
Samy Ali Hussein Aziza
Department of Biochemistry, Faculty of Veterinary Medicine Moshtohor, Benha University, Egypt
Mamdouh El-Haggar
Department of Biochemistry, Faculty of Veterinary Medicine Moshtohor, Benha University, Egypt
Omayma Ahmed Abo-Zaid
Department of Biochemistry, Faculty of Veterinary Medicine Moshtohor, Benha University, Egypt
Mohammed Ragaa Hassanien
Department of Biochemistry, Faculty of Veterinary Medicine Moshtohor, Benha University, Egypt
Ragab El-Shawarby
Department of Forensic Medicine and Toxicology, Faculty of Veterinary Medicine Moshtohor, Benha University, Egypt
Asian Journal of Biochemistry
Year: 2014 | Volume: 9 | Issue: 1 | Page No.: 1-15







ABSTRACT
Diabetic peripheral neuropathy is the most common complication of long-standing diabetes mellitus. The objective of this study was to investigate the fatty acids composition abnormalities of sciatic nerve in Streptozotocin (STZ)-induced Diabetic Neuropathy (DNP) in male rats. Six groups of 20 rats were studied. Group I (control group): Received no drugs, Group II (diabetic group) A single dose of Streptozotocin (STZ) (50 mg kg-1 i.p.) was used for the induction of diabetes in rats, Group III (normal α-lipoic acid-treated group), Group IV (diabetic alpha-lipoic acid-treated group), Group V (diabetic insulin-treated group), Group VI (diabetic alpha-lipoic acid and insulin-treated group). Eight weeks after diabetes induction therapeutic treatment with alpha-lipoic acid (54 mg kg-1 b.wt. i.p daily) and insulin (2 U s.c daily) were given either alone or in combination and continued for six weeks. Equivalent volumes of saline were given subcutaneously to the rats in the other diabetic and non diabetic control groups. Blood samples and sciatic nerve tissues were collected from all animal groups two times at 4 and 6 weeks from the onset of treatment which begin after eight weeks of Diabetic Neuropathy (DNP) induction for determination of serum glucose and sciatic nerve triacylglycerols, non Esterified Fatty Acids (NEFA) and membrane fatty acids composition of sciatic nerve. The obtained results revealed that, a significant increase in serum glucose and sciatic nerve NEFA concentrations with marked decrease in triacylglycerols and fatty acids composition percentage including palmitic acid, palmitoleic acid, oleic acid and arachidic acid were observed in sciatic nerve of STZ-induced DNP in rats. Administration of α-lipoic acid with insulin significantly lowered serum glucose and sciatic nerve NEFA concentrations as well as several fatty acids contents including myristic, palmitic, palmitoleic, oleic, cis-vaccenic, linoleic, alpha linolenic, arachidic and godoleic acid with increase in margaric, stearic and eicosadienoic acid percentage in addition to triacylglycerols level in sciatic nerve. The results suggest that, treatment with α-lipoic acid combined with insulin administrations could ameliorate the altered myelin of sciatic nerve lipids induced by diabetes and attenuate the changed fatty acid composition membrane in sciatic nerve and improved the metabolic control of diabetic neuropathy. The results indicated that the beneficial effect of α-lipoic acid treatment in diabetic neuropathy.
PDF Abstract XML References Citation
Received: October 15, 2013;
Accepted: November 01, 2013;
Published: March 04, 2014
How to cite this article
Samy Ali Hussein Aziza, Mamdouh El-Haggar, Omayma Ahmed Abo-Zaid, Mohammed Ragaa Hassanien and Ragab El-Shawarby, 2014. Fatty Acids Composition Abnormalities of Rats Sciatic Nerve in Streptozotocin-induced
Diabetic Neuropathy. Asian Journal of Biochemistry, 9: 1-15.
DOI: 10.3923/ajb.2014.1.15
URL: https://scialert.net/abstract/?doi=ajb.2014.1.15
DOI: 10.3923/ajb.2014.1.15
URL: https://scialert.net/abstract/?doi=ajb.2014.1.15
INTRODUCTION
Diabetes is a syndrome which is characterized by hyperglycemia, lipoprotein abnormalities and raise the proportion of basic metabolism and a defect in enzymes of high oxidative stress resulting from damage to the beta cells in the pancreas (Sharma et al., 2010). Diabetes is a serious chronic diseases worldwide and is caused by defects in insulin production, insulin secretion and insulin signals (Skyler, 2007). The causes are many different types of diabetes and that a complex interaction between genetics and environmental factors (Chitra et al., 2010). Type I diabetes is the consequence of an autoimmune-mediated destruction of pancreatic β-cells, leading to insulin deficiency. Patients require insulin treatment for survival (Malaisse, 1983). While, Type II diabetes mellitus developed by metabolic abnormalities such as impaired insulin secretion, increased hepatic glucose production and decreased insulin-stimulate of glucose uptake in peripheral (Kakadiya et al., 2010). In cell culture systems triacylglycerols mediate the uptake and transfer of fatty acids to membrane phospholipids (Blank et al., 1993) have demonstrated that over expressed mitochondria glycerol-3-phosphate acyltransferase directs incorporation of exogenous fatty acid into triacylgycerols rather than phospholipids (Igal et al., 2001).
The normal physiological functions of neuronal membranes largely dependent on the structure and although many factors can affect the membrane volatility index and the main one is the composition of membrane lipids: Cholesterol reduces membrane fluidity and PUFAs increase it (Yehuda et al., 2002). There are many reports that the oxidative stress caused by diabetes plays an important role in the development and progression of diabetic neuropathy (Deferrari et al., 2003). This is one of the commonest long-term complications of diabetes mellitus and leads to much of the morbidity and mortality, resulting in a huge economic burden for diabetes care (Das, 1994). This is probably a common fat nerve with myelin lipids and oxidative damage to myelin can reasonably contribute to neurological disorders diabetic neuropathy(Cunha et al., 2008).
Diabetic peripheral neuropathy is the majority complication of diabetes has long been leading in many cases, are in satisfactory condition clinically significant, such as pain and leg ulcers and amputation happen (Said, 2007). It is believed that, a large number of neural mechanisms of anatomical, physiological and neurological neurochemical to promote the development and maintenance of diabetic neuropathic pain (Edwards et al., 2008). Oxidative stress incorporation promote the formation of free radicals and/or antioxidant defenses in the pathogenesis of experimental diabetic neuropathy defect (Vincent et al., 2004). The protective effects of antioxidants administered externally studied extensively in animal models in recent years, thus providing insight into the relationship between free radicals, diabetes and its complications (Maritim et al., 2003). Alpha-lipoic acid-has potent antioxidant activity and is used clinically to treat diabetic neuropathy. In addition, α-lipoic acid-improve insulin stimulated glucose disposal and improves microcirculation and reduces the symptoms of peripheral neuropathy, possibly attenuated oxidative stress (Vessal et al., 2003). Also, α-lipoic acid appears to have a potent therapeutic role in addition to its role in management of diabetic neuropathy in protection of diabetic complications due to oxidative stress (El-Nabarawy et al., 2010).
