
Dikayani
School of Life Sciences and Technology, Institute of Technology, Bandung, Indonesia
State of Islamic Unirversity Sunan Gunung Djati Bandung, Indonesia
Sri Nanan B. Widiyanto
School of Life Sciences and Technology, Institute of Technology, Bandung, Indonesia
Erly Marwani
School of Life Sciences and Technology, Institute of Technology, Bandung, Indonesia
Rina Ratnasih
School of Life Sciences and Technology, Institute of Technology, Bandung, Indonesia
Asian Journal of Biochemistry
Year: 2015 | Volume: 10 | Issue: 6 | Page No.: 312-317







ABSTRACT
Environment has influenced the growth and development of agricultural crops including banana (Musa acuminata L.). Environmental stresses are caused by biotic factors such as fungi, bacteria and herbivore and abiotic factors such as temperature, water, light and salinity. Salinity stresses cause a decrease in the production of banana, because they affect the growth of the plant in the changing balance of Na+ and Cl‾ ions and changes in the plant’s primary metabolites including fatty acid compounds. The purpose of the research was to identify the fatty acid compounds. The research method was conducted in 2 stages: phase (1) Developing in vitro culture of banana shoots with NaCl treatment of 0, 50, 100, 150 and 200 mM concentrations, (2) Gas Chromatography Mass Spectrometry (GCMS) analysis on the roots of in vitro banana planlets. The results showed that the fatty acid compounds involved in the protection against salt stress. The fatty acid the highest area compound in 0 mM treatment 9 Octadecanoic acid methyl ester were (14:36%), 50 mM treatment 9-octadecanoic acid methyl ester 13.13% and 100 mM treatment Tridecanoic acid 11.8%. In 150 mM treatment 9,12 octadecanoic acid 31.03%, whereas, the highest area in 200 mM treatment 9,12 octadecanoic acid 36.49%.
PDF Abstract XML References Citation
Received: August 21, 2015;
Accepted: October 16, 2015;
Published: October 29, 2015
How to cite this article
Dikayani, Sri Nanan B. Widiyanto, Erly Marwani and Rina Ratnasih, 2015. Composition of Fatty Acids due to Salinity in the Root of in vitro Culture of Musa acuminata L. Planlets of Barangan Cultivars. Asian Journal of Biochemistry, 10: 312-317.
DOI: 10.3923/ajb.2015.312.317
URL: https://scialert.net/abstract/?doi=ajb.2015.312.317
DOI: 10.3923/ajb.2015.312.317
URL: https://scialert.net/abstract/?doi=ajb.2015.312.317
INTRODUCTION
Stresses that often occur in the environment, such as drought, salinity and extreme temperature, are three abiotic stresses that are mostly found in agricultural lands; the three stresses are the cause of the decline in agricultural production (Rodriguez et al., 2005). Agricultural crops experiencing stresses caused by drought, extreme temperature and salinity can undergo osmotic stresses, causing loss of turgor pressure, cell membrane damage, disruption of protein synthesis and enzyme activity (Hasegawa et al., 2000; Kosova et al., 2013). In addition, abiotic stresses are caused by chemicals such as Reactive Oxygen Species (ROS), methyl jasmonat and other chemicals such as salinity (Do Nascimento and Fett-Neto, 2010).
Salinity naturally develops from a variety of processes, including those related to natural geology, hydrology and pedology. Climatic factors and water management may have something to do with salinity. Evapotranspiration has an important role in the pedogenesis of saline and sodic soil in arid and semi-arid lands (Allakhverdiev et al., 2000; Koca et al., 2007; Yadav et al., 2011). Under salinity stress, plants form active metabolite compounds to protect them from the environmental stresses.
Primary and secondary metabolites in banana plants are natural products, having their composition in protein molecules. Primary metabolites such as carbohydrates, proteins and fats are needed in a plant’s growth and development while secondary metabolites are compounds that are not essential for the plant’s growth and development but are often involved in the environmental adaptation to biotic and abiotic stresses (Do Nascimento and Fett-Neto, 2010). Salinity is generally related with changes in plant lipid metabolism such as fatty acid (Bybordi, 2011; Ashraf and Harris, 2004). The purpose of the research conducted was to determine the chemical compounds of fatty acids formed at the roots of the banana planlets with concentrations of 0, 100, 150 and 200 mM.
MATERIALS AND METHODS
Materials: The explants used were the shoots of banana planlet of PT Multi Agro Kultura Pamulang Tangerang. The basic media used were that of Murashige and Skoogs (MS), with Benzylaminopurin (BAP) 1.5 ppm and NaCl as growth regulators.
