
A.H. Gorji
Department of Plant Breeding, Islamic Azad University, Broujerd Branch, Broujerd, Iran
M. Zolnoori
Department of Plant Breeding, Islamic Azad University, Broujerd Branch, Broujerd, Iran
Asian Journal of Biotechnology
Year: 2011 | Volume: 3 | Issue: 4 | Page No.: 368-377







ABSTRACT
The aim of this study was to molecularly characterize the thirty-eight wheat cultivars recommended for various regions of Iran and to assess mutual genetic distances, through the use of microsatellite markers. Polymorphic information content (PIC value) calculated to assess the informativeness of each marker ranged from 0.19 to 0.78 with a mean of 0.49.Twenty three Polymorphic Microsatellite Markers (PMM) delineated all 34 of the samples, revealing a total of 73 Simple Sequence Repeat (SSR) alleles, i.e., an average of 3.2 alleles per locus. Genetic distances among the 38 genotypes ranged from 0.11 (between genotypes S-83-20 and N-83-3 ) to 0.88 (between genotypes D-83-2 and M-83-5) the mean distance being 0.47. Thirteen groups were obtained by using the unweighted pair-group method with arithmetic means analysis (UPGMA) and fourteen through the Tocher method. Both methods produced similar clusters with one to thirteen genotypes per group. Twenty three polymorphic microsatellite markers delineated all 38 of the samples, revealing a total of 37 simple sequence reapt (ssr) alleles, i.e., an average of 3.1 alleles per locus. The results indicate that these tools may be used to protect intellectual property and for breeding and selection programs.
PDF Abstract XML References Citation
Received: October 22, 2010;
Accepted: February 08, 2011;
Published: June 15, 2011
How to cite this article
A.H. Gorji and M. Zolnoori, 2011. Genetic Diversity in Hexaploid Wheat Genotypes using Microsatellite Markers. Asian Journal of Biotechnology, 3: 368-377.
DOI: 10.3923/ajbkr.2011.368.377
URL: https://scialert.net/abstract/?doi=ajbkr.2011.368.377
DOI: 10.3923/ajbkr.2011.368.377
URL: https://scialert.net/abstract/?doi=ajbkr.2011.368.377
INTRODUCTION
The primitive cultivars, landraces and wild relatives of crop plants constitute a pool of useful genetic variability required for the effective breeding programs. Most of the rich plant biodiversity which supported agriculture for the past 9000 years has been eroded or being rapidly eroded due to the introduction of new High Yielding Varieties (HYVs) The danger of erosion of genetic resources was recognized as early as 1974 when the Consultative Group on International Agricultural Research established International Board of Plant Genetic Resources with a view to develop a global network of genetic resources centers. Genetic variability of the Plant Genetic Resources of a country conditions breeder’s activities and the development of cultivars with improved adaptation. In order to safeguard this genetic heritage, germplasm collections of many important crops are currently conserved ex situ in genebanks. In many cases, this is the only possible type of conservation because of social and economical impediments. However, ex situ collections may have a considerable percentage of duplicate accessions as well as entries misclassified (erroneous identification or accidental mixing) that difficult their effective conservation and use in breeding. The proper maintenance of genetic variation including heterogeneity of the accessions is one of the major aspects of genebank management. To resolve these problems it is essential to monitor intra- and inter-cultivar variability using polymorphic markers for genotype identification. Molecular markers proved to be an important way to increase selection efficiency and there are good prospects for marker-assisted selection in improving drought responses in wheat (Quarrie et al., 2003).
Genetic diversity is the basis for genetic improvement of plants. Knowledge of germplasm diversity has a significant impact on the improvement of crop plants. Due to modern breeding, it has been suggested that genetic diversity in wheat has been increasingly narrowed for example Fu and Somers (2009) conducted a genome-wide examination of 75 Canadian hard red spring wheat (Triticum aestivum L.) cultivars released from 1845 to 2004 using 370 SSR markers that were widely distributed over all 21 wheat chromosomes and indicated that genetic diversity in this wheats increasingly narrowed.
