
B.Y. Kamaruzzaman
International Islamic University Malaysia, Jalan Sultan Ahmad Shah, Bandar Indera Mahkota, 25200 Kuantan, Pahang, Malaysia
LiveDNA: 60.4888
B. Akbar John
International Islamic University Malaysia, Jalan Sultan Ahmad Shah, Bandar Indera Mahkota, 25200 Kuantan, Pahang, Malaysia
K. Zaleha
International Islamic University Malaysia, Jalan Sultan Ahmad Shah, Bandar Indera Mahkota, 25200 Kuantan, Pahang, Malaysia
K.C.A. Jalal
International Islamic University Malaysia, Jalan Sultan Ahmad Shah, Bandar Indera Mahkota, 25200 Kuantan, Pahang, Malaysia
Asian Journal of Biotechnology
Year: 2011 | Volume: 3 | Issue: 3 | Page No.: 302-309







ABSTRACT
An effort to assess the utility of 650 bp Cytochrome C oxidase subunit I (DNA barcode) gene in delineating the members horseshoe crabs (Family: xiphosura) with closely related sister taxa was made. A total of 33 sequences were extracted from National Center for Biotechnological Information (NCBI) which include horseshoe crabs (Class: Merostomata), beetles (Class: Insecta), common crabs (Class: Melacostraca) and scorpion (Class: Arachnida) sequences. Constructed phylogram through comprehensive dry lab methodology using advanced software predictive tools showed that beetles are closely related with horseshoe crabs than common crabs (Class: Melacostraca). It was interesting to note that terrestrial Scorpion (Class: Arachnida) were distantly related to horseshoe crabs (Class: Merostomata). Phylogram and observed Genetic Distance (GD) data were also revealed that Limulus polyphemus was distantly related to all the other horseshoe crab species. Tachypleus gigas was closely related with Carcinoscorpius rotundicauda than with Tachypleus tridentatus. The observed mean Genetic Distance (GD) value was higher in 3rd codon position in all the selected group of organisms. Among the horseshoe crabs high GC content was observed in L. polyphemus (38.32%) and lowest was observed in T. tridentatus (32.35%). We conclude that COI sequencing (barcoding) could be used in identifying and delineating evolutionary relatedness with closely related species.
PDF Abstract XML References Citation
Received: October 05, 2010;
Accepted: November 19, 2010;
Published: February 07, 2011
How to cite this article
B.Y. Kamaruzzaman, B. Akbar John, K. Zaleha and K.C.A. Jalal, 2011. Molecular Phylogeny of Horseshoe Crab. Asian Journal of Biotechnology, 3: 302-309.
DOI: 10.3923/ajbkr.2011.302.309
URL: https://scialert.net/abstract/?doi=ajbkr.2011.302.309
DOI: 10.3923/ajbkr.2011.302.309
URL: https://scialert.net/abstract/?doi=ajbkr.2011.302.309
INTRODUCTION
Mitochondrial DNA (mtDNA) analysis has been employed in the evolutionary study of the animal species for more than 30 years (Brown et al., 1979; Avise and Walker, 1999). Its higher mutational rate and lower effective population size than the nuclear DNA make mtDNA a powerful tool to probe for evolutionary studies. This fact provoked a proposal to standardize DNA-based species identification by analyzing a uniform segment of the mitochondrial genome. A library of sequences from taxonomically verified voucher specimens could be built with this approach which could serve as DNA identifiers for species, in short, DNA barcodes (Herbert et al., 2003). For animals, 648 bp segment of the mitochondrial gene cytochrome C oxidase I (COI), which can be readily recovered from diverse species with a limited set of primers, was declared as a DNA barcode (Kevin et al., 2007). For this DNA barcoding approach to be effective, it must be possible to distinguish between intraspecific and interspecific mtDNA variation. The simplest test is whether the genetic distance within the species is lesser than those between species.