Methods: There were two stages performed, which are: (1) development of the shoot culture of banana planlet with the treatment of concentrations of NaCl 0, 50, 100, 150 and 200 mM, (2) analysis of active compounds using Gas Chromatography Mass Spectrometry (GCMS) in the roots of banana planlet with the concentrations of 0 mM NaCl, 50, 100, 150 and 200 mM. This study was conducted from January, 2013 until June, 2013 in laboratory Molecular Cell Biology School of Life Science and Technology, ITB, Indonesia.
Procedure for the making of in vitro banana culture: The explants of banana shoots were cultured in vitro in the Murashige and Skoog (MS) media of 4.4 g L–1, sucrose 30 g, gel 8 g and an addition of growth regulator Benzylaminnopurin (BAP) at a concentration of 1.5 ppm. The NaCl with the concentration of 0, 50, 100, 150 and 200 mM was added into the treatment media. In vitro culture was made in a sterile state, the medium was sterilized using an autoclave.
GAS CHROMATOGRAPHY MASS SPECTROMETRY ANALYSIS PROCEDURE
Extraction and determination of fatty acid compounds: Extraction was performed on 1 g sample of roots with the concentrations of 0 mM NaCl, 100 mM and then made finer by an addition of methanol. The result of the extraction was incubated in an ultrasonic bath for 1 h and then filtered using filter paper (Babu et al., 2012).
Gas chromatography mass spectrometry analysis: The extract was injected into the GCMS system of Fisons GC 8000, GC model 8060 paired with MS Fisons MD 800 in EI (Electron Impact), with an energy electron of 70 eV and masses ranging from 25-700 MLZ. The capillarity column was that of low-bleed CP-Sil 8 CB-MS (30 m×0.32 mm, i.d) of 0.25 um of film thickness. An injector was set at 280°C and a detector at 290°C and GC in splitless at 1 min. Temperature was set between 70-135°C with 2°C min–1 for10 min, between 135-220°C with 4°C min–1 for 10 min, between 220-270°C with 3.5°C min–1 for 20 min. After a run of 10 min at 70°C for the next injection, the average speed was 1.9 mL min–1. Identification of compounds was compared to the retention time with the right chemical compound and spectral data obtained from Wiley and NIST libraries. Each determination was duplicated (Proestos et al., 2006).
RESULTS AND DISCUSSION
The result of GCMS analysis showed several fatty acid compounds depicted in Figures, which are: with 0 mM treatment: 9 octadecanoic acid methyl ester, 11 octadecanoic acid methyl ester, oleic acid and the highest is 9-octadecanoic acid 14.36% (Fig. 1a) while with 50 mM NaCl treatment Pentadecanoic acid 14 methyl, methyl ester, Tritanopic acid, 9 octadecanoic acid methyl ester 13.13%. About 100 mM NaCl treatment: 9 octadecanoic acid methyl ester, 9-octadecanoic acid D ester and the highest compound is 9-octadecanoic acid methyl ester 13.13% (Fig. 1b). Concentrate NaCl 100 mM treatment hexadecanoic acid, tridecanoic acid, 9-octadecanoic acid and the highest area was tridecanoic acid 11.8% (Fig. 1c). In concentrate NaCl 150 mM 9-octadecanoic acid 2,3 dihydromethyl methyl ester, 9-octadecanoic acid methyl ester, 9,12 octadecanoic acid, oleic acid. The highest compound was 9,12 octadecanoic acid 31.03% (Fig. 1d). In 200 mM treatment obtained oleic acid, hexadecanoic acid methyl ester, 9,12 octadecanoic acid, 9-octadecanoic acid methyl ester, 9,12 octadecanoic acid, 9-octadecanoic acid, 2,3 dihydroxy methyl ester. The highest area compound 9,12 octadecanoic acid 36.49% (Fig. 1e).
Fatty acid compounds experienced quantity changes, such as 9 octadecanoic acid methyl ester which decreased 14:36% at 0 mM treatment and 0.97% at 100 mM treatment; the presence of other types of fatty acids at 100 mM treatment was also found, such as tridecanoic acid and hexadecanoic acid methyl ester; 150 mM treatment oleic acid 2.99%. Concentrate NaCl 200 mM treatment 9 octadecanoic acid methyl ester 10.86 and 3.51%. Many studies about fatty acid or essential oil of seeds of peanut, Papaver somniferum L., Satureja thymbra and S. cuneifolia (Ozcan and Seven, 2003; Schulz et al., 2005; Erinc et al., 2009; Goren et al., 2003) and others research in Rosa canina L., Poppy seed (Ozcan, 2002; Bozan and Temelli, 2003).
