
M.K. Somda
Centre de Recherche en Sciences Biologiques Alimentaires et Nutritionnelles (CRSBAN), Universite de Ouagadougou (Burkina Faso), 03 BP: 7131 Ouagadougou, 03/Burkina Faso
A. Savadogo
Centre de Recherche en Sciences Biologiques Alimentaires et Nutritionnelles (CRSBAN), Universite de Ouagadougou (Burkina Faso), 03 BP: 7131 Ouagadougou, 03/Burkina Faso
C.A.T. Ouattara
Centre de Recherche en Sciences Biologiques Alimentaires et Nutritionnelles (CRSBAN), Universite de Ouagadougou (Burkina Faso), 03 BP: 7131 Ouagadougou, 03/Burkina Faso
A.S. Ouattara
Centre de Recherche en Sciences Biologiques Alimentaires et Nutritionnelles (CRSBAN), Universite de Ouagadougou (Burkina Faso), 03 BP: 7131 Ouagadougou, 03/Burkina Faso
A.S. Traore
Centre de Recherche en Sciences Biologiques Alimentaires et Nutritionnelles (CRSBAN), Universite de Ouagadougou (Burkina Faso), 03 BP: 7131 Ouagadougou, 03/Burkina Faso
Asian Journal of Biotechnology
Year: 2011 | Volume: 3 | Issue: 1 | Page No.: 77-83







ABSTRACT
The use of mango peel (agricultural waste with no appreciable value to industries or competitive use as food) as alternative and cost-effective biomass for the production of bioethanol was examined. Thus, the aim of this study was to select performed yeasts strains able to ferment sugars containing in peel mango. The methods used, included: enzymatic hydrolysis with Bacillus and simultaneous saccharification and fermentation with W6 and B1, yeasts isolated from wine and among bakers yeasts, respectively. The kinetics parameters results showed that the optimum alcohol releasing like 35% (g E/g S) and 30% (g E/g S) respectively from W6 and B1 be spread out at 40°C. The maximum volume of ethanol produced from peel mango by W6 and B1 were, respectively 13 and 10.1 g L ¯1 at the 120th.
PDF Abstract XML References Citation
Received: September 11, 2009;
Accepted: May 20, 2010;
Published: July 08, 2010
How to cite this article
M.K. Somda, A. Savadogo, C.A.T. Ouattara, A.S. Ouattara and A.S. Traore, 2011. Thermotolerant and Alcohol-Tolerant Yeasts Targeted to Optimize Hydrolyzation from Mango Peel for High Bioethanol Production. Asian Journal of Biotechnology, 3: 77-83.
DOI: 10.3923/ajbkr.2011.77.83
URL: https://scialert.net/abstract/?doi=ajbkr.2011.77.83
DOI: 10.3923/ajbkr.2011.77.83
URL: https://scialert.net/abstract/?doi=ajbkr.2011.77.83
INTRODUCTION
Tolerance to high temperatures and high ethanol concentrations are important properties of microorganisms of interest to industry. The fermentation efficiency of S. cerevisiae at high temperatures is very low due to increased fluidity in membranes to which the yeast responds by changing its fatty acids composition (Ohta et al., 1988; Van Uden, 1984). For an efficient conversion of fibrous biomass to ethanol, another desirable microorganism characteristic is the capacity to ferment other sugars besides glucose, such as d-xylose and cellobiose obtained from hydrolysis of the material (Banat et al., 1998; Miller, 1959).
The microorganisms employed in the fermentation of sugars from raw biomass into ethanol are principally bacteria and yeasts (Almeida et al., 2007; Klinke et al., 2004). The Saccharomyces cerevisiae yeast has proved to be more robust than bacteria, being more tolerant to ethanol and inhibitors present in hydrolysates of raw biomass materials like cellulosic (Olson and Hahn-Hagerdal, 1996).
Due to their potential applications, thermotolerant yeasts at temperatures above 35°C have been the object of many studies (Szczodrak and Targonsky, 1988; Liu et al., 2009).