Accordingly, this study was performed to investigate the fatty acids composition in the sciatic nerve of STZ-diabetic rats. Moreover, the effect of treatment with α-lipoic acid, insulin and their combination on serum glucose and triacylglycerols, non Esterified Fatty Acids (NEFA) and membrane fatty acids composition abnormalities of sciatic nerve in Streptozotocin (STZ)-induced diabetic neuropathy in rats were also investigated.
MATERIALS AND METHODS
Experimental animals: This study was carried out on 120 white male albino rats, 12-16 weeks old and weighted 220-250 g. Rats were housed in separated metal cages and kept at constant environmental and nutritional conditions throughout the period of experiment. The animals were fed on constant ration and water was supplied ad libitum. All animals were acclimatized for minimum period of two weeks prior to the beginning of study.
Drugs used
Alpha-lipoic acid (Thiotacid)R: Alpha-lipoic acid (Thioctic acid)® manufactured by EVA pharma for pharmaceuticals and Medical Apliances, Egypt.
Human insulin (HumulinR U-100): Humulin R manufactured by LILLY Egypt, under License from ELI LILLY USA.
Diabetes induction: Rats were fasted for 18 h and allowed free access of water. The experimental induction of diabetes in male rats was induced by a single intraperetinoel (i.p) injection of 50 mg kg-1 body weight of Streptozotocin (STZ) freshly dissolved in citrate buffer, pH 4.5. A week later, STZ-treated rats were fasted for 12 h and blood samples were collected from the orbital venous sinus for glucose determination. Only those rats in diabetic group with blood glucose levels higher than 250 mg dL-1 were considered diabetic (Ramanathan et al., 1999).
Eight weeks after diabetes induction therapeutic treatment with alpha-lipoic acid (54 mg kg-1 b.wt. i.p. daily) and insulin (2 U s.c daily) were given either alone or in combination and continued for six weeks. Equivalent volumes of saline were given subcutaneously to the rats in the other diabetic and non diabetic control groups. Diabetic neuropathy in rats were develop within 8 weeks after induction of diabetes (Kumar et al., 2005).
Animal grouping: After eight weeks of diabetes induction indicating development of diabetic neuropathy all rats were randomly divided into six main equal groups, 20 animal each, placed in individual cages and classified as follow: Group I (control normal group): Received no drugs, served as control for all experimental groups. Group II (control diabetic group): Received equivalent volumes of saline were given subcutaneously and served as STZ-induced diabetic group. Group III (normal alpha-lipoic acid-treated group): Received alpha-lipoic acid at a doselevel of (54 mg kg-1 b.wt. i.p. daily). Group IV (diabetic alpha-Lipoic acid-treated group): Received alpha-lipoic acid at a dose level of (54 mg kg-1 body weight i.p daily) for six weeks (Gruzman et al., 2004). Group V (diabetic insulin-treated group): Received subcutaneous injection of insulin at a dose level of 2 U each morning for six weeks (Izbeki et al., 2008). Group VI (diabetic alpha-lipoic acid with insulin-treated group): Received alpha-lipoic acid at a dose level of (54 mg kg-1 b.wt. i.p. daily) and insulin at dose of 2 U injected subcutaneously each morning for six weeks.
Sampling: Random blood samples and sciatic nerve specimence were collected from all animal groups (control and experimental group). Two times at 4 and 6 weeks, from the onset of treatment, after eight weeks of diabetes induction.
| • | Blood samples: Blood samples for serum separation were collected after over night fasting by ocular vein puncture at the end of each experimental period and serum was separated by centrifugation at 2500 rpm for 15 min. The clean, clear serum was proceed directly for glucose determination |
| • | Sciatic nerve samples: Sciatic nerves from the spin to the peroneal bifurcation were dissected, rinsed in ice-cold saline solution and frozen in liquid nitrogen after removal of adherent tissue. Samples were kept at -80°C in liquid nitrogen until use |
Lipid extraction in sciatic nerve were performed according to the method described by Peuchant et al. (1989). On the day of the homogenate preparation sciatic nerve segments were measured, weighed and cut into small pieces and then homogenized with bout 5 mL of isopropanol added to each tube and the tubes were shaken vigorously. After that sufficient amount of anhydrous sodium sulfate was then added to each tube to remove the water. The mixture was vortexed for 2 min and then filtered or centrifuged at 3000 rpm for 10 min. After centrifugation, aliquots were analyzed for the determination of triacylglycerols, non Esterified Fatty Acids (NEFA) concentrations and fatty acids composition.
Biochemical analysis: Seurm glucose, in addition to sciatic nerve triacylglycerols and Non Esterified Fatty Acids (NEFA) concentrations were analyzed colorimetrically according to the methods described by Trinder (1969), Fossati and Prencipe (1982) and Schuster and Pilz (1979), respectively. Fatty acids composition percentage in sciatic nerve were performed according to the method described by Ackman (1991).
Statistical analysis: The obtained data were statistically analyzed by one-way analysis of variance (ANOVA) followed by the Duncan multiple test. All analyses were performed using the statistical package for social science (SPSS 13.0 software 2009) (Field, 2009). Values of p<0.05 were considered to be significant.
RESULTS AND DISCUSSION
Diabetic peripheral neuropathy is the most common complication of diabetes has long been leading in many cases, are in satisfactory condition clinically significant, such as pain, foot ulcers and amputation (Said, 2007). It is believed that a large number of neural mechanisms of anatomical, physiological and neurological neurochemical to promote the development and maintenance of diabetic neuropathic pain (Edwards et al., 2008). Hyperglycemia clearly plays a key role in the development and progression of diabetic neuropathy as well as the other microvascular complications of diabetes. In addition, high blood sugar levels is an important factor in chronic neuropathy in a hurry (Tomlinson and Gardiner, 2008). However, you can start a pre-diabetic neuropathy, people and shows moderate weakness in the metabolism of glucose but not high blood sugar chronic public (Smith and Singleton, 2008).
