
P. Banerjee
Department of Biotechnology and Dr. B.C. Guha Centre for Genetic Engineering and Biotechnology, India
K. Panda
Department of Biotechnology and Dr. B.C. Guha Centre for Genetic Engineering and Biotechnology, India
P. Nandi
Department of Environmental Science, 3Department of Biochemistry, University of Calcutta, 35, Ballygunge Circular Road, Kolkata-700019, India
S.K. Banerjee
Department of Biotechnology and Dr. B.C. Guha Centre for Genetic Engineering and Biotechnology, India
Asian Journal of Biochemistry
Year: 2013 | Volume: 8 | Issue: 1 | Page No.: 1-13







ABSTRACT
Health damaging effects of carbonated soft drink are largely based on epidemiological observations. The present study aimed to examine the possible physiological and biochemical effects of such drink in animal. Through oxiblot assay, fluorescence activated cell sorting assay and other biochemical investigations; it has been shown that consumption of such drink makes oxidative damages. Extensive such damages at the level of proteins in vivo in the liver, kidney and blood of experimental animal, guinea pigs, which cannot synthesize vitamin C like humans have been observed. These effects are also associated with significant cellular damages. In vitro studies reviled that such drink contains oxidants those are capable of direct protein oxidation. These observations establish physiological and biochemical link between consumption of carbonated soft drink and its deleterious effects in vitamin C deficient animals.
PDF Abstract XML References Citation
Received: September 06, 2012;
Accepted: January 23, 2013;
Published: May 25, 2013
How to cite this article
P. Banerjee, K. Panda, P. Nandi and S.K. Banerjee, 2013. Oxidative Damage of Liver, Kidney and Serum Proteins with Apoptosis of above Tissues in Guinea Pigs Fed on Carbonated Soft Drink. Asian Journal of Biochemistry, 8: 1-13.
DOI: 10.3923/ajb.2013.1.13
URL: https://scialert.net/abstract/?doi=ajb.2013.1.13
DOI: 10.3923/ajb.2013.1.13
URL: https://scialert.net/abstract/?doi=ajb.2013.1.13
INTRODUCTION
Carbonated soft drinks have rapidly spread their influence in world’s consumer market and these are almost a part of our daily food habits in social life (Teerasong et al., 2010). The main active ingredients in these carbonated soft drinks are carbonated water, high sugar or fructose corn syrup, phosphoric acid, caramel color, natural flavors and caffeine (Encyclopaedia Britannica, 2012). Consumption of carbonated soft drinks have been implicated with several health damaging effects including obesity, (Malik et al., 2006) type II diabetes, (Vartaman et al., 2007) metabolic syndrome risk, (Yoo et al., 2004) elevated blood pressure (Raben et al., 2002) osteoporosis (Heaney and Rafferty, 2001) and hypokalemia (Tsimihodimos et al., 2009). Excess sugar consumption increases production of destructive free radicals and lowers level of key antioxidants, as reported by Mohanty et al. (2000). Type II diabetes resulting from high sugar consumption is also associated with an increase in free radical generation, leading to damage of fats, proteins and DNA. Fructose consumption might result an impact on the development of insulin resistance and cardiovascular diseases if taken in large volume (Tappy and Le, 2010). Fructose directly stimulates the endothelial inflammatory process and reduce Nitric Oxide (NO) levels in human aortic endothelial cells in vitro (Glushakova et al., 2008). High fructose feeding in rats showed increased level of oxidative stress markers and increased reactive oxygen species in circulatory leukocytes (Al-Awwadi et al., 2005). The antioxidant ability of caffeine has been reported in contrast with its pro oxidant effects derived from its action mechanism such as systemic release of catecholamines (Olcina et al., 2006). Phosphoric acid toxicity has been reported to accelerate mammalian aging (Ohnishi and Raijaque, 2010) consumption of soft drink has deleterious effects on oral mucosa in rats due to its phosphoric acid content (Kapicloglu et al., 2000). It has been reported that phosphoric acid in soft drink is associated with increased risk for chronic kidney disease (Saldana et al., 2007). Effect of cola drink on fertility has been reviewed (Imai et al., 2010).
Though epidemiological studies have been made over the years to investigate consumption of carbonated soft drinks and their possible effects on human health, so far studies on the effects on carbonated soft drinks at the biochemical and cellular level are scanty (Amato et al., 1998; Milei et al., 2011). Recently effect of carbonated soft drinks on the activities of alanine amino transferase and aspartate amino transferase in serum and kidney in rats has been reported (Jeroh et al., 2012) and effect of coke consumption on lipid peroxide and alkaline phosphatase activity in serum and liver of animal has been reported (Awhin, 2012).
The present study was designed to ascertain the effects of carbonated soft drink, Carbonated soft drink at the cellular and biochemical levels in guinea pigs, animal which cannot synthesize vitamin C, like humans and are suitable for studying oxidative damages. The experimental approach were made to investigate impact of such drink on proteins, DNA and in cellular apoptosis in experimental animal. Carbonated soft drink obtained from Indian market was fed to the experimental animal, guinea pigs for a period of 0 to 60 days.