Stress or environmental stimuli can cause structural changes and/or metabolic changes in an organism acting as expression activator for genes involved in the synthesis of specific compounds that protect the organism (Banat et al., 1998). Abiotic factors can be temperature, osmotic stress, anaerobic conditions, heavy metals, growth regulators, ultraviolet radiation, metabolic repressors and pH (Chen and Chen, 2004; Rajoka et al., 2005; Cakar et al., 2005).
Stress due to temperature has been the most studied abiotic factor, where both heat and cold induce the synthesis or storing of a group of proteins that increase stress resistance (Hiraishi et al., 2006; Yacoob and Filion, 1987). Most of the time yeasts performances are researched to valorize biomass in this way it is necessary to find thermotolerant and alcohol-tolerant yeasts. Among the biomass researched, carbohydrates coming from mango are mainly important in Burkina Faso. And yet hydrolization of mango peel could optimize carbohydrates rate.
During processing of mango, byproducts such as peel and kernel are generated. Peel contributes about 15-20% of the fruit (Beerh et al., 1976). As peel is not currently utilized for any commercial purposes, it is discarded as a waste and becoming a source of pollution. This waste should be treated as a specialized residue due to high levels.
The objective of this study was to select yeasts strains able to ferment sugars containing in peel mango pre-treated at high temperature and also able to resist alcoholic degree above.
MATERIALS AND METHODS
Yeast strains and media: The selection was carried out on a total of 20 yeast strains. The strains codified as W1 to W10 were S. cerevisiae strains of wild origin isolated from wine cultures. Also the strains codified as B1 to B10 the Baker’s yeast microorganism commonly used in industrial wine making. These strains were selected in Laboratory of Biotechnology and Microbiology during 2009.
The maintenance medium (used in agar plates) contained 20 g of glucose, 20 g of agar, 5 g of peptone, 5 g of MgSO4•7H2O per liter. The growth medium utilized in the liquid inoculation contained 50 g glucose, 5 g of yeast extract, 1 g of KH2PO4, 0.3 g of NH4Cl and 2 g of MgSO4•7H2O per liter. The fermentation medium in 0.05 M citrate buffer pH 4.8; 1.5 g peptone, 5 g of yeast extract, 1 g of KH2PO4, 0.3 g of NH4Cl and 2 g of MgSO4•7H2O per liter (Reference:Standard medium for yeasts growth).
Growth conditions: The evaluation of biomass was carried out in an orbital shaker at 150 rpm at different temperatures and times in 250 mL Erlenmeyer flasks, where each one contained 50 mL of growth medium.
The flasks were inoculated with a cell suspension at 5% (v/v) with 5x105 cells mL-1. The biomass increase was determined by gravimetric and the results were expressed in mg in dry base.
Acclimatization treatment: The first selections of strains were carried out on Petri dishes with the maintenance medium by incubating them in an oven at 35, 40, 42, 45 and 50°C to isolate thermotolerant strains during 72 h. Biomass production and ethanol yield were evaluated. The fermentation yield was calculated assuming that 0.51 g ethanol/glucose in the culture medium. The verification of the thermotolerant qualities of the cultures obtained after acclimatization were performed by growing the yeasts on Petri dishes in the maintenance medium and incubating at 40, 42, 45 and 50°C.
Alcohol treatment: The same yeasts submitted to acclimatization treatment have been used alcohol treatment. Fermentation was conducted to 10, 12, 14, 15 and 18% (v/v) of alcohol (ethanol). Biomass production and ethanol yield were evaluated. The alcohol tolerant qualities verification of cultures is obtained by growing the yeasts on Petri dishes.
Colony selection: Colonies from strains that grew and produced ethanol at 40, 42 and 45°C and supported alcohol 15% were isolated from different biotope, in a maintenance medium; and evaluated by measuring the CO2 displacement in a growth medium in a standard Hungate tube saccharimeter at 40°C and 24 h. The homogeneous cultures that achieved displacement above 30 mm were evaluated measuring the glucose consumption and production of biomass and ethanol. The concentration of reducing sugars was determined by dinitrosalicylic (DNS) colorimetric assay using glucose as sugar standard (Miller, 1959). The ethanol concentration was determined by distillation method (Humphrey and Okafoagu, 2007).