There are four extant species of horseshoe crabs, Tachypleus tridentatus, Tachypleus gigas, Carcinoscorpius rotundicauda and Limulus polyphemus (Pocock, 1902). The first three species inhabit the Southeast Asian coast and the last species the East Coast of North America.
These are known as living fossils, have maintained their morphology almost unchanged for the past 150 million years. The little morphological differentiation among horseshoe crab lineages has resulted in substantial controversy concerning the phylogenetic relationship among the extant species of horseshoe crabs, especially among the three species in the Indo-Pacific region. Earlier studies suggest that the three species constitute a phylogenetically irresolvable trichotomy (Xia, 2000). For elucidating their phylogenetic relationships, two proteins, coagulogen and hemocyanin, have been investigated (Shishikura et al., 1982; Srimal et al., 1985; Sugita, 1986). Miyazaki et al. (1989) first investigated tropomyosin which is one of the major structural proteins involved in many types of cells, to elucidate prevailing phylogenetic relationships among horseshoe crabs and his result suggested that L. polyphemus is phylogenetically differentiated far from the three Asian species. But the results were not those which were worth studying from the detailed phylogenetic viewpoint, that is, the patterns of the four species were equally different from each other. When the proteins were cleaved with chymotrypsin or trypsin, only smaller differences than those obtained with V8 protease were found (Miyazaki et al., 1989).
What remains unresolved is the phylogenetic relationship among the three Indo-Pacific species. T. gigas and T. tridentatus were grouped together on the basis of morphological traits (Fisher, 1984), but C. rotundicauda and T. tridentatus appear to be more closely related on the basis of amino acid sequence divergence of a fibrinopeptide like protein (Shishikura et al., 1982) and coagulogen (Srimal et al., 1985) and on interspecific hybridization studies (Sekiguchi and Sugita, 1980). In a phylogenetic study of the four species of horseshoe crabs employing two-dimensional electrophoresis of skeletal and cardiac muscles, the similarity index is the greatest between T. gigas and C. rotundicauda for cardiac muscles, but between T. tridentatus and T. gigas for skeletal muscles (Miyazaki et al., 1987). Phylogenetic analyses to find out the genetic relatedness among the four horseshoe crab species based on two partial mitochondrial genes, (16S ribosomal RNA (rRNA) and cytochrome oxidase subunit I (COI)) was also carried out (Xia, 2000). But comprehensive study on genetic relatedness among horseshoe crabs and its related sister taxa is scarce.
As, COI has been proposed as a barcode gene for most of the eukaryotes (Herbert et al., 2003). To enhance our understanding on the phylogeny of horseshoe crab and its genetic relatedness with other sister taxa, present study was conducted to find the phylogenetic cues in mitochondrial COI region and to find out evolutionary relationship of horseshoe crabs with closely related taxa.
MATERIALS AND METHODS
Sequence features: A total of 33 barcode sequences belong to horseshoe crab (12), insects including beetle (8), common crab (5) and scorpion (8) were extracted from National Center for Biotechnological Information (NCBI) via FASTA format during March 2010. Artemia franciscana (NC_001620.1) was shuffled among the sequences was clearly occupied separate branch in the phylogram as an out-group. Pair wise sequence alignment of nucleotide sequences were performed using ClustalX 2.0.6 (Thomson, 1997). The GC content of all 33 barcodes was estimated by BioEdit sequence alignment editor (Hall et al., 1999). MEGA 4.1 Beta3 (Tamura et al., 2007) was used to construct phylogenetic trees via Neighborhood joining method using Kimura 2-parameter and to calculate genetic distance of the given set of sequences in each codon position.