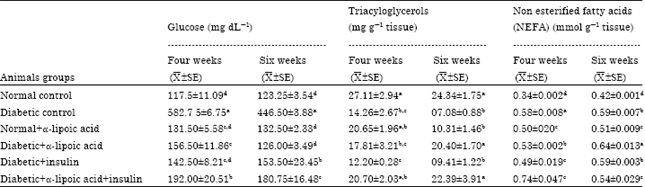
The factors like oxidative stress, Advanced Glycation Endproducts (AGE) formation, lipid peroxidation which results from diabetic condition can stimulate inflammatory process (Negi et al., 2010). The recorded data demonstrated in (Table 1) revealed that, a significant increase in serum glucose level and sciatic Nerve non Esterified Fatty Acids (NEFA) with marked decrease in triacylglycerols concentrations were observed in sciatic nerve in Streptozotocin-induced diabetic neuropathy in male rats allover the period of the experiment.
The increase in serum glucose level in STZ-induced diabetic rats are nearly similar to those reported by Sayyed et al. (2006) who observed that, STZ-induced diabetic rats showed approximately five-fold increase in the blood glucose levels after STZ administration. Dias et al. (2005) reported that, the plasma glucose concentration was significantly increased in diabetic rats.
| Table 1: | Effects of treatment with alpha-lipoic acid, insulin and their combination on serum glucose, triacyloglycerols and non esterified fatty acids of sciatic nerve in Streptozotocin-induced diabetic neuropathy in male rats |
 | |
| Data are represented as | |
The developed hyperglycemia have been attributed to the specific toxic effects of STZ uptake through glucose transporter-2 (GLUT-2), these toxic effects lead to end organ damage through activation of the aldose reductasc pathway leading to toxic accumulation of sorbitol in nervous system (Greene et al., 1999), increased diacyl glycerol synthesis with consequent activation of Protein Kinase C isoform (PKC) in vascular tissue, initiating diabetic complications (Craven et al., 1995) and increased oxidative stress with subsequent alterations in celluar redox balance (Williamson et al., 1993). Treatment with α-lipoic acid, insulin and their combination significantly reduced elevated serum glucose level in Streptozotocin (STZ)-induced diabetic neuropathy in rats allover the periods of the experiments. Vessal et al. (2003) reported that, oral administration of α-LA has shown hypoglycemic effects against STZ-induced diabetes in rats.
These effects can be attributed to the antioxidant supplements to reduce the concentration of glucose in the blood and strengthen the restoration of pancreatic islets and thus increase insulin production in STZ-induced diabetic rats. The observations are consistent with the overall improvement of the complications associated with diabetes with insulin (Nathan et al., 2009). Streptozotocin-injected mice had significantly higher blood glucose level. Insulin only correct high blood sugar and some change in diabetic neuropathic pain in rats (Kuhad and Chopra, 2009).
The recorded significant decrease in triacylglycerols concentration in sciatic nerve observed in Streptozotocin-induced diabetic neuropathy in male rats are nearly similar to those of Zhu and Eichberg (1990) who reported that, the content of 1,2-diacylglycerol was reduced in sciatic nerves from streptozocin induced diabetic rats. Tsai et al. (1992) and Blank et al. (1993) demonstrated that, in cell culture systems triacylglycerols mediate the uptake and transfer of fatty acids to membrane phospholipids. Recently, Igal et al. (2001) have demonstrated that, during expressed mitochondrial glycerol 3-phosphate acyl carrier direct involvement of external fatty triacylgycerols rather than phospholipids. Treatment with α-lipoic acid combined with insulin to streptozotocin (STZ)-induced diabetic neuropathy in rats showed a significant increase in triacylglycerols concentration in sciatic nerve after six weeks of drugs administration when compared with the diabetic non-treated group. Similarly, Ozkan et al. (2005) showed that, brain lipid levels were higher in the diabetes 1, diabetes 2, diabetic triple antioxidant combination (vitamin E, vitamin C and α-lipoic acid) and α-lipoic acid groups than control. The increased levels in the diabetes 1 and triple antioxidant combination groups may be consequences of the regular insulin injections with distributed widely insulin receptors in the brain, with particularly high concentrations of neurons (Schwartz et al., 1992). Aplha-lipoic acid has metabolic as well as antioxidant effects (Dickinson et al., 2002; Manzella et al., 2001); it is a cofactor in the pyruvate dehydrogenase enzyme complex which catalyzes the synthesis of acetyl-CoA. Unsaturated fatty acids are synthesized by fatty acid desaturases; monounsaturated fatty acids are synthesized by stearoyl CoA desaturase (SCD) (Ntambi, 1995). Also, stearoyl CoA desaturase inserts a cis double bond at the delta-9 position of fatty acid substrates, a key rate-limiting step in the synthesis of unsaturated fatty acids. Palmitate and stearate are the preferred substrates are converted to Palmitulyat and oleate, respectively. These monounsaturated fatty acids are the main components of cell membrane phospholipids and triglycerides and stores found in adipose tissue (Kim and Ntambi, 1999). The significant increase in sciatic nerve non Esterified Fatty Acids (NEFA) concentration observed in Streptozotocin-induced diabetic neuropathy in male rats are nearly similar to those reported by King and Loeken (2004) who recorded that, Free Fatty Acids (FFA) which are elevated in diabetes and insulin resistance, may also contribute to the increased production of Reactive Oxygen Species (ROS) due to increased mitochondrial uncoupling and β-oxidation. In addition, high blood sugar and FFA lead induced oxidative stress to activate signaling pathways sensitive to stress. Ford et al. (2001) observed that, oxidative stress and defective fatty acid metabolism are important factors in the development of impaired nerve blood flow and function. Also, Chattopadhyay et al. (1992) reported that, alloxan causes diabetes in mice leads to increased a main fatty acids (NEFA) in the whole sciatic nerve endoneurium. Increased for NEFA levels are more pronounced throughout the diabetic nerve (40% of control) than it was part of her optic nerve sheath (20-30%). The observed changes in the composition of the phospholipid fatty acids as well, including an increase in the the linoleate (18:2 n-6) nervous fiber sheath phosphatidylserine and low arachidonate (20:4 n-6) in phosphatidylethanolamine and phosphatidylinositol nerve in people with diabetes. Based on total lipid phosphorous, NEFA levels in both whole nerve and sciatic endoneurium were as much as 40% higher than in control nerves. There exists very little information on the amounts and composition of NEFA in the peripheral nervous system. Arachidoic (20:4 n-6) and docosahexaenoic (22:6 n-3) acids are formed from their respective, parent compounds linoleic (18:2 n-6) and alpha linolenic (18:3 n-3) through a series of elongation and desaturation steps. Delta-6 desaturase catalyses the rate limiting first step reaction in the conversion of linoleic to arachidoic and alpha linolenic to docosahexaenoic and the final desaturation step in the synthesis of docosahexaenoic (Sprecher, 1992).