Dry weights were determined by drying yeast cells from 5 mL samples, for 24 h at 65°C, after centrifugation at 2000 g for 10 min and washing twice with 0.9% NaCl (w/v). All determinations were done by means of standard curves; results were the mean of three repetitions.
Peel liquefaction optimization: The hydrolysis was conducted at 55°C, with α-amylases produced by Bacillus A1M1 (Lab., of Poitiers). The peel hydrolysis performance was evaluated based on the liquefaction yield (g-reducers sugars/g-substrate).
Initially 1 L slurries containing 50 g L-1 of mango substrate (peel) were prepared in distilled water and α-amylases from Bacillus were added; the samples were hydrolysed at the suitable temperature for 4 h with mild agitation (100 rpm).
Simultaneous saccharification and fermentation of mango substrate: The simultaneous peel hydrolyze and fermentation was carried out in a 500 mL Erlenmeyer flask at 10% substrate consistency in the SFF medium containing peel mango pre-treated as substrates (200 mg L-1 potassium metabisulphite). After liquefaction, the pH was adjusted to 4.5, potassium metabisulphite and α-amylases from Bacillus have been added and temperature growing to 55°C to optimize reducers sugars releasing. At last yeasts inoculums (5% v/v with 2.5x106 cells mL-1) from the selected strains were added independently for each flask at 40°C. These yeasts were thermotolerant. SFF was performed at 40°C for 7 days at 150 rpm. The ethanol concentration was determined during fermentation process.
The theoretical SFF yield was calculated by assuming that all the potential glucose in the pretreated material was available for fermentation. The experiments were performed in triplicate.
Determination of ethanol
Determination of quantity of ethanol produced: The distillate collected over a slow heat at 78°C was measured using a measuring cylinder and expressed as the quantity of ethanol produced in g L-1 by multiplying the volume of distillate collected at 78°C by the density of ethanol (0.8033 g mL-1). The G L-1 is equivalent to the yield of 100 g of dried substrate (Humphrey and Okafoagu, 2007).
Determination of percentage ethanol concentration: A standard ethanol density curve was prepared by taking series of percentage (v/v) ethanol solutions, which were prepared in volumetric flasks and the weight was measured. The density for each of the prepared ethanol solutions was calculated and a standard curve of density against percentage ethanol was plotted. The percentage ethanol concentration of ethanol produced was obtained by comparing its density with the standard ethanol density curve (600 nm).
RESULTS
Screening of thermotolerant and alcohol tolerant strains: All the yeasts grew in the maintenance medium at 35 and 40°C also resist at alcohol degree until 14°C. Only two strains (W2; B1) grew at 42°C and strains (B1, W5; W6) at 45°C.
| • | Selection of thermotolerant and Alcohol tolerant strains for ethanol production |
The yeasts W6 and B1 present good yield of alcohol bioconversion respectively between 0.35 and 0.30 (g g-1).
| • | Peel liquefaction optimization |
| • | Ethanol production from peel mango sugar performed by simultaneous saccharification and fermentation (SSF) |
DISCUSSION
Screening of thermotolerant and alcohol tolerant strains: Only two strains (W2; B1) grew at 42°C and strains (B1, W5; W6) at 45°C. These strains could synthesis or storing of a group of proteins that increase stress resistance to the high temperature and alcohol degree. The same phenomenal resistance has been observed and explained by Hiraishi et al. (2006).
Selection of thermotolerant and alcohol tolerant strains for ethanol production: The strains that grew at 40, 42 and 45°C in the maintenance medium were incubated in the growth medium and the biomass and ethanol production were evaluated at 72 h in the same temperatures. These kinetics parameters results showed that the optimum alcohol releasing like 35% (g E/g S) and 30% (g E/g S), respectively from W6 and B1 be spread out at 40°C for the both kind of yeasts. In opposite the biomass get lower level. The thermopile nature of strains W6 and B1 is shown in Table 1. These two yeasts are indeed able to continue a fermentation rate close to normal beyond 37°C. This property could possibly be utilized simultaneously with amylase from Bacillus for peel substrate fermentation. The similar process using simultaneous yeast and amylase was realized on starch (Jamai et al., 2007). At finally they found 49% (g E/g S).