Salinity affects plants via osmotic effects, effects of specific ions and oxidative. Osmotic effects may reduce the induction of salt in water potentials. Salinity causes reduction of ions K+ and Ca2+, hence reducing the level of Na+ and Cl‾, also, it can induce cellular accumulation and damage active oxygen species. Active oxygen species can damage membrane lipids, proteins and nucleic acids (Pitman and Lauchli, 2002; Mittler, 2002).
Primary metabolism in plants produce primary metabolites consisting of proteins, carbohydrates and fats which are essential for the growth and development of plants including bananas. Primary metabolite synthesis is closely related to environmental factors, both biotic and abiotic. Abiotic factors consist of, among others, temperature, light, water and salinity; thus, when the environmental factors are disturbed, it may cause stresses on the plants, one of which is salinity stress. It is not only related to Na+ cellular homeostasis but also involves the presence of a chemical compound, i.e., changes in the level and composition of fatty acids or glycerol lipid in the adaptation to salinity stresses (Zhang and Blumwald, 2001).
Salt tolerance is not only related to the cellular homeostasis of Na+ ions but also involves the adaptation of secondary metabolite effects on the effects of salinity such as oxidative damages and changes in the level and composition of fatty acids of glycerol lipid in the roots and leaves (Yu et al., 1998; Wu et al., 1998). In addition to fatty acids involved in defense salinity stresses (Aghaleh and Nikman, 2009).
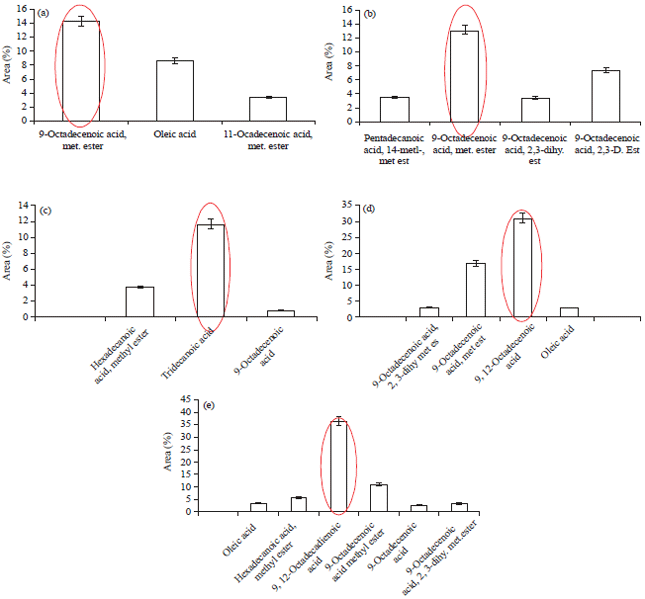 | |
| Fig. 1(a-e): | GCMS analysis of NaCl (a) 0 mM, (b) 50 mM, (c) 100 mM, (d) 150 mM and (e) 200 mM on the roots |
It is evident from the results of the analysis which showed that there were changes in the quantity of 9-octadecenoic acid methyl ester; a decrease of 14:36% from treatments 0 mM. There was also the presence of other types of fatty acids, formed at the treatment of 100 mM, i.e. Tridecanoic acid and Hexadecanoic acid methyl ester.
CONCLUSIONS
The fatty acids involved against salinity stressed and protected plant from environmental factor limiting. Salinity effected compound composition, each concentration of NaCl fatty acid causing komposisi difference. The results of gas chromatography mass spectrometry analysis on the roots of in vitro banana planlets due to the influence of salinity with various concentrations of fatty acid biosynthesis showed that fatty acid compounds were contained in the control, the highest is 9-octadecanoic acid methyl ester (14:36%). In 50 mM treatment obtained the highest area is 9-octadecanoic acid methyl ester 13.13% and 100 mM NaCl treatment tridecanoic acid 11.8%. In 150 mM treatment the highest area is 9,12-octadecanoic acid 31.03%, whereas at the 200 mM treatment 9,12-octadecanoic acid 36.49%.
ACKNOWLEDGMENT
The financial support of this research was provided from BPPS DIKTI. We thank to School Life of Science and Technology ITB for the facilities. We thank to State Islamic University Sunan Gunung Djati Bandung for support in the PhD Programme.
REFERENCES
- Aghaleh, M. and V. Niknam, 2009. Effect of salinity on some physiological and biochemical parameters in explants of two cultivars of soybean (Glyicine max L.). J. Phytol., 1: 86-94.
Direct Link - Allakhverdiev, S.I., A. Sakamoto, Y. Nishiyama, M. Inaba and N. Murata, 2000. Ionic and osmotic effects of NaCl-induced inactivation of photosystems I and II in Synechococcus sp. Plant Physiol., 123: 1047-1056.