Treatment with α-lipoic acid in diabetic rats significantly decreased sciatic nerve (NEFA) after four weeks, followed by significant increase after six weeks. Treatment with insulin alone significantly decreased sciatic nerve (NEFA) after four weeks of drug administration. Also, the value of (NEFA) was significantly decreased in diabetic rats injected with α-lipoic acid combined with insulin all over the period of the experiments. These results are nearly similar to those reported by Gijoh et al. (1999) who reported that, α-lipoic acid administration to aged rats reduced the level of free fatty acids. In addition, α-lipoic acid promotes free fatty acid oxidation and thereby reduces the concentration of free fatty acids. Also, Lykkesfeldt et al. (1998) reported that, α-lipoic acid-supplemented aged rats showed a reduction in the levels of triglycerides and this may be attributed to an increase in the activity of lipoprotein lipase which in turn stimulates the clearance of triglycerides.
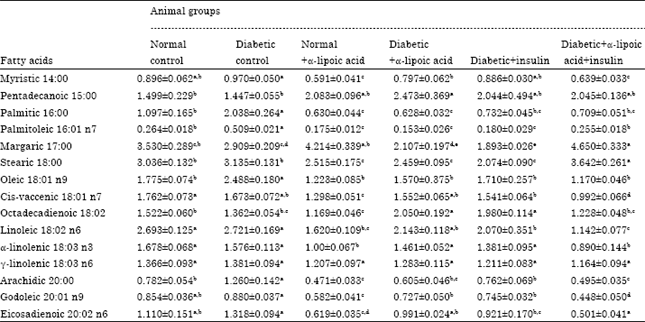
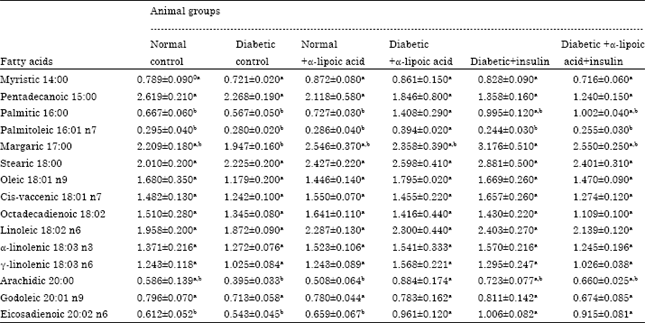
The obtained resuts demonstrated in (Tables 2 and 3) revealed that, a significant increase in palmitic, palmitoleic, oleic and arachidic were observed in sciatic nerve tissue of Streptozotocin-induced diabetic neuropathy in rats.
| Table 2: | Effects of four weeks treatment with alpha-lipoic acid, insulin and their combination on fatty acids composition percentage and main fatty acids changes of sciatic nerve in streptozotocin-induced diabetic neuropathy in male rats |
 | |
| Data are represented as | |
| Table 3: | Effects of six weeks treatment with alpha-lipoic acid, insulin and their combination on fatty acids composition percentage and main fatty acids changes of sciatic nerve in streptozotocin-induced diabetic neuropathy in male rats |
 | |
| Data are represented as | |
Ischemia and hypoxia are often associated with the liberation of fatty acids from complex lipids by the action of lipolytic enzymes, a process which is especially pronounced in the central nervous system. In addition, alterations in fatty acid metabolism are an important component of diabetes mellitus (Golay et al., 1987). The proposed changes in the metabolism of fatty acids as one factor in the development of diabetes metabolism (Cameron et al., 1991). Long chain fatty acid ester accumulation interferes with cell metabolism and membrane function (Brecher, 1983).
It has been demonstrated that accumulation of long-chain fatty acids and their esters in diabetic nerve may adversely effect phosphoinositide and PKC metabolism with secondary effects on Na+-K+-ATPase activities (Pugliese et al., 1991). Polyunsaturated fatty acids are important structural components of allocating liquidity and selective membranes (Douillet and Ciavatti, 1995). Deficiencies in both docosahexaenoic (22:6 n-3) and arachidonic acids (20:4 n-6) have been associated with disorders of neuro-visual development. Brenner (2003) reported that in experimental type I diabetes mellitus, the depressed delta-6 desaturase is restored by insulin which stimulates transcription of the gene.
Depression of the D-6 and D-5 enzyme Desaturases in diabetes is associated with lower arachidonic acid and the upper linoleic content in almost all tissues. Ozkan et al. (2005) observed that, oleic acid levels were decreased in both diabetic and non-diabetic rats, possibly because of reduced stearoyl CoA desaturase (SCD) activity (Kim and Ntambi, 1999). Douillet and Ciavatti (1995) found lower levels of monounsaturated fatty acids in the liver, aorta and plasma of diabetic rats. Thus, stearoyl CoA (SCD) desaturase his great physiological relevance of the organization, performance is sensitive to the change in diet, hormonal imbalance, development processes, changes in temperature, metals, alcohol, peroxisomal pro-liferators phenolic compounds (Ntambi, 1995; Kim and Ntambi, 1999). Changes in the ratio of 18:1/18:0 may be involved in various diseases including cancer, diabetes and neurological diseases.