Peel liquefaction optimization: The hydrolysis was conducted up to 4 h on mango peel containing 22.62% (g g-1) of carbohydrates and the results shown in Fig. 1, shows that releasing of reducers sugars increased with enzymatic activity of Bacillus. Likewise it is noticed that there has not significant change or variation of reducers sugars rate without enzyme presence.
| Table 1: | Kinetics parameters for two yeasts at 40, 42 to 45°C at 72 h and average value of 3 replicates |
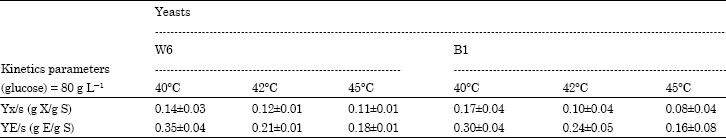 | |
| Yx/s (g X/g S): Yield of biomass (g) produced by substrate consumed (g). YE/s (g E/g S): Yield of ethanol (g) produced by substrate consumed (g). X: Biomass, S: Substrate, E: Ethanol | |
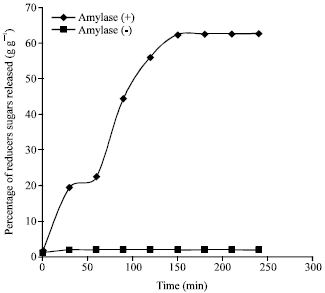 | |
| Fig. 1: | Following of reducers sugars percentage releasing through carbohydrates hydrolysis on peel mango, incubated with α-amylase of Bacillus |
The liquefaction conduct at middle temperature (55°C) result a higher yield of reducers sugars released (62% g g-1), which might be related to the type of enzyme utilized.
Thus fermentescibles sugars percentage obtained by using Bacillus enzymatic activity is relatively very higher than enzyme absence (2% g g-1). It can be explained that the enzyme has the capacity of decomposing into hexose all polysaccharides which are built up of glucose residues united by α-1,4 glycosidic bonds and also it is thermostable. Therefore this potentiality Bacillus could be used to fermentation process in medium containing polysaccharides substrates like mango peel.
Ethanol production from peel mango sugar performed by Simultaneous Saccharification and Fermentation (SSF): Peel mango was used to produce ethanol through enzymatic hydrolysis and SSF with, respectively two yeasts strains (W6, B1). In SSF, the two different microorganisms behaved differently, according to their nutrient requirements, but synergistically in the degradation of organic substrate. An enzyme (carbohydrate hydrolases produced by Bacillus A1M1, Lab., of Poitiers) was able to hydrolyze peel mango. The saccharification products were simultaneously utilized by yeasts strains (W6, B1) for ethanol production. These yeasts are able to produced ethanol due to the presence of pyruvate decarboxylase (PDC) and alcohol dehydrogenase (ADH), which are key enzymes in ethanol formation, as reported by Gunasekaran and Chandra (2007). Figure 2 shows that the maximum volume of ethanol (13 g L-1) produced from peel mango by W6 and B1 (10.1 g L-1) in this study at the 120th h is lower than the results found by researchers, who also reported maximum ethanol yield at 120th h from fresh fruit (64.01 g L-1) and waste fruits (21.14 g L-1) using Z. mobilis. The higher ethanol yield from fresh fruit was due to higher presence of fructose and glucose in fresh fruits, as stated by Micheal and Rosaline (2000). The maximum volume of ethanol produced from peel mango is lower than the 59 g L-1 reported by Gunasekaran and Chandra (2007) at 120th h from cassava starch hydrolysate.
This is due to cassava containing more carbohydrates, which could be fermented to ethanol. Present results are in agreement with those found previously by these authors quoted.
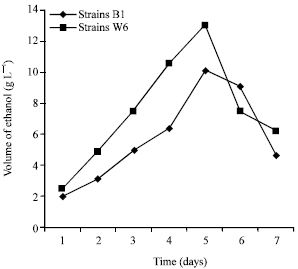 | |
| Fig. 2: | Ethanol produced (g L-1) from mango peel using yeasts strains B1 and W6 separately and each with amylase presence simultaneously |
The results revealed that ethanol could be produced from agricultural residues, such as peel mango using thermotolerants and alcohol tolerants yeasts strains (W6, B1) as fermenting organisms. Considering the cost-effectiveness, in addition to being a means to control environmental pollution, the use of peel mango for ethanol production is concluded as a worthwhile venture.