Narrow genetic diversity is a problem in breeding for adaptation to biotic stresses, like diseases and abiotic stresses, such as drought or salt tolerance. Therefore, it is necessary to investigate the genetic diversity in wheat germplasm in order to broaden the genetic variation in future breeding programme. Morphological traits can be used for assessing genetic diversity but are often influenced by the environment. The use of molecular markers for the evaluation of genetic diversity is receiving much attention. Microsatellites, also termed simple sequence repeats (SSRs) have been proposed as one of the most suitable markers for the assessment of genetic variation and diversity among wheat varieties/lines, because they are multiallelic, chromosome-specific and evenly distributed along chromosomes (Roder et al., 1998). Wheat microsatellites have been successfully used for the construction of genetic linkage maps of wheat (Elouafi and Nachit, 2004). Microsatellite markers are useful and becoming popular for different applications in wheat breeding due to their high level of polymorphism and easy handling and are used to evaluate genetic diversity of hexaploid wheat (Triticum aestivum L.) landraces in relation to their geographic origin (Al-Khanjari et al., 2007). Ijaz and Khan (2009) estimated the genetic divergence among wheat accessions as well as cultivated varieties of Pakistan with the help of SSR markers. This study addressed the utilization of microsatellite markers, to determine genetic diversity and relationships at the molecular level, among 63 genotypes of wheat. Ciuca and Petcu (2009) studying the same markers (SSR) in doubled haploid lines derived from a cross between cultivars Izvor and Jiana, used as checks in our study, found that markers Xwmc9, Xwmc596 and Xwmc603 cosegregated, showing a complete correspondence between grouping the lines according to the alleles of the three markers. The objectives of the present study were to examine the genetic factors that affected microsatellite diversity and to use wheat microsatellite markers for characterization and assessment of the genetic diversity of a set of hexaploid wheat germplasms from the gene bank.
MATERIALS AND METHODS
The experiment was conducted in Department of Plant Breeding, Islamic Azad University, Broujerd Branch, Iran during fall and winter season 2009. In this study, we evaluated genetic diversity within a representative compilation of the germplasm of Iranian wheat, in order to assess allelic frequencies in an informative set of microsatellite loci, thereby permitting a more accurate identification of varieties of Iranian wheat. Thirty-eight genotypes, developed at various research centers, both private and public, were used (Table 1). DNA extraction from the seeds was undertaken according to the protocol described by McDonald et al. (1994) but with certain adjustments. Forty seeds from each cultivar were ground utilizing a MA 048 mill (Marconi) and 60 mg sub-samples of flour collected therefrom. DNA concentration was determined spectrophotometrically (Sambrook et al., 1989).
| Table 1: | List bread wheat cultivars and their pedigree |
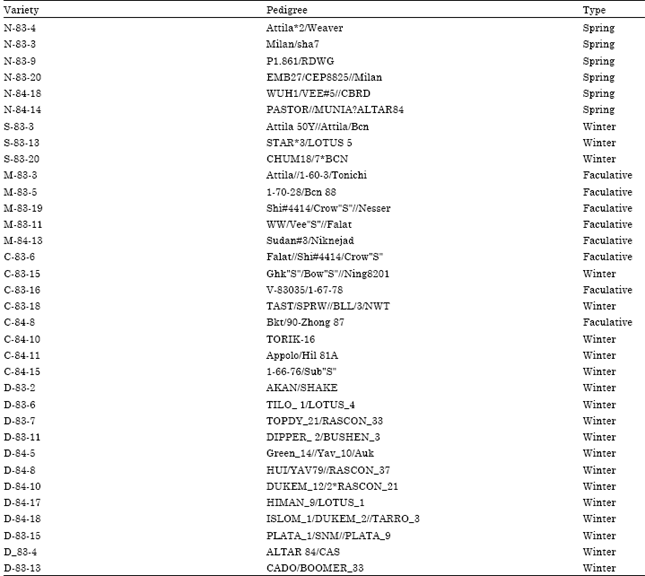 | |
| * = Unmapped loci, ? = Unknown position | |
Microsatellite-loci amplification was performed by using DNA samples in a 40 μL volume containing 125 mM KCl, 25 mM Tris-HCl (pH 8.3), 5 mM MgCl2, 0.8 μM of each primer, one unit of Taq DNA polymerase enzyme, 250 μM of each of the deoxynucleotides (dATP, dTTP, dGTP and dCTP) and 150 ng of DNA. Amplification was achieved with a 8 min initial step at 94°C followed by 40 cycles of 1 min at 94°C, 1 min at 55°C, 2 min at 72°C and finally a 7 min step at 72°C after the 40th cycle. Amplified fragments were separated and revealed on 7% polyacrylamide denaturing gels stained with silver-nitrate. The Polymorphic Information Content (PIC) of each microsatellite locus was evaluated through allelic frequency:
where, pij represents the frequency from the jth allele to the ith primer (Anderson et al., 1993).
