directly proportional to HCO3¯ concentration in the sample. The concentration of HCO3¯ in the unknown sample was quantified from the equation:
Determination of potassium (K+): The serum K+ components of the samples was determined using the method of Henry (1974). This is determined by using the sodium tetraphenyl boron in a specifically prepared mixture to produce a colloidal suspension, the turbidity of which is proportional to the K+ concentration in the range of 2-7 mEq L-1 and which was read spectrophotometrically in a uv spectrophotometer at 500 nm against a reagent blank.
The concentration of the K+ was calculated thus:
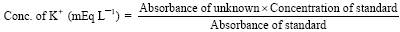 |
Determination of alkaline phosphatase (ALP): Alkaline phosphatase was measured using the method of Tietz (1976). The principle is based on the fact that ALP acts upon the AMP-buffered sodium Thymolphthalein monophosphate. The addition of an alkaline reagents stops enzyme activity and simultaneously develops a blue chromogen which is measured photometrically at 590nm against a reagent blank.
The concentration of ALP in the unknown sample (IU L-1) was calculated using the equation:
Statistical analysis: Data was subjected to analysis of variance using the Statistical Package for Social Science (SPSS) 15.0 windows version. Results are presented as Mean±SD. Means with differences were separated using the Duncan Multiple Range Test and results were considered significant at p<0.05.
RESULTS AND DISCUSSION
Analysis of the nutritive constituents of the commercial feeds that were administered to the rats indicated that it contained 16% protein, 5% fat, 7% crude fibre, 1% calcium, 0.45% phosphorous, 0.75% lysine, 0.36% methionine and 0.3% salt (Table 1).
Alloxan is known to destroy the β-cells of the islet of the Langerhans of the pancreas that function in the regulation of insulin secretion and thus leads to an increase in the blood concentration of glucose and type I diabetes mellitus (Eleazu et al., 2010). This accounts for the increase in the blood glucose of the diabetic animals. The fundamental mechanism underlying hyperglycemia in diabetes mellitus involves the over production of glucose (excessive hepatic glycogenolysis and gluconeogenesis) and or decreased utilization of glucose by the tissues (Latner, 1958; Dohi et al., 1998).
However, findings from the study reveal that all concentrations of the extract administered significantly ameliorated the hyperglycemia that resulted from the diabetes (p<0.05) as shown in Table 2.
Diabetes mellitus brings about an increase in the plasma glucose leading to cell dehydration and movement of K+ into the extra cellular fluid. This leads to an increase in the activity of parietal cells of the distal and cortical collecting tubules leading to increased renal excretion of K+ (Yared and Chiasson, 2003). In addition, glucosuria as observed in diabetes also leads to excretion of excess water, Na+ and K+ in urine (Yared and Chiasson, 2003). It is therefore possible that the electrolyte and water loss usually observed in diabetes would lead to depletion of the Extracellular Fluid (ECF) electrolytes and this could lead to the secretion of electrolytes by parietal and non-parietal cells.
| Table 1: | Nutrient composition of the feeds (growers feed)administered to the rats |
 | |
| Table 2: | Effect of mistletoe extract on serum chemistry of diabetic and non-diabetic rats |
 | |
Values in the same row with the same superscripts are not significantly different at p>0.05, n = 4 animals per group | |
| Table 3: | Effect of mistletoe extract on the enzyme activities of diabetic and non-diabetic rats |
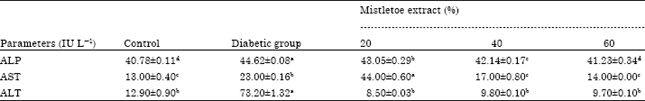 | |
Values in the same row with different superscripts are significantly different from each other at p<0.05, ALP: Alkaline phosphatase, AST: Aspartate transaminase, ALT: Alanine transaminase | |
This may therefore account for the significant reduction in the Na+ and K+ of the diabetic rats compared with the control as observed in Table 2.