| Table 1: | Total number of organisms used to construct the phylogram and their respective accession ID with total sequence length (bp) is given. Percentage of GC content for each organism was calculated using Bioedit sequence alignment editor V.7.0.9.0 (Hall et al., 1999) |
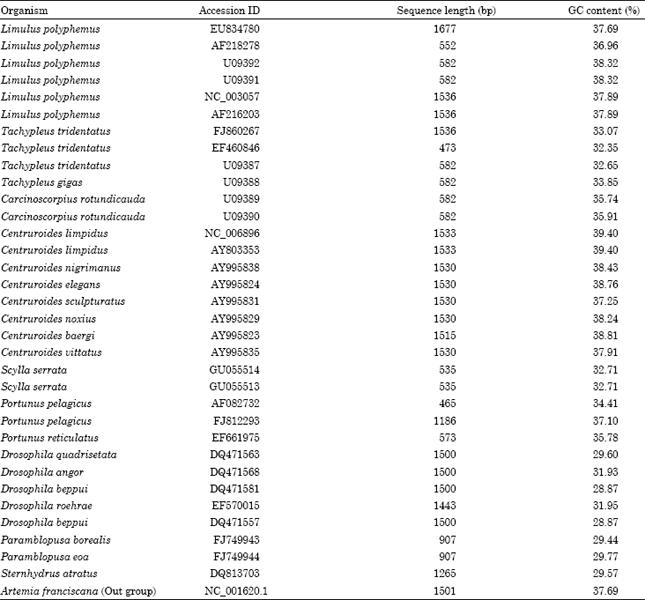 | |
Details of sequences used to generate the phylogram are given in Table 1.
RESULTS
Phylogenetic analysis: A phylogenetic tree was constructed to verify the efficiency of coxI gene in delineating closely related species of horseshoe crab and to check its evolutionary relationship with other groups of organisms which were proved to be closest relatives of xiphosurans. The constructed phylogram showed two distinct Clads (A and B). Artemia franciscana (NC_001620.1), used as an out group was clearly branched separately, signified the reliability of constructed phylogram. Almost all the internal branches showed high bootstrap value (>90).
Clad A includes horseshoe crabs, insects (beetles) and common crab species and all the scorpion species were distinctly clustered in clad B. As it was expected beetles (Class: Insecta) used in this study was perfectly arranged in clad A showing higher genetic similarity (lesser genetic distance) with horseshoe crab and common crabs were showing higher genetic distance with xiphosurans. Terrestrial scorpion species used in this analysis were clumped together in Clad B indicating their distant genetic relatedness with Horseshoe crab species. The phylogram also showed that L. polyphemus was genetically distinct from 3 other species of horseshoe crabs. Among the three Asian horseshoe crab species T. gigas showed closer genetic relatedness with C. rotundicauda than with T. tridentatus hence they occupied same internal nod in the phylogram.
Among the horseshoe crabs L. plyphemus had comparatively higher GC content than the other species of horseshoe crabs. Average GC content in L. polyphemus was 37.84% followed by C. rotundicauda 35.82%. T. gigas had higher average GC content (33.85%) than T. tridentatus (32.69%) but lower than the other two species.
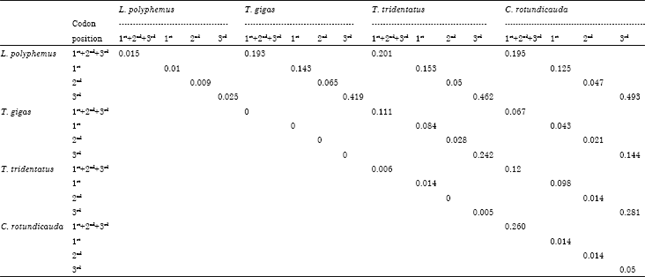
Mean genetic distance within L. polyphemus at 1st, 2nd and 3rd codon position were 0.01, 0.009 and 0.025, respectively. Likewise for T. tridentatus it was 0.014, 0 and 0.005 at 1st, 2nd and 3rd codon position, respectively. In case of C. rotundicauda the mean genetic distance values were 0.014, 0.014 and 0.05 at 1st, 2nd and 3rd codon position, respectively. Among the Asian horseshoe crab species T. gigas showed closer genetic relatedness (lower GD value) with C. rotundicauda with the value of 0.043, 0.021 and 0.144 at 1st, 2nd and 3rd codon position, respectively than with T. tridentatus with the values of 0.084, 0.028 and 0.242, respectively at 1st, 2nd and 3rd codon position. Calculated genetic distance data showed higher genetic distance in third codon position than its corresponding first and second codon positions (Table 2).