Genetic relationships among accessions were evaluated through a dissimilarity matrix, using the complement of a similarity index for multi-allelic data and co-dominant with the aid of GENES software (Cruz, 2006).
From dissimilarity assessments, genotypes were clustered by means of the hierarchical unweighted pair-group method with arithmetic means analysis (UPGMA), Tocher optimization method, Ward's method and single linkage and complete methods. A cophenetic correlation analysis between the original dissimilarity matrix and the dissimilarity matrices obtained from dendograms was carried out to define the hierarchical clustering method which best represented the original data. Clustering analysis using hierarchical methods was undertaken with STATISTICA software (StatSoft Inc Tulsa, OK, USA) and Tocher optimization clustering analysis with GENES software (Cruz, 2006).
RESULTS
Besides eight unmapped loci (Table 2), 12 genome-A loci, 13 genome-B loci and 10 genome-D loci were evaluated. From these 43 loci, 23 presented polymorphisms among the 38 wheat genotypes analyzed (54%). In these 23 polymorphic loci, the numbers of alleles observed per locus ranged from two to five, making a total of 75 alleles with a mean of 3.26 alleles per locus. PIC values calculated in order to estimate the informativeness of each polymorphic locus, varied from 0.19 to 0.78 with a mean of 0.49 (Table 2).
The variability’s obtained in A, B and D genomes were similar. In genome A, six out of the twelve loci evaluated were polymorphic (50%), with 21 alleles (3.5 alleles per locus), whereas, in genome B, seven out of thirteen loci were polymorphic (54%), with 18 alleles (2.6 alleles per locus) and in genome D, six out of ten SSR loci were polymorphic (60%), with 18 alleles (3 alleles per locus).Among the eight unmapped SSR loci, four were polymorphic (50%), with 18 alleles (4.5 alleles per locus). PIC values were 0.54; 0.49; 0.40 and 0.60 for A, B and D genomes, as well as the unmapped loci, respectively. Genetic distances among the genotypes ranged from 0.10, between S-83-20 and N-83-3, to 0.88 between D-83-2 and M-83-5, with a mean distance of 0.48. The highest frequencies of genetic distance occurring between 0.40 and 0.50 (Fig. 1).
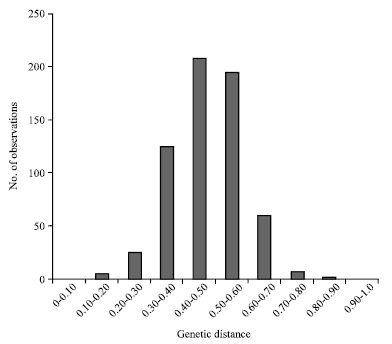 | |