MATERIALS AND METHODS
Carbonated soft drink: Carbonated soft drink, available in Indian market in 600 mL bottle, was used to feed the animal.
Chemicals and reagents: Sodium Dodecyl Sulphate (SDS), acrylamide, bisacrylamide, Ammonium Persulphate (APS), N,N,N’,N’-tetramethyl ethylene diamine (TEMED), Coomasie brilliant blue, bovine serum albumin (BSA), 5,5’ dithiobis 2-nitrobenzoic acid (DTNB), polyethylene sulphonyl fluoride (PMSF), ethylene diamine tetraacetate (EDTA), potassium chloride, sodium hydroxide, 2,4-dinitrophenylhydrazine (DNPH), hydrocholoric acid, trichloroacetic acid (TCA), ethanol, ethyl acetate, guanidium hydrochloride, 4-(2-hydroxyethyl)-1-piperazineethanesulfonic acid (HEPES), n-hexane, formalin, paraffin, xylene, ethanol, sodium citrate, triton X-100, fluorescein (Roche), 4, 6'-diamidino-2-phenylindole (DAPI), Protein Reagent (Bio-Rad, USA), sodium chloride, potassium chloride, disodium hydrogen phosphate, potassium dihydrogen phosphate, Tris-HCl, dithiothreitol (DTT), Tween 20, glycerol, glycine, methanol, skimmed milk, 1° Antibody: Rabbit Anti-DNP Antibody and 2° Antibody: Goat Anti-Rabbit IgG (HRP-conjugated), Electrogenerated Chemiluminescent (ECL) detection kit (Amersham Biosciences), acetic acid, heparin, 1xFluorescence-Activated Cell Sorting (FACS) lysis buffer, 1xFACS binding buffer, annexin V-Flurescein isothiocyanate (FITC), propidium iodide staining solution (PI), Beta-Mercaptoethanol (BME), bromophenol blue, sodium acetate, sodium thiosulphate, glutaraldehyde, silver nitrate, formaldehyde.
All the above reagents were of analytical grade and were purchased from the Sigma Chemical Company, E. Merck Ltd and SRL (India).
Animals: Young male guinea pigs, weighing 450-500 g were used for all experimental purposes. They were of pure breed with white and black and white and brown colors. They were housed in well circulated, clean animal house at comfortable temperature, with 12 h light and 12 h dark conditions. There were separate cage was used for each guinea pig. All results are of three different sets of experiments and each set consists of three control and fifteen experimental animals. Guinea pigs have been used because many of the metabolic characteristics in guinea pigs are similar to that of humans (Stith and Das, 1982) including that gunieas pigs cannot smnthesise vitamin C like humans and are thus suitable animal for the purpose of studying cellular oxidative damage.
Both control and experimental animals for the in vivo studies on the effect of carbonated soft drink-induced oxidative damage were fed a special stock diet with following composition: Wheat bran: 70%; Casein: 20%; Sucrose: 8%; AOAC vitamin mixture (without ascorbate): 1%; AOAC Salt mixture: 1%. The same diet was prepared manually with pure ingredients. This was done as per procedure of NIH guidelines (NRC, 1985). Both control and experimental animals were supplemented with vitamin C (2 mg day-1) along with either water (control group) or carbonated soft drink (experimental group) to protect the animals from scurvy.
The volumes of carbonated soft drink which were administered orally to guinea pigs varied from 6-8 mL depending on body weight of the animal. Here volume of the carbonated soft drink was determined proportionately assuming average body weight of guinea pigs 500 g compared to 600-1200 mL soft drink consumption per day by an individual of average 60 kg b.wt. in India. All animal treatment procedures met the NIH guidelines (NRC, 1985) and were conducted with institutional animal ethics committee approval.
Oxyblot assay: Oxidative damage of liver tissue and serum proteins were evidenced by immunoblotting of the dinitrophenylhydrazone derivatives of protein carbonyls (Misra et al., 2003). Whole liver tissue weighing 1 g was first minced and suspended in 9 mL of lysis buffer then homogenized, followed by centrifugation at 16,500 g for 15 min at 4°C. This process of centrifugation was repeated thrice until a clear whole tissue extract solution was formed. Lysates were spun to remove all insoluble materials. The protein concentration was approximately maintained at 50-60 μg for serum and 80-90 μg for liver tissue according to requirement. Protein concentration was measured using Bio-Rad protein estimation kit. SDS solution (20%) in equal volume was added to the sample extract and after 5 min incubation, 30 μL of 2, 4-dinitrophenyl-hydrazine (DNPH) solution was added and the reaction mixture was incubated for 15 min at room temperature. This was neutralized with 22.5 μL of neutralizing solution as per protocols defined in the oxyblot protein oxidation detection kit (Intergen company, NY, USA). This treated tissue extract was then separated by SDS-PAGE and blotted on a PVDF (polyvinylidene fluoride) membrane. The membrane was then incubated with primary antibody, specific to the DNP-moiety, after blocking with 5% nonfat milk. This step was followed by incubation with a secondary antibody conjugated with horseradish peroxidase (goat anti-rabbit IgG) directed against the primary antibody.