Direct Link - Ashraf, M. and P.J.C. Harris, 2004. Potential biochemical indicators of salinity tolerance in plants. Plant Sci., 166: 3-16.
CrossRefDirect Link - Babu, M.A., M.A. Suriyakala and K.M. Gothandam, 2012. Varietal impact on phytochemical contents and antioxidant properties of Musa acuminata (Banana). J. Pharmaceut. Sci. Res., 4: 1950-1955.
Direct Link - Bozan, B. and F. Temelli, 2003. Extraction of poppy seed oil using supercritical CO2. J. Food Sci., 68: 422-426.
CrossRefDirect Link - Bybordi, A., 2011. Effects of NaCl salinity levels on lipids and proteins of canola (Brassica napus L.) cultivars. Romanian Agric. Res., 28: 197-206.
Direct Link - Erinc, H., A. Tekin and M.M. Ozcan, 2009. Determination of fatty acid, tocopherol and phytosterol contents of the oils of various poppy (Papaver somniferum L.) seeds. Grasas Aceites, 60: 375-381.
Direct Link - Goren, A.C., G. Bilsel, M. Altun and F. Satil, 2003. Fatty acid composition of seeds of Satureja thymbra and S. cuneifolia. Zeitschrift Naturforschung C, 58: 502-504.
CrossRefDirect Link - Hasegawa, P.M., R.A. Bressan, J.K. Zhu and H.J. Bohnert, 2000. Plant cellular and molecular responses to high salinity. Annu. Rev. Plant Physiol. Plant Mol. Biol., 51: 463-499.
CrossRefDirect Link - Koca, H., M. Bor, F. Ozdemir and I. Turkan, 2007. The effect of salt stress on lipid peroxidation, antioxidative enzymes and proline content of sesame cultivars. Environ. Exp. Bot., 60: 344-351.
CrossRefDirect Link - Kosova, K., I.T. Prasil and P. Vitamvas, 2013. Protein contribution to plant salinity response and tolerance acquisition. Int. J. Mol. Sci., 14: 6757-6789.
CrossRefDirect Link - Mittler, R., 2002. Oxidative stress, antioxidants and stress tolerance. Trends Plant Sci., 7: 405-410.
CrossRefPubMedDirect Link - Ozcan, M., 2002. Nutrient composition of rose (Rosa canina L.) seed and oils. J. Med. Food, 5: 137-140.
CrossRefDirect Link - Ozcan, M. and S. Seven, 2003. Physical and chemical analysis and fatty acid composition of peanut, peanut oil and peanut butter from COM and NC-7 cultivars. Grasas Aceites, 54: 12-18.
Direct Link - Proestos, C., D. Sereli and M. Komaitis, 2006. Determination of phenolic compounds in aromatic plants by RP-HPLC and GC-MS. Food Chem., 95: 44-52.
CrossRefDirect Link - Rodriguez, P., A. Torrecillas, M.A. Morales, M.F. Ortuno and M.J. Sanchez-Blanco, 2005. Effects of NaCl salinity and water stress on growth and leaf water relations of Asteriscus maritimus plants. Environ. Exp. Bot., 53: 113-123.
CrossRefDirect Link - Schulz, H., G. Ozkan, M. Baranska, H. Kruger and M. Ozcan, 2005. Characterisation of essential oil plants from Turkey by IR and Raman spectroscopy. Vibrat. Spectrosc., 39: 249-256.
CrossRefDirect Link - Yadav, S., M. Irfan, A. Ahmad and S. Hayat, 2011. Causes of salinity and plant manifestations to salt stress: A review. J. Environ. Biol., 32: 667-685.
PubMedDirect Link - Yu, B., H. Gong and Y. Liu, 1998. Effects of calcium on lipid composition and function of plasma membrane and tonoplast vesicles isolated from roots of barley seedlings under salt stress. J. Plant Nutr., 21: 1589-1600.
CrossRefDirect Link - Zhang, H.X. and E. Blumwald, 2001. Transgenic salt-tolerant tomato plants accumulate salt in foliage but not in fruit. Nat. Biotechnol., 19: 765-768.
CrossRefPubMedDirect Link - Wu, J., D.M. Seliskar and J.R. Gallagher, 1998. Stress tolerance in the marsh plant Spartina patens: Impact of NaCl on growth and root plasma membrane lipid composition. Physiologia Plantarum, 102: 307-317.
CrossRefDirect Link
Dekiansyah Reply
This paper give information a lot of compound in Musa acuminata, usefull for science. Thank you