| Fig. 1: | Frequency distribution of genetic distance obtained in thirty-eight wheat cultivars with twenty-three SSR polymorphic loci |
| Table 2: | Microsatellite markers used in assessment of genetic diversity of wheat |
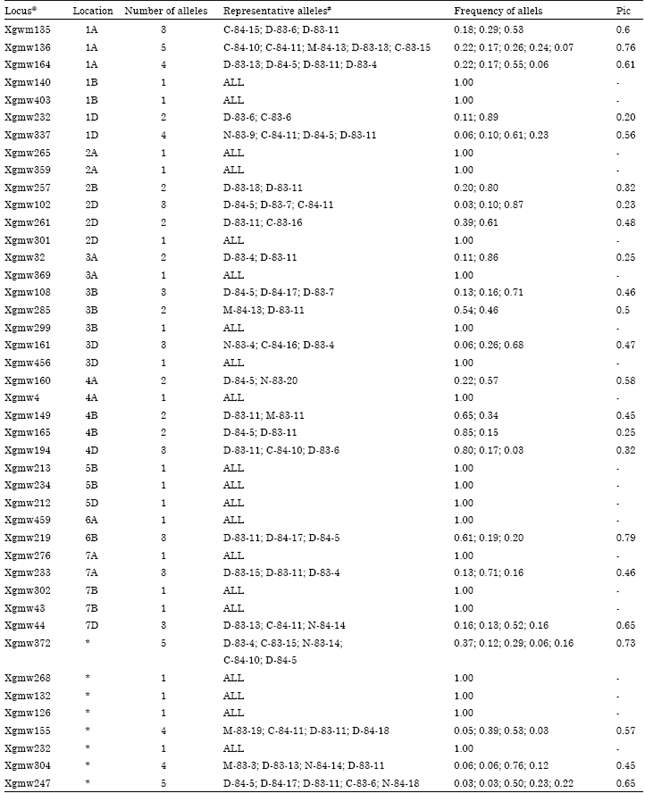 | |
| *Unmapped loci. #Each genotypes represents the allele whose frequency is shown in the afterward column in the same order | |
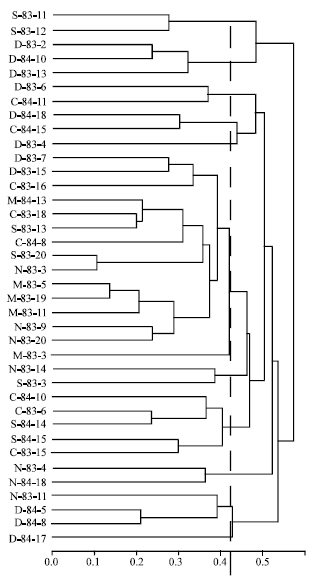 | |
| Fig. 2: | Clustering assessment obtained by UPGMA based on dissimilarity measurement of thirty-eight iranian wheat genotypes. The hatched line indicates the cutogg for cluster formation |
Table 1 indicates the pedigree of the 38 genotypes. A group of three genotypes (M-83-5,M-83-19 and M-83-11) and another pair of genotypes (N-83-9 and N-83-20) could not be distinguished by pedigree. There were 69 different parents in the genealogy of these wheat genotypes, of which a few participated in more than one pedigree. From the 628 parentage coefficients (CP) possible between these 38 genotypes, only 25 were different from zero. Therefore, it was not possible to correctly group these cultivars by genealogy alone. By using molecular markers, it was possible to distinguish all 38 genotypes and group these by genetic similarity. We used four hierarchical methods. Cophenetic correlations obtained between the original dissimilarity matrix and matrices from dendograms were 66, 43, 45 and 52% for the UPGMA, Ward's, complete-linkage and single-linkage methods, respectively. Due to its higher degree of cophenetic correlation, the UPGMA approach (Fig. 2) was chosen for graphical representation of genotype clusters.
Furthermore, clusters obtained by UPGMA, when a cutoff is at a distance of 0.43, paralleled those obtained by the Tocher optimization method (Table 3). Only two of the 38 genotypes presented some clustering divergence between methods. Genotype D-83-15 was clustered in group 1 by UPGMA and in group 7 by Tocher.
| Table 3: | Grouping of 38 wheat genotypes assessed from genetic distances obtained by microsatellite marker data, using UPGMA and the Tocher's methods |
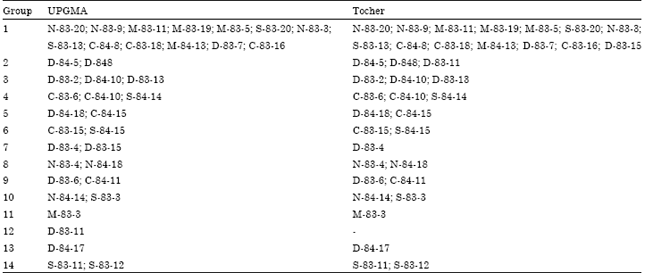 | |
Genotype D-83-11, clustered in group 2 by UPGMA, remained isolated in group 12 by Tocher. The other 36 genotypes were clustered similarly by both methods, thus indicating a high degree of clustering consistency.