There is increasing evidence that fatty acids act as mediators and modulators of central nervous system activity, perhaps because alterations in the composition of neuronal membranes change the membrane mobility. In addition, fatty acids may alter the brain microcirculation and modulate the production and activity of various neurotransmitters including some neuropeptides (Yehuda et al., 1998). In diabetes, the linoleic acid (18:2 n-6) concentration is generally a little above normal, whereas the eicosatrienoic (20:3 n-6) and arachidonic (20:4 n-6) acid concentrations are consistently below normal. When diabetics are treated with insulin, the concentrations of eicosatrienoic and arachidonic acids rise significantly. Some of the changes in phospholipid acyl groups appear to be due to decreased desaturase activity in diabetic tissue, including the Δ6 desaturase catalyzing the conversion of linoleic (18:2n-6) to γ-linolenic (18:3n-6) acid (Faas and Carter, 1990). Docosahexaenoic acid (22:6n-3) and arachidonic acid, (20: 4n-6) are found in high concentrations in structural lipids of the central nervous system and have been shown to be important for brain development and function (Innis, 1991; Uauy et al., 2001). Ruiz-Gutierrez et al. (1993) reported that, fatty acid composition is changed in humans and animals with diabetes. Diabetes inhibits delta-6-desaturase which converts linoleic acid, into gamma linolenic acid, the precursor of arachidonic acid and ultimately, several vasoactive prostanoids. In experimental and clinical diabetes. γ-linolenic production is reduced. Consequently the levels of Dihomo Gamma Linolenic Acid (DHGLA) which is a product of γ-linolenic elongation and arachidonic acid (Jamal, 1990). Moreover, diabetes reduces the rate of limiting desaturation steps, particularly delta-6 desaturation that converts linoleic acid to γ-linolenic acid and α-linolenic acid to stearidonic acid. Thus the availability of raw materials, essential fatty acids in Diabetes is further exacerbated by the increase in mass due to increased Reactive Oxygen Species (ROS) (Cameron and Cotter, 1997a). Essential fatty acids and especially arachidonic acid are reduced in peripheral nerve (Lowndes et al., 1990). Cotter and Cameron (1997) demonstrated that, the ability to provide sufficient amounts of long-chain polyunsaturated fatty acids such as arachidonic acid (20:4n-6) depends greatly on the conversion of linoleic acid (18:2n-6) to γ-linolenic acid (18:3n-6) via the rate-limiting enzyme Δ6-desaturase. Impaired 18:2n-6 metabolism is associated with diabetic neuropathy and altered prostanoid synthesis (Cameron et al., 1991) involved in the microvascular complications of diabetes (Jamal, 1994). In addition, diabetes impairs metabolic acids cut through the Δ6 and Δ5 desaturases enzyme activity, the enzyme that converts linoleic acid and dietary α-linolenic acid to Polyunsaturated Long-chain Fatty Acids (PUFA), including γ linolenic acid, arachidonic acid, eicosapentaenoic acid and docosahexaenoic acid (Horrobin, 1988). As a result, it has reduced levels of arachidonic acid and docosahexaenoic acid in membrane phospholipids of several tissues, including the sciatic nerve, red blood cells and in patients suffering from diabetes type 1 and diabetes in animals (Ruiz-Gutierrez et al., 1993). By changing membrane properties, PUFA can modify the activity of transmembrane enzymes, such as the Na, K-ATPase which is implicated in the propagation of nerve impulses. Sima and Sugimoto (1999) reported that, in human and experimental diabetes, the level of linoleic acid is normal. However, gamma linolenic acid is reduced as a result of a deficit in the enzyme delta 6 desaturates. This deficit is thought to be responsible for many of the microvascular changes in diabetic neuropathy. Moreover, Ghebremeskel et al. (2002) observed that, in diabetes there is a decrease in membrane arachidonic and docosahexaenoic acids a concomitant increase in linoleic and alpha-linolenic acids. This metabolic is thought to be due to impaired activity of Δ6 and Δ5-desaturases. Additionally, Diabetes impairs essential fatty acid metabolism by decreasing activities of desaturases and as a result, arachidonic acid and docosahexaenoic acids levels are reduced in membrane phospholipid of several tissues, including RBC and sciatic nerve, in patients with type 1 diabetes and in diabetes rats (Ruiz-Gutierrez et al., 1993). Furthermore, in the diabetic rats, the concentration of oleic acid was also observed to increase significantly. This observation was correlated with an earlier study that showed an increase in the concentration of oleic acid in the membrane of both type I and type II diabetic patients (Seigneur et al., 1994). Thus, the reduced availability of essential fatty acid intermediates in diabetes is further exacerbated by increased destruction due to elevated ROS (Cameron and Cotter, 1997b).
Administration of α-lipoic acid to normal rats significantly decreased the percentage of myristic, palmitice, palmitoleic, stearic, cis-vaccenic, octadecandienoic, linoleic, α-linolenic, arachidic, gadoleic and eicosadienoic in sciatic nerve after four weeks of treatment when compared with control normal rats. Treatment with α-lipoic acid alone to Streptozotocin (STZ)-induced diabetic neuropathy in rats significantly decreased the percentage of myristic, palmitic, palmitoleic, stearic, oleic, arachidic and gadoleic and significantly increase the percentage of pentadecanoic and octadecandienoic in sciatic nerve after four weeks of treatment. However, treatment with α-lipoic acid alone to streptozotocin (STZ)-induced diabetic rats significantly increased in percentage of palmitic, palmitoleic, arachidic and eicosadienoic acids in sciatic nerve membrane after six weeks of treatment when compared with diabetic non-treated group. Ozkan et al. (2005) observed that, the brain palmitic acid level was low in the diabetes 2 group. This decrease is related to the insufficient of insulin during the period of the experiment. The effects of insulin are associated with energy metabolism, notably the regulation of glucose and fatty acid metabolism. Insulin receptors are widely distributed n the brain, with particularly high concentrations in neurons (Schwartz et al., 1992). Freychet (2000) stated that insulin was the first hormone to be implicated in the control of body weight by the central nervous system. While the palmitic acid level rose in the non-diabetic α-lipoic acid group. The increase palmitic acid level in the α-lipoic acid group may be due to the metabolic effect of α-lipoic acid. Alpha-lipoic acid has metabolic as well as antioxidant effects (Dickinson et al., 2002; Manzella et al., 2001); it is a cofactor in the pyruvate dehydrogenase enzyme complex which catalyzes the synthesis of acetyl-CoA.
Treatment with insulin alone to Streptozotocin (STZ)-induced diabetic neuropathy in rats significantly decreased the percentage of palmitic, palmitoleic, margaric, stearic, oleic, linoleic, arachidic, gadoleic and eicosadienoic acids with significant increase in the percentage of octadecandienoic acid in sciatic nerve membrane after four weeks of treatment as well as increase in the percentage of margaric and eicosadienoic acids after six weeks of treatment when compared with diabetic non treated rats group. In diabetes, the linoleic acid (18:2 n-6) concentration is generally a little above normal, whereas, the eicosatrienoic (20:3 n-6) and arachidonic (20:4 n-6) acid concentrations are consistently below normal. When diabetics are treated with insulin, the concentrations of eicosatrienoic and arachidonic acids rise significantly. Brenner (2003) reported that, in experimental type I diabetes mellitus, the depressed delta-6 desaturase is restored by insulin which stimulates transcription of the gene. The fatty acid composition of cell membranes can influence membrane-associated phenomena such as the interaction between insulin and its receptor (Stubbs and Smith, 1984).