Hyperglycemia as seen in uncontrolled diabetes is known to result in metabolic acidosis (Stoner, 2005) which causes an increase in respiratory activities and thus leads to an increase in the loss of CO2 from the blood. The loss of CO2 from the blood results in a reduction in bicarbonates concentration of the ECF. This loss of bicarbonates from the ECF could be responsible for the significant reduction in the bicarbonate levels of the diabetic animals.
In the ECF, an inverse relationship is known to exist between the bicarbonates and chlorides in order to keep the anion concentration constant. Thus a decrease in the bicarbonate concentration in the ECF will likely cause an increase in the concentration of chlorides. This increase in ECF chloride concentration may therefore account for the difference in the chloride concentration observed in the diabetic animals as observed in Table 2. However, none of the concentrations of the extract administered significantly ameliorated the altered K+ and Na+ levels of the diabetic animals compared with the diabetic control (p>0.05), 40 and 60% of the extract were most effective in ameliorating the altered bicarbonate levels of the diabetic with respect to the diabetic control while all the concentrations of the extract administered significantly ameliorated the altered chloride levels of the diabetic animals compared with the diabetic control and non diabetic (p<0.05).
The liver is the most vital organ for the metabolism of drugs and other toxicants. The destruction of the liver cell results in the impairment of the liver cell membrane permeability which results in the leakage of tissue contents into the blood stream (Saeed et al., 2008). In addition, physical trauma or disease process can cause lyses, resulting in the release of these intracellular enzymes into the blood. It has been reported that the liver is necrotized in diabetic rats which leads to increased activities of AST, ALT and Alkaline Phosphatase enzymes as they leak from the liver cytosol into the blood stream (Saeed et al., 2008) and this is also an indicator of the hepatotoxicity of alloxan. Therefore, the increase in the activities of AST, ALT and ALP in serum of the diabetic animals as observed in Table 3, is mainly due to the leakage of these enzymes from the liver cytosol into the blood stream (Mansour et al., 2002; Whitehead et al., 1999).
| Table 4: | Effect of mistletoe extract on the body weight of diabetic and non-diabetic rats (g) |
 | |
n = 4 animals per group | |
| Table 5: | Percentage change in glucose, weight and growth rate |
 | |
Phosphatase activity is normally high in diseased states and is often used as a tool in clinical investigations (Bull et al., 2000). Data generated from this study indicates that all concentrations of the extract administered could significantly ameliorate the altered phosphatase and transaminase activities of the diabetic rats (p<0.05) (Table 3).
The loss in weight in the diabetic groups as observed in Table 4, is attributed to the alloxan that was injected into the animals. Alloxan is known to destroy the β-cells of the islets of the langerhams of the pancreas that function in insulin regulation, producing type 1 diabetes (Eleazu et al., 2010). The destruction of the pancreas results in the utilization of non-carbohydrate moieties such as protein for the synthesis of glucose. The loss of structural proteins in increased gluconeogenesis together with increased lipolysis and increased synthesis of ketone bodies results in severe weight loss. All the concentration of the extracts that were administered ameliorated the weight loss observed in the diabetic animals compared with the control.
There was a 12.23±3.87, 8.47±2.69 and 2.95±1.44% increase in weights of the diabetic rats respectively, after administration of 20, 40 and 60% extracts of mistletoe to the diabetic rats. Similarly, we recorded a 44.97±6.82, 32.57±2.73 and 10.68±3.54% increase in growth rates after administration of 20, 40 and 60% extracts of mistletoe respectively to the diabetic rats while we recorded a 42.88±0.52, 21.47±0.23 and 16.54±3.37% decrease in fasting blood glucose levels after administration of 20, 40 and 60% extracts of mistletoe respectively to the diabetic animals (Table 5). The study shows that 20% of the aqueous extract of mistletoe flour was most effective in ameliorating the hyperglycemic status of the diabetic rats with a corresponding increase in body weights and growth rates.
CONCLUSION
The result of the study carried out reveals that mistletoe has anti-diabetic potentials and could be useful in improving the altered electrolytes that arise from diabetes. Finally, the dietary supplement could protect the liver cells from free radical damage that arises from diabetes.