The 14 clusters formed revealed high genetic diversity among the wheat cultivars. Group 1, consisting of 13 cultivars by the Tocher method and 14 by the UPGMA, contained genotypes from all origins. The three M-83 genotypes with the same genealogy (M-83-5,M-83-19 and M-83-11) were closely clustered. The two genotype from with the same pedigree (N-83-9 and N-83-20), were also clustered within this group (Fig. 2). The results obtained in this study revealed that there is significant genetic divergence among Iranian wheat genotypes, so that breeding programs can exploit expressive genetic variability by using only adapted germplasm. This observations can be set forth based on molecular data as much as genealogy. However, pedigree can not group genotypes by genetic similarity, whereas molecular data can do so.
Table 2 shows the frequencies of all alleles revealed in polyacrylamide gels. Since fragment size characterization is not completely reproducible in this system, a representative genotype was placed for each allele, so as to enable recognizing alleles identical to those observed in this study when using these same genotypes in future studies. This is important, especially in cases where it is necessary to define the genetic identity of a certain genotype. On knowing allele frequency, identity and exclusion probabilities can then be calculated (Schuster et al., 2006). Results obtained in this work show that a highly variable wheat germplasm has been adapted for usage in Iran. Therefore, it is possible to widely exploit this vast variability in segregating populations within breeding programs, by simply using only adapted genotypes. The results also provide information on the informativeness of those microsatellite loci and allelic frequencies that may be applied in evaluating still further the wheat germplasm used in Iran, as well as in protecting intellectual property.
These genetic profiles or fingerprints may be useful in the protection of genotypes for ensuring genetic purity and for generating further information to underpin breeding programs (Bertini et al., 2006). In addition and due to the allelic frequencies obtained in each locus, it is possible to acquire genetic identity and exclusion probabilities in cases involving the protection of intellectual-property (Schuster et al., 2006).
To date, no evaluation of Iranian wheat germplasm by using microsatellite markers has been undertaken. The markers applied here were found to be polymorphic, with a potential for germplasm characterization. With this in mind, these should be chosen according to their informativeness and measured by PIC magnitude values. On evaluating 23 Brazilian wheat cultivars, Bertan et al. (2006) arrived at similar results.
DISCUSSION
Results obtained with polymorphic primers in this study were consistent with those reported by other authors. Ahmed (2002) observed from two to eight alleles per locus, with a mean of 3.6, in 13 wheat genotypes of diverse origin analyzed with 43 SSR markers. Akkaya and Buyukunal-Bal (2004) obtained PIC values ranging from 0.36 to 0.87 with a mean of 0.68 when evaluating 19 SSR highly polymorphic loci in 11 wheat cultivars. We observed a mean of 3.26 alleles per locus within a range from two to five and for PIC values ranging from 0.20 to 0.79, a mean of 0.48. The microsatellite profiles, characterized with 23 pairs of polymorphic markers showed that the loci employed delineated all the 38 genotypes. Thus, these markers can be used to define the genetic profile of each genotype.
Several authors reported that microsatellites are more variable than most of other molecular markers that are useful tools studying the genetic diversity of germplasm. Plaschke et al. (1995) has used wheat microsatellite for the first time for studing the genetic diversity in closely related European bread wheat varieties. The authors obtained on an average 6.2 alleles per locus by using 23 markers. They also confirmed that a small number of markers were sufficient to distinguish closely related wheat genotypes and to select genotypes for higher genetic diversity. Prasad et al. (2000) assessed of the genetic diversity of 55 genotypes of wheat obtained from 29 countries and found the average number of alleles to be 7. On the other hand, Amer et al. (2001) detected genetic diversity in 15 Libyan wheat genotypes and found the average number of 4.5 alleles per locus. The authors suggested that a relatively small number of primers could be used to distinguish all genotypes and to estimate the genetic diversity. In recent years Huang et al. (2002) investigated 998 wheat accessions of bread wheat obtained from Gatersleben Gene Bank and reported 18.1 alleles per locus. The PIC values are informative to examine the extent of diversity. The PIC values in this study ranged from 0.65 to 0.88 with an average of 0.76. In earlier studies on wheat, the PIC values ranged from 0.21 to 0.94 (Roder et al., 1995; Plaschke et al., 1995; Prasad et al., 2000; Huang et al., 2002). Genetic Diversity in Hexaploid Wheat Genotypes obtained in the earlier studies on wheat may be attributed either to a small population (e.g., 18 genotypes used by Roder et al., 1995), to a set of closely related genotypes (Plaschke et al., 1995) or to a set of fewer microsattellite primers (e.g., 15 markers used by Roder et al. (1995).