Treatment with α-lipoic acid combined with insulin in Streptozotocin (STZ)-induced diabetic neuropathy in rats significantly decreased the percentage of myristic, palmitic, palmitoleic, oleic, cis-vaccenic, linoleic, α-linolenic, arachidic and gadoleic acids, with significantly increased the percentage of margaric and stearic acids in sciatic nerve membrane after four weeks and eicosadienoic acid after six weeks of treatment when compared with diabetic non treated rats group. Similarly, Ozkan et al. (2005) suggested that, the brain lipid levels in the diabetic+vitamin E, vitamin C and lipoic acid (D+VEVCALA), diabetes 1 and diabetes 2 groups were higher than in the control group. Total lipid was also higher in the non-diabetic rats treated with alpha-lipoic acid and vitamin C (VC). Total cholesterol was higher in the diabetes 1 and diabetes 2 groups than in controls. Who also added that, in respect of fatty acid composition, palmitic acid levels were lower in the diabetes 2 and non-diabetic vitamin E (VE) groups than the control group but higher in the non-diabetic alpha-lipoic acid group. Oleic acid (18:1 n-9) levels were in the diabetic and non-diabetic group, than the control group but higher in the non-diabetic alpha-lipoic acid group. Arachidonic acid (20:4 n-6) levels were similar in the diabetes 1 D+VEVCALA and control groups but higher in the non-diabetic VE, VC, ALA and VEVCLA groups and lower in the diabetes 2 group. Docosahexaenoic acid (22:6 n-3) was elevated in the diabetes 2 and VEVCALA groups. Levels of arachidonic and docosahexaenoic acids in brain may be decreased by the effects of cancer, diabetes and some degenerative diseases. Desaturase activities in the D-6 pathway may be increased by insulin and diet restriction (Frenkel et al., 2004). Insulin therapy can correct this defect by influencing the desaturase level. The obtained results confirm that antioxidants and insulin treatment have beneficial effects on the D-6 desaturase system and unsaturated fatty acid levels. Antioxidants may have protective effects on unsaturated fatty acids and that insulin exerts important effects on enzymes in the D-6 pathway (Brenner, 2003; Cho et al., 1999).
They may also reduce oxidative stress and thus impair progression to LDL oxidation, cell membrane lipid peroxidation and decreased endoneurial blood flow, thereby reducing peripheral nerve and vascular dysfunction. Alpha-lipoic acid is a naturally occurring short chain fatty acid with sulfhydryl groups that has potent antioxidant activity and is clinically used to treat diabetic neuropathy (Vessal et al., 2003). Alpha-lipoic acid is a vital cofactor in the multienzyme complexes that catalyze the oxidative decarboxylation of α-keto acids. It plays an important role in lipid biosynthesis by replacing coenzyme. In addition to its cofactor role, α-lipoic acid is a powerful antioxidant and possesses numerous cellular functions as well as beneficial effects in conditions with elevated oxidative stress (Arivazhagan et al., 2003). Celik and Ozkaya (2002) showed that, lipoic acid administration eventually results in a decrease in 14:0, 16:0 and total saturated fatty acid and an increase in 18:2, 18:3, 20:4, 22:6, total unsaturated fatty acid and PUFA levels, thus substantiating the antioxidant properties of lipoic acid. Moreover, who illustrates that, lipoate administration may be more effective than vitamin E in preventing lipid peroxidation in tissues. This may be attributed to the bioactivity of lipoic acid directly reacting with various reactive oxygen species, as well as its ability to interfere with oxidation processes in lipid and the aqueous cellular compartment (Packer et al., 1995).
CONCLUSION
From the obtained results it could be concluded that, distinct abnormalities of fatty acids composition was observed in STZ-induced diabetic neuropathy in rats. Also, in a STZ-treated rat model of DNP, biochemical analysis of sciatic nerve myelin lipids revealed that diabetes alters myelin’s fibers of sciatic nerve NEFA, triacylglycerols and fatty acid composition contents in a pattern that can change membrane fluidity. It may be possible that, treatment with α-lipoic acid combined with insulin administrations could ameliorate the altered myelin of sciatic nerve lipids induced by diabetes and attenuate the changed fatty acid composition of the membrane and improved the metabolic control of diabetic neuropathy.
REFERENCES
- Kuhad, A. and K. Chopra, 2009. Tocotrienol attenuates oxidative-nitrosative stress and inflammatory cascade in experimental model of diabetic neuropathy. Neuropharmacology, 57: 456-462.
CrossRefPubMedDirect Link - Arivazhagan, P., K. Ramanathan and C. Panneerselvam, 2001. Effect of DL-α-lipoic acid on the status of lipid peroxidation and antioxidants in mitochondria of aged rats. J. Nutr. Biochem., 12: 2-6.
CrossRefPubMedDirect Link - Blank, M.L., Z.L. Smith and E. Snyder, 1993. Arachidonate-containing triacylglycerols: Biosynthesis and a lipolytic mechanism for the release and transfer of arachidonate to phospholipids in HL-60 cells. Biochim. Biophys. Acta, 1170: 275-282.
PubMedDirect Link - Brecher, P., 1983. The interaction of long-chain acyl CoA with membranes. Mol. Cell. Biochem., 57: 3-15.
PubMedDirect Link - Brenner, R.R., 2003. Hormonal modulation of Δ6 and Δ5 desaturases: Case of diabetes. Prostaglandins Leukotrienes Essent. Fatty Acids, 68: 151-162.
CrossRefDirect Link - Cameron, N.E. and M.A. Cotter, 1997. Metabolic and vascular factors in the pathogenesis of diabetic neuropathy. Diabetes, 46: S31-S37.
PubMedDirect Link - Cameron, N.E. and M.A. Cottter, 1997. Effects of antioxidants on nerve and vascular dysfunction in experimental diabetes. Diabetes Res. Clin. Pract., 45: 137-146.
PubMedDirect Link - Cameron, N.E., M.A. Cotter and S. Robertson, 1991. Essential fatty acid diet supplementation: Effects on peripheral nerve and skeletal muscle function and capillarization in streptozocin-induced diabetic rats. Diabetes, 40: 532-539.
CrossRefDirect Link - Celik, S. and A. Ozkaya, 2002. Effects of intraperitoneally administered lipoic acid, vitamin E and linalool on the level of total lipid and fatty acids in guinea pig brain with oxidative stress induced by H2O2. J. Biochem. Mol. Biol., 35: 547-552.
PubMed - Chitra, V., P.V. Varma, K.A. Raju and K.J. Prakash, 2010. Study of antidiabetic and free radical scavenging activity of the seed extract of Strychnos nuxvomica. Int. J. Pharmacy Pharma. Sci., 2: 106-110.
Direct Link - Cho, H.P., M.T. Nakamura and S.D. Clarke, 1999. Cloning, expression and fatty acid regulation of the human delta-5 desaturase. J. Biol. Chem., 274: 37335-37339.
PubMedDirect Link - Cotter, M.A. and N.E. Cameron, 1997. Effects of dietary supplementation with arachidonic acid rich oils on nerve conduction and blood flow in streptozotocin-diabetic rats. Prostaglandins Leukot. Essent. Fatty Acids, 56: 337-343.
PubMedDirect Link - Deferrari, G., M. Ravera and V. Berruti, 2003. Treatment of diabetic nephropathy in its early stages. Diabetes. Metab. Res. Rev., 19: 101-114.