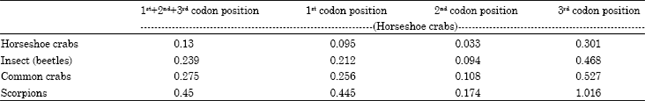
Mean GD value within the four species of horseshoe crabs was 0.095, 0.033 and 0.301 at 1st, 2nd and 3rd codon position respectively. The GD value between Insects (beetle) and horseshoe crabs were 0.201, 0.094 and 0.468 at 1st, 2nd and 3rd codon position respectively proved that horseshoe crabs are genetically more closer to insects followed by common crabs (GD values: 1st = 0.256; 2nd = 0.108 and 3rd = 0.527). Terrestrial scorpion species were showing higher mean GD value with horseshoe crabs (1st = 0.445; 2nd = 0.174 and 3rd = 1.016) clearly demarcated their distant genetic relatedness (Table 3). Figure 1 shows the results.
| Table 2: | Average genetic distance (GD) between four available species of horseshoe crabs observed in different codon positions |
 | |
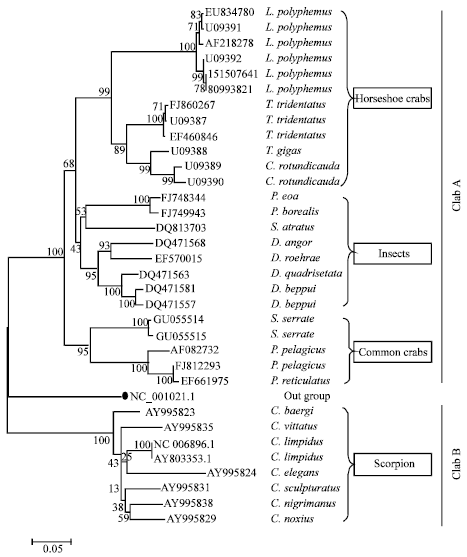 | |
| Fig. 1: | The evolutionary history was inferred using the Neighbor-Joining method (Saitou and Nei, 1987). The optimal tree with the sum of branch length = 1.67404499 is shown in figure. The percentage of replicate trees in which the associated taxa clustered together in the bootstrap test (500 replicates) is shown next to the branches (Felsenstein, 1985). The tree is drawn to scale, with branch lengths in the same units as those of the evolutionary distances used to infer the phylogenetic tree. The evolutionary distances were computed using the Kimura 2-parameter method (Kimura, 1980) and are in the units of the number of base substitutions per site. Codon positions included were 1st+2nd+3rd+Noncoding. All positions containing gaps and missing data were eliminated from the dataset (Complete deletion option). There were a total of 434 positions in the final dataset. Phylogenetic analyses were conducted in MEGA4 (Tamura et al., 2007) |
| Table 3: | Mean genetic distance (GD) values of different groups of organisms with reference to horseshoe crab at all the possible codon position indicating 3rd codon position shows higher GD value |
 | |
DISCUSSION
The constructed phylogram clearly indicated that horseshoe crabs are distantly related with scorpions at DNA level. Among the horseshoe crabs L. polyphemus showed distant genetic relatedness with other 3 species of Asian horseshoe crabs. This result was also well corresponded with previous finding by Miyazaki et al. (1987) where in two dimensional electrophoresis techniques clearly demarcated L. polyphemus from the three other species. T. gigas showed closer genetic relationship with C. rotundicauda than with T. tridentatus. This observation was also well coincide with the previous findings (Xia, 2000) where partial mitochondrial DNA sequence analysis revealed grater similarity index between T. gigas and C. rotundicauda than with T. tridentatus.