The results of the present study were compared with those of Huang et al. (2002) and Alamerew et al. (2004). Huang et al. (2000) reported that all accessions that originated from the same geographic region did not cluster in the same group. The reason for this might be that similar genetic variation occurred independently in different geographic regions or that artificial transfer of an accession from one region to another resulted in a false determination of the geographic origin.
REFERENCES
- Ahmed, M., 2002. Assessment of genomic diversity among wheat genotypes as determined by simple sequence repeats. Genome, 45: 646-651.
PubMed - Akkaya, M.S. and E.B. Buyukunal-Bal, 2004. Assessment of genetic variation of bread wheat varieties using microsatellite markers. Euphytica, 35: 179-185.
CrossRefDirect Link - Alamerew, S., S. Chebotar, X. Huang, M. Roeder and A. Boerner, 2004. Genetic diversity in ethiopian hexaploid and tetraploid wheat germplasm assessed by microsatellite markers. Genet. Resour. Crop Evol., 51: 559-567.
CrossRefDirect Link - Anderson, J.A., G.A. Churchill, J.E. Autrique, S.D. Tanksley and M.E. Sorrells, 1993. Optimizing parental selection for genetic linkage maps. Genome, 36: 181-186.
CrossRefPubMedDirect Link - Bertini, C.H.C.M., I. Schuster, T. Sedyiama, E.G. Barros and M.A. Moreira, 2006. Characterization and genetic diversity analysis of cotton cultivar using microsatellite. Genet. Mol. Biol., 29: 321-329.
Direct Link - Huang, X.Q., S.L.K. Hsam, F.J. Zeller, G. Wenzel and V. Mohler, 2000. Molecular mapping of wheat powdery mildew resistance gene Pm24 and marker validation for molecular breeding. Theor. Applied Genet., 101: 407-414.
CrossRefDirect Link - Ijaz, S. and I.A. Khan, 2009. Molecular characterization of wheat germplasm using microsatellite markers. Genet. Mol. Res., 8: 809-815.
PubMed - McDonald, M.B., L.J. Elliot and P.M. Sweenery, 1994. DNA extraction from dry seeds for RAPD analyses in varietal identification studies. Seed Sci. Technol., 22: 171-176.
Direct Link - Plaschke, J., M.W. Ganal and M.S. Roder, 1995. Detection of genetic diversity in closely related bread wheat using microsatellite markers. Theor. Applied Genet., 91: 1001-1007.
CrossRefDirect Link - Prasad, M., R.K. Varshney, J.K. Roy, H.S. Balyan and P.K. Gupta, 2000. The use of microsatellites for detecting DNA polymorphism, genotype identification and genetic diversity in wheat. Theor. Applied Genet., 100: 584-592.
CrossRefDirect Link - Roder, M.S., J.S. Plaschke, U. Konig, A. Borner and M.E. Sorrells, 1995. Abundance, variability and chromosomal location of microsatellites in wheat. Mol. General Genet., 246: 327-333.
Direct Link - Roder, M.S., V. Korzun, K. Wendehake, J. Plaschke, M.H. Tixier, P. Leroy and M.W. Ganal, 1998. A microsatellite map of wheat. Genetics, 149: 2007-2023.
PubMedDirect Link - Fu, Y.B. and D.J. Somers, 2009. Genome-wide reduction of genetic diversity in wheat breeding. Crop Sci., 49: 161-168.
CrossRef - Quarrie, S.A., D. Dodig, S. Pekic, J. Kirby and B. Kobiljski, 2003. Prospects for marker-assisted selection of improved droughtresponses in wheat. Bulg. J. Plant Physiol., 2003: 83-95.
Direct Link