CrossRefPubMedDirect Link - Dias, A.S., M. Porawski, P.S. Collado and J.G. Gallego, 2005. Quercetin decreases oxidative stress, NF-kappaB activation and iNOS overexpression in liver of streptozotocin-induced diabetic rats. J. Nutr., 135: 2299-2304.
PubMedDirect Link - Dickinson, P.J., A.L. Carrington, G.S. Frost and A.J.M. Boulton, 2002. Neurovascular disease, antioxidants and glycation in diabetes. Diabetes Metab. Res. Rev., 18: 260-272.
Direct Link - Douillet, C. and M. Ciavatti, 1995. Effect of vitamin E treatment on tissue fatty acids and cholesterol content in experimental diabetes. J. Nutr. Biochem., 6: 319-326.
CrossRef - Edwards, J.L., A.M. Vincent, H.T. Cheng and E.L. Feldman, 2008. Diabetic neuropathy: Mechanisms to management. Pharmacol. Ther., 120: 1-34.
CrossRefPubMedDirect Link - Peuchant, E., R. Wolff, C. Salles and R. Jensen, 1989. One-step extraction of human erythrocyte lipids Allowing rapid determination of fatty acid composition. Anal. Biochem., 181: 341-344.
CrossRefDirect Link - Faas, F.H. and W.J. Carter, 1990. Altered fatty acid desaturation and microsomal fatty acid composition in the streptozotocin diabetic rat. Lipids, 15: 953-961.
CrossRefDirect Link - Fossati, P. and L. Prencipe, 1982. Serum triglycerides determined colorimetrically with an enzyme that produces hydrogen peroxide. Clin. Chem., 28: 2077-2080.
CrossRefPubMedDirect Link - Frenkel, A.L., L. Canetti and Z. Halpern, 2004. Effects of nutritional lipids on diabetic manifestations and Δ6 desaturase mRNA level in streptozotocin treated mice. Nutr. Res., 24: 303-312.
CrossRefDirect Link - Freychet, P., 2000. Insulin receptors and insulin actions in the nervous system. Diabetes Metab. Res. Rev., 16: 390-392.
CrossRef - Ghebremeskel, K., D. Bitsanis, E. Koukkou, C. Lowy, L. Poston and M.A. Crawford, 2002. Liver triacylglycerols and free fatty acids in streptozotocin-induced diabetic rats have atypical n-6 and n-3 pattern. Comp. Biochem. Physiol. C Toxicol. Pharmacol., 132: 349-354.
CrossRefPubMedDirect Link - Gijoh, M.A., D.M. Spencer, A.L. Kaiser and C.C. Leslie, 1999. Role of phosphorylation sites and the C2 domain in regulation of cytosolic phospholipase A2. J. Cell Biol., 145: 1219-1232.
PubMedDirect Link - Golay, A., A.L. Swislocki, Y.D. Chen and G.M. Reaven, 1987. Relationships between plasma-free fatty acid concentration, endogenous glucose production and fasting hyperglycemia in normal and non-insulin-dependent diabetic individuals. Metabolism, 36: 692-696.
CrossRefPubMedDirect Link - Craven, P.A., R.K. Studer, H. Negrete and F.R. DeRubertis, 1995. Protein kinase C in diabetic nephropathy. J. Diabetes Complication, 9: 241-245.
CrossRefDirect Link - Greene, D.A., S.A. Lattimer and A.A. Sima, 1999. Sorbitol, phosphoinositides and sodium-potassium-ATPase in the pathogenesis of diabetic complications. N. Engl. J. Med., 316: 599-606.
PubMedDirect Link - Gruzman, A., A. Hidmi, J. Katzhendler, A. Haj-Yehie and S. Sasson, 2004. Synthesis and characterization of new and potent Α-lipoic acid derivatives. Bioorg. Med. Chem., 12: 1183-1190.
CrossRefDirect Link - Horrobin, D.F., 1988. The roles of essential fatty acids in the development of diabetic neuropathy and other complications of diabetes mellitus. Prostaglandins Leukot Essent. Fatty Acides, 31: 181-197.
PubMedDirect Link - Innis, S.M., 1991. Essential fatty acids in growth and development. Prog. Lipid. Res., 30: 39-103.
CrossRefPubMedDirect Link - Izbeki, F., T. Wittman, A. Rosztoczy, N. Linke, N. Bodi, E. Fekete and M. Bagyanszki, 2008. Immediate insulin treatment prevents gut motility alterations and loss of nitrergic neurons in the ileum and colon of rats with Streptozotocin-induced diabetes. Diab. Res. Clin. Pract., 80: 192-198.
CrossRefDirect Link - Jamal, G.A., 1994. The use of gamma linolenic acid in the prevention and treatment of diabetic neuropathy. Diabet. Med., 11: 145-149.
CrossRefPubMedDirect Link - Jamal, G.A., 1990. Pathogenesis of diabetic neuropathy: The role of the n-6 essential fatty acids and their eicosanoid derivatives. Diabet. Med., 7: 574-579.
PubMedDirect Link - Chattopadhyay, J., E.W. Thompson and H.H. Schmid, 1992. Nonesterified fatty acids in normal and diabetic rat sciatic nerve. Lipids, 27: 513-517.
PubMedDirect Link - Kakadiya, J., M. Shah and N. Shah, 2010. Glimepiride reduces on experimentally induced is chemical reperfusion in diabetic rats. Int. J. Applied Biol. Pharm. Technol., 1: 275-285.
Direct Link - Kim, Y.C. and J.M. Ntambi, 1999. Regulation of stearoyl-CoA desaturase genes: Role in cellular metabolism and preadipocyte differentiation. Biochem. Biophys. Res. Commun., 266: 1-4.
CrossRef - King, G.L. and M.R. Loeken, 2004. Hyperglycemia-induced oxidative stress in diabetic complications. Histochem. Cell Biol., 122: 333-338.
CrossRefPubMedDirect Link - Lowndes, J.M., M. Hokin-Neaverson and P.J. Bertics, 1990. Kinetics of phosphorylation of Na+/K(+)-ATPase by protein kinase C. Biochim. Biophys. Acta, 1052: 143-151.
CrossRefPubMedDirect Link - Lykkesfeldt, J., T.M. Hagen, V. Vinarsky and B.N. Ames, 1998. Age-associated decline in ascorbic acid concentration, recycling and biosynthesis in rat hepatocytes-reversal with (R)-α -lipoic acid supplementation. FASEB J., 12: 1183-1189.
Direct Link - Manzella, D., M. Barbieri, E. Ragno and G. Paolisso, 2001. Chronic administration of pharmacologic doses of vitamin E improves the cardiac autonomic nervous system in patients with type 2 diabetes. Am. J. Clin. Nutr., 73: 1052-1057.