Insects were showing higher genetic similarity (lesser genetic distance) with xiphosurans (Horseshoe crabs) than the common crabs indicating that horseshoe crab might probably evolved from the ancient aquatic insects. Eurypterids (e.g., sea scorpions) have traditionally been regarded as close relatives of horseshoe Crabs; together forming a group called Merostomata. Subsequent studies placed eurypterids closer to the arachnids (e.g., spiders, terrestrial scorpions, mites and ticks) in a group called Metastomata (Pavlicek et al., 2008). There has also been a prevailing idea that eurypterids are closely related to terrestrial scorpions (Raz et al., 2009). The most recent study of relationships between arachnids and their relatives recognized Eurypterida, Xiphosura and Arachnida as three major groups, but was not able to resolve details between them (Shultz, 2007). Same result was reflected in the present study by separating scorpion species in a clad B indicating that horseshoe crabs have lesser genetic relatedness with terrestrial scorpions (Fig. 1).
Another interesting observation made from the calculated genetic distance data was higher genetic distance was observed in third codon position than its corresponding first and second codon positions. Similar observation was made by Ward et al. (2005) while barcoding of fishes from Australian waters. Simmons et al. (2006) also observed that greater phylogenetic signal is often found in parsimony-based analyses of third codon positions of protein-coding genes relative to their corresponding first and second codon positions, even for early-derived basal clades (Khan et al., 2010; Siemion and Przemyslaw, 1994). Average genetic distance among the different groups of test organisms used in this study showed higher GD value at 3rd codon position indicating that detailed study on 3rd codon position might reveal possible evolutionary information among the closely related groups of organisms.
CONCLUSION
The COI sequence in the phylogram constructed clearly clustered the selected species in individual group, proving the efficacy of COI gene in delineating the members of evolutionarily cryptic groups of organisms. Constructed phylogram and genetic distance data clearly proved that horseshoe crabs are more genetically related to insects (Class: Insecta) than with common crabs and are distantly related with terrestrial scorpions. But further studies need to be conducted to prove this concept by analyzing total mitochondrial DNA sequence. It is also evident from the present study that the greater phylogenetic signal is often found in third codon position relative to their corresponding first and second codon positions as reported by Ward et al. (2005) and Simmons et al. (2006).
ACKNOWLEDGMENTS
This research work was funded by IIUM internal endowment fund (Ref No. EDW B100960435). We wish to express our sincere gratitude to International Islamic University Malaysia (IIUM) and Institute of Oceanography and Maritime Study (INOCEM) for providing infrastructural facilities. One of us extent his thanks to Dr. Shahriza, University Malaysia Terengganu (UMT) for his valuable comments on this present work.
REFERENCES
- Avise, J.C. and D.E. Walker, 1999. Species realities and numbers in sexual vertebrates: Perspectives from an asexually transmitted genome. Proc. Natl. Acad. Sci. USA., 96: 992-995.
CrossRefDirect Link - Brown, W.M., M. Jr. George and A.C. Wilson, 1979. Rapid evolution of animal mitochondrial DNA. Proc. Natl. Acad. Sci. USA., 76: 1967-1971.
Direct Link - Felsenstein, J., 1985. Confidence limits on phylogenies: An approach using the bootstrap. Evolution, 39: 783-791.
CrossRefDirect Link - Hall, T.A., 1999. BioEdit: A user-friendly biological sequence alignment editor and analysis program for windows 95/98/NT. Nucleic Acid Symp. Ser., 41: 95-98.
Direct Link - Hebert, P.D.N., A. Cywinska, S.L. Ball and J.R. de Waard, 2003. Biological identification through DNA barcodes. Proc. Biol. Sci., 270: 313-321.