ACKNOWLEDGMENT
Author is grateful to Dr. Aly Savadogo for his guidance. Also my thank to Pr. Alfred S. Traore Director of CRSBAN where I done research training.
REFERENCES
- Almeida, J., T. Modig, A. Petersson, H. Hn-Ha, B. Gerdal, G. Liden and M. Gorwa-Grauslund, 2007. Increased tolerance and conversion of inhibitors in lignocellulosic hydrolysates by Saccharomyces cerevisiae. J. Chem. Technol. Biotechnol., 82: 340-349.
CrossRefDirect Link - Banat, I., P. Nigam, D. Singh, R. Marchant and A. McHale, 1998. Review: Ethanol production at elevated temperatures and alcohol concentrations: Part I- Yeasts in general. World J. Microbiol. Biotechnol., 14: 809-812.
CrossRefDirect Link - Cakar, Z., U. Seker, C. Tamerler, M. Sonderegger and U. Sauer, 2005. Evolutionary engineering of multiple-stress resistant Saccharomyces cerevisiae. FEMS Yeast Res., 5: 569-578.
PubMed - Chen, K. and Z. Chen, 2004. Heat shock proteins of thermophilic and thermotolerant fungi from Taiwan. Bot. Bull. Acad. Sinica, 45: 247-256.
Direct Link - Hiraishi, H., M. Mochizuki and H. Takagi, 2006. Enhancement of stress tolerance in Saccharomyces cerevisiae by overexpression of ubiquitin ligase Rsp5 and ubiquitin-conjugating enzymes. Biosci. Biotechnol. Biochem., 70: 2762-2765.
PubMed - Humphrey, C.N. and U.C. Okafoagu, 2007. Optimization of ethanol production from Garcinia kola (bitter kola) pulp agrowaste. Afr. J. Biotechnol., 6: 2033-2037.
Direct Link - Klinke, H., A. Thomsen and B. Ahring, 2004. Inhibition of ethanol-producing yeast and bacteria by degradation products produced during pre-treatment of biomass. Applied Microbiol. Biotechnol., 66: 10-26.
CrossRefDirect Link - Jamai, L., K. Ettayebi, J.E.L. Yamani and M. Ettayebi, 2007. Production of ethanol from starch by free and immobilized Candida tropicalis in the presence of α-amylase. Bioresour. Technol., 98: 2765-2770.
CrossRef - Liu, L.Z., M. Menggen and S. Mingzhou, 2009. Evolutionarily engineered ethanologenic yeast detoxifies lignocellulosic biomass conversion inhibitors by reprogrammed pathways. Mol. Genet. Genomics, 282: 233-244.
PubMed - Miller, G.L., 1959. Use of dinitrosalicylic acid reagent for determination of reducing sugar. Anal. Chem., 31: 426-428.
CrossRefDirect Link - Ohta, K., S. Wijeyaratne and S. Hayashida, 1988. Temperature-sensitive mutants of a thermotolerant yeast, Hinsenula polymorpha. J. Fermentation Technol., 66: 455-459.
CrossRef - Olsson, L. and B. Hahn-Hagerdal, 1996. Fermentation of lignocellulosic hydrolysates for ethanol production. Enzyme Microb. Technol., 18: 312-331.
CrossRefDirect Link - Rajoka, M., A. Khalid and M. Ferhan, 2005. Kinetic and thermodynamics of ethanol production by a thermotolerant mutant of Saccharomyces cerevisiae. Lett. Applied Microbiol., 40: 316-321.
PubMed - Szczodrak, J. and Z. Targonsky, 1988. Selection of thermotolerant yeast strains for simultaneous saccharification and fermentation of cellulose. Biotechnol. Bioeng., 31: 300-303.
CrossRefDirect Link - Yacoob, K. and W. Filion, 1987. The effect of cold temperature stress on gene expression in maize. Biochem. Cell. Biol., 65: 112-119.
Direct Link