PubMed - Nathan, D.M., B. Zinman, R. Miller and T.J. Orchard, 2009. Modern-day clinical course of type 1 diabetes mellitus after 30 years duration: The diabetes control and complications trial/epidemiology of diabetes interventions and complications and Pittsburgh epidemiology of diabetes complications experience (1983-2005). Arch. Intern. Med., 169: 1307-1316.
CrossRefPubMed - Negi, G., A. Kumar, R.K. Kaundal, A. Gulati and S.S. Sharma, 2010. Functional and biochemical evidence indicating beneficial effect of Melatonin and Nicotinamide alone and in combination in experimental diabetic neuropathy. Neuropharmacology, 58: 585-592.
CrossRefPubMedDirect Link - Ozkan, Y., O. Yilmaz, A.I. Ozturk and Y. Ersan, 2005. Effects of triple antioxidant combination (vitamin E, vitamin C and α-lipoic acid) with insulin on lipid and cholesterol levels and fatty acid composition of brain tissue in experimental diabetic and non-diabetic rats. Cell Biol. Int., 29: 754-760.
CrossRefPubMedDirect Link - Pugliese, G., R.G. Tilton and J.R. Williamson, 1991. Glucose-induced metabolic imbalances in the pathogenesis of diabetic vascular disease. Diabetes Metab. Rev., 7: 35-59.
CrossRefPubMedDirect Link - Ramanathan, M., A.K. Jaiswal and S.K. Bhattacharya, 1999. Superoxide dismutase, catalase and glutathione peroxidase activities in the brain of streptozotocin induced diabetic rats. Indian. J. Exp. Biol., 37: 182-183.
PubMed - Ruiz-Gutierrez, V., P. Stiefel, J. Villar, M.A. Garcia-Donas, D. Acosta and J. Carneado, 1993. Cell membrane fatty acid composition in type 1 (insulin-dependent) diabetic patients: Relationship with sodium transport abnormalities and metabolic control. Diabetologia, 36: 850-859.
CrossRefPubMedDirect Link - Said, G., 2007. Diabetic neuropathy: A review. Nat. Clin. Pract. Neurol., 3: 331-340.
CrossRefPubMedDirect Link - Schwartz, M.W., D.P. Figlewicz, D.G. Baskin, S.C. Woods and D. Porte Jr., 1992. Insulin in the brain: A hormonal regulator of energy balance. Endocr. Rev., 13: 387-414.
CrossRefPubMedDirect Link - Seigneur, M., G. Freyburger, H. Gin, M. Claverie and D. Lardeau et al., 1994. Serum fatty acid profiles in type I and type II diabetes: Metabolic alterations of fatty acids of the main serum lipids. Diabetes Res. Clin. Pract., 23: 169-177.
CrossRefDirect Link - Sharma, V.K., S. Kumar, H.J. Patel and S. Hugar, 2010. Hypoglycemic activity of Ficus glomerata in alloxan induced diabetic rats. Int. J. Pharm. Sci. Rev. Res., 1: 18-22.
Direct Link - Sima, A.A.F. and K. Sugimoto, 1999. Experimental diabetic neuropathy: An update. Diabetologia, 42: 773-788.
CrossRefDirect Link - Skyler, J.S., 2007. Prediction and prevention of type 1 diabetes: Progress, problems and prospects. Clin. Pharmacol. Ther., 81: 768-771.
PubMedDirect Link - Smith, A.G. and J.R. Singleton, 2008. Impaired glucose tolerance and neuropathy. Neurologist, 14: 23-29.
CrossRefPubMedDirect Link - Stubbs, C.D. and A.D. Smith, 1984. The modification of mammalian membrane polyunsaturated fatty acid composition in relation to membrane fluidity and function. Biochimica Biophysica Acta, 779: 89-137.
CrossRefPubMedDirect Link - Tomlinson, D.R. and N.J. Gardiner, 2008. Glucose neurotoxicity. Nat. Rev. Neurosci., 9: 36-45.
PubMedDirect Link - Trinder, P., 1969. Determination of glucose in blood using glucose oxidase with an alternative oxygen acceptor. Ann. Clin. Biochem., 6: 24-27.
CrossRefDirect Link - Tsai, S., A.Y. Sun, H.D. Kim and G.Y. Sun, 1992. Manganese exposure to PC-12 cells alters triacylglycerol metabolism and promotes neurite outgrowth. Life Sci., 52: 1567-1575.
PubMedDirect Link - Uauy, R., D.R. Hoffman, P. Peirano, D.G. Birch and E.E. Birch, 2001. Essential fatty acids in visual and brain development. Lipids, 36: 885-895.
CrossRefDirect Link - Vessal, M., M. Hemmati and M. Vasei, 2003. Antidiabetic effects of quercetin in streptozocin-induced diabetic rats. Comp. Biochem. Physiol. Part C: Toxicol. Pharmacol., 135: 357-364.
CrossRefDirect Link - Vincent, A.M., J.W. Russell, P. Low and E.L. Feldman, 2004. Oxidative stress in the pathogenesis of diabetic neuropathy. Endocrin Rev., 25: 612-628.
CrossRefPubMedDirect Link - Williamson, J.R., K. Chang, M. Frangos, K.S. Hasan and Y. Ido et al., 1993. Hyperglycemic pseudohypoxia and diabetic complications. Diabetes, 42: 801-813.
PubMedDirect Link - Yehuda, S., S. Rabinovitz, R.L. Carasso and D.I. Mostofsky, 2002. The role of polyunsaturated fatty acids in restoring the aging neuronal membrane. Neurobiol. Aging., 23: 843-853.
PubMedDirect Link - Zhu, X. and J. Eichberg, 1990. 1,2-diacylglycerol content and its arachidonyl-containing molecular species are reduced in sciatic nerve from streptozotocin-induced diabetic rats. J. Neurochem., 55: 1087-1090.
PubMedDirect Link - Pugliese, G., R.G. Tilton and J.R. Williamson, 1991. Glucose-induced metabolic imbalances in the pathogenesis of diabetic vascular disease. Diabetes Metab. Rev., 7: 35-59.
CrossRefPubMedDirect Link - Pugliese, G., R.G. Tilton and J.R. Williamson, 1991. Glucose-induced metabolic imbalances in the pathogenesis of diabetic vascular disease. Diabetes Metab. Rev., 7: 35-59.
CrossRefPubMedDirect Link - Maritim, A.C., R.A. Sanders and J.B. Watkins, 2003. Effects of alpha-lipoic acid on biomarkers of oxidative stress in streptozotocin-induced diabetic rats. J. Nutr. Biochem., 14: 288-294.
Direct Link