PubMedDirect Link - Miyazaki, J.I., T. Ishimoda-Takagi, K. Sekiguchi and T. Hirabayashi, 1989. Comparative study of horseshoe crab Tropomyosin. Comp. Biochem. Physiol., 93: 681-687.
CrossRef - Kerr, K.C.R., M.Y. Stoeckle, C.J. Dove, L.A. Weigt, C.M. Francis and P.D.N. Hebert, 2007. Comprehensive DNA barcode coverage of North American birds. Mol. Ecol. Notes, 7: 535-543.
CrossRefDirect Link - Khan, S.A., P.S. Lyla, B.A. John, C.P. Kuamr, S. Murugan and K.C.A. Jalal, 2010. DNA barcoding of Stolephorus indicus, Stolephorus commersonnii and Terapon jarbua of Parangipettai coastal waters. Biotechnology, 9: 373-377.
CrossRefDirect Link - Kimura, M., 1980. A simple method for estimating evolutionary rates of base substitutions through comparative studies of nucleotide sequences. J. Mol. Evol., 16: 111-120.
CrossRefDirect Link - Miyazaki, J.I., K. Sekiguchi and T. Hirabayashi, 1987. Application of an improved method of two-dimensional electrophoresis to the systematic study of horseshoe crabs. Biol. Bull., 172: 212-224.
Direct Link - Pavlicek, T., H.K. Mienis, S. Raz, V. Hassid, A. Rubenyan and E. Nevo, 2008. Gastropod biodiversity at the evolution canyon microsite, lower Nahal Oren, Mount Carmel, Israel. Biol. J. Linnean Soc., 93: 147-155.
CrossRef - Saitou, N. and M. Nei, 1987. The neighbor-joining method: A new method for reconstructing phylogenetic trees. Mol. Biol. Evol., 4: 406-425.
CrossRefPubMedDirect Link - Sekiguchi, K. and H. Sugita, 1980. Systematics and hybridization in the four living species of horseshoe crabs. Evolution, 34: 712-718.
Direct Link - Siemion, I.Z. and J.S. Przemyslaw, 1994. The informational context of the third base in amino acid codons. Biosystems, 33: 139-148.
CrossRef - Shishikura, F., S. Nakamura, K. Takahashi and K. Sekiguchi, 1982. Horseshoe crab phylogeny based on amino acid sequences of the fibrino-peptide-like peptide. J. Exp. Zool., 223: 89-91.
CrossRef - Shultz, J.W., 2007. A phylogenetic analysis of the arachnid orders based on morphological characters. Zool. J. Linnean Soc., 150: 221-265.
CrossRef - Simmons, M.P., L.B. Zhang, C.T. Webb and A. Reeves, 2006. How can third codon positions outperform first and second codon positions in phylogenetic inference: An empirical example from the seed plants. Syst. Biol., 55: 245-258.
CrossRef - Srimal, S., T. Miyata, S. Kawabata, T. Miyata and S. Iwanaga, 1985. The complete amino acid sequence of coagulogen isolated from southeast Asian horseshoe crab, Carcinoscorpius rotundicauda. J. Biochem., 98: 305-318.
Direct Link - Tamura, K., J. Dudley, M. Nei and S. Kumar, 2007. MEGA4: Molecular Evolutionary Genetics Analysis (MEGA) software version 4.0. Mol. Biol. Evol., 24: 1596-1599.
CrossRefPubMedDirect Link - Ward, R.D., T.S. Zemlak, B.H. Innes, P.R. Last and P.D. Hebert, 2005. DNA barcoding Australias fish species. Philos. Trans. Royal Soc. Lond. B Biol. Sci., 360: 1847-1857.
PubMed - Xia, X., 2000. Phylogenetic relationship among horseshoe crab species: Effect of substitution models on phylogenetic analyses. Syst. Biol., 49: 87-100.
Direct Link