REFERENCES
- Aldhahi, W. and O. Hamdy, 2003. Adipokines, inflammation and the endothelium in diabetes. Curr. Diab. Rep., 3: 293-298.
CrossRefPubMedDirect Link - Cooper, G.R., 1973. Methods for determining the amount of glucose in blood. CRC Crit. Rev. Clin. Lab. Sci., 4: 101-145.
CrossRefPubMedDirect Link - Deeni, Y.Y. and N.M. Sadiq, 2002. Antimicrobial properties and phytochemical constituents of the leaves of African mistletoe (Tapinanthus dodoneifolius (DC) Danser) (Loranthaceae): An ethnomedicinal plant of Hausaland, Northern Nigeria. J. Ethnopharmocol., 83: 235-240.
CrossRefPubMedDirect Link - Dohi, T., K. Kawamura, K. Morita, H. Okamola and A. Tsiyimoto, 1988. Alterations of the plasma selenium concentrations and the activities of tissue peroxide metabolism enzymes in streptozotocin-induced diabetic rats. Horn. Metab. Res., 20: 671-675.
CrossRefPubMedDirect Link - Eleazu, C.O., P.N. Okafor and I. Ahamefuna, 2010. Total antioxidant capacity, nutritional composition and inhibitory activity of unripe plantain (Musa paradisiacae) on oxidative stress in alloxan induced diabetic rabbits. Pak. J. Nutr., 9: 1052-1057.
CrossRefDirect Link - Henry, R.J., 1974. Clinical Chemistry: Principles and Techniques. 2nd Edn., Harper and Row, New York, USA., ISBN-13: 9780061411816, Pages: 1629.
Direct Link - Henry, R.J., N. Chiamori, O.J. Golub and S. Berkman, 1960. Revised spectrophotometric methods for the determination of glutamic-oxalacetic transaminase, glutamic-pyruvic transaminase and lactic acid dehydrogenase. Am. J. Clin. Pathol., 34: 381-398.
CrossRefPubMedDirect Link - King, H., R.E. Aubert and W.H. Herman, 1998. Global burden of diabetes, 1995-2025: Prevalence, numerical estimates and projections. Diabetes Care, 21: 1414-1431.
CrossRefPubMedDirect Link - Mansour, H.A., A.S.A. Newairy, M.I. Yousef and S.A. Sheweita, 2002. Biochemical study on the effects of some Egyptian herbs in alloxan-induced diabetic rats. Toxicology, 170: 221-228.
CrossRefPubMedDirect Link - Saeed, M.K., Y. Deng and R. Dai, 2008. Attenuation of biochemical parameters in streptozotocin-induced diabetic rats by oral administration of extracts and fractions of Cephalotaxus sinensis. J. Clin. Biochem. Nutr., 42: 21-28.
Direct Link - Obatomi, D.K., E.O. Bikomo and V.J. Temple, 1994. Anti-diabetic properties of the African mistletoe in streptozotocin-induced diabetic rats. J. Ethnopharmacol., 43: 13-17.
Direct Link - Stoner, G.D., 2005. Hyperosmolar hyperglycemic state. Am. Family Physician, 71: 1723-1730.
Direct Link - Morelli, V. and R.J. Zoorob, 2000. Alternative therapies: Part I. depression, diabetes, obesity. Am. Family Physician, 62: 1051-1060.
Direct Link - Wild, S., G. Roglic, A. Green, R. Sicree and H. King, 2004. Global prevalence of diabetes: Estimates for the year 2000 and projections for 2030. Diabetes Care, 27: 1047-1053.
CrossRefPubMedDirect Link - Whitehead, M.W., N.D. Hawkes, I. Hainsworth and J.G.C. Kingham, 1999. A prospective study of the causes of notably raised aspartate transaminase of liver origin. Gut, 45: 129-133.
CrossRefPubMedDirect Link - Yared, Z. and J.L. Chiasson, 2003. Ketoacidosis and the hyperosmolar hyperglycemic state in adult diabetic patients. Diagnosis and treatment. Minerva Med., 94: 409-418.
PubMedDirect Link