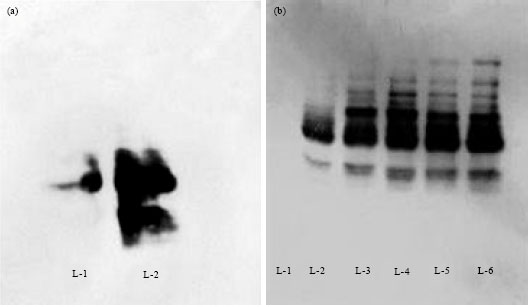 | |
| Fig. 1(a-b): | Oxyblot of liver proteins of guinea pigs administered carbonated soft drink against water as control, (a) after 0 and 30th day of administered orally for 30 days, L-1: water for 30 days, L-2: Carbonated soft drink for 30 days, (b) Time dependent manner after 0th, 7th, 15th, 30th ,45th and 60th days, of administion, L-1: water as the drink, L-2: Carbonated soft drink for 7 days, L-3: Carbonated soft drink for 15 days, L-4: Carbonated soft drink for 30 days, L-5: Carbonated soft drink for 45 days, L-6: Carbonated soft drink for 60 days |
This membrane was then treated with chemiluminescent reagent ECL chemiluminescent detection kit (GE Healthcare, USA) and the chemiluminescent signals from the responsive bands captured by autoradiography on a photographic film, as presented in Fig. 1a, b and Fig. 2a, b.
Terminal deoxyncleotidyl Transferase (TUNEL) assay for assessing cellular damage of liver and kidney tissue: The paraffin embedded tissue sections (5 μm) were deparaffinized, washed and permeabilized. The measurement of apoptotic cells was carried out using ‘In situ cell death detection kit, fluorescein’ (Roche) to exclusively determine cellular apoptosis based on Terminal deoxynucleotidyl transferase mediated 2'-deoxy Uridene 5' triphosphate (dUTP) Nick End Labelling (TUNEL) reaction (Banerjee et al., 2008). After the reaction, the slides were washed with phosphate buffer saline and DNA fragmentation was detected by labeling with fluorescein labelled dUTP using terminal deoxynucleotidyl transferase. The cells were examined using a fluorescence microscope (Olympus Bx40) at an excitation wavelength of 488 nm. Digital images were captured with cool CCD camera (Olympus; magnification, 10x). The nuclei were counted by counter staining with 4', 6'-diamidino-2-phenylindole (DAPI) at excitation wavelength of 350 nm, as presented in Fig. 3 and 4 .
Fluorescence-activated cell sorting assay (FACS assay): FACS assay (Das et al., 2011) was conducted to determine oxidative damage. Soon after the animals were sacrificed; a part of their blood was instantaneously collected in heparin.
 | |
| Fig. 2(a-b): | (a) Oxyblot and (b) Silver stain of serum proteins of guinea pigs administered carbonated soft drink against water as control, orally between 0-60 days, of administerated. L-1: water as the drink, L-2: Carbonated soft drink for 30 days, L-3: Carbonated soft drink for 60 days. Here beyond 30 days, feeding unto 60 days not only brought damages in the serum protein but also led to protein degradation (Fig. 2b) |
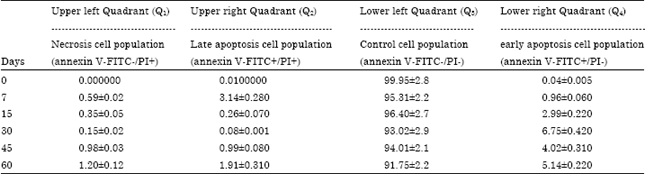
The blood was then centrifuged at 250 g for 10 min at room temperature. The precipitate containing blood cells was then collected and to it 1xFACS lysis buffer solution was added and it was then again centrifuged at 250 g for 10 min at room temperature. Then cellular precipitate was washed twice with 1x phosphate buffer saline and was centrifuged at 250 g for 10 min at room temperature. Finally the blood cells were suspended in 1x FACS binding buffer. From which 100 μL of the solution was transferred in FACS culture tube and to it 3-5 μL of annexin V-FITC (Boehringer Mannheim) and propidium iodide staining solution (PI) were added parellely according to manufacturer’s protocols. Then the solution was gently vortexed and was incubated for 15 min at room temperature in dark. After the incubation period 400 μL of 1xFACS Ca2+ enriched binding buffer was added to it to make a total volume of 500 μL and then it was directly subjected to FACS analysis. FACS Calibur, Becton Dickinson, San Jose, CA. Data was acquired and analysed using the Cell Quest programme (Becton Dickinson). Electronic compensation of the instrument was done to exclude overlapping of the emission spectra. Total 10,000 events were acquired, the cells were properly gated and dual parameter dot plot of FL2-H linear (X-axis; Flous-fluorescence) versus FL1-H linear (Y-axis; PI-fluorescence) was observed. PI and Annexin V-FITC emissions were detected in the FL 1 and FL 2 channels of FACS Calibur flow cytometer, using emission filters of 623 and 520 nm, respectively. The percentages of distribution of normal, early apoptosis, late apoptosis and necrosis cells were calculated and depicted in Fig. 4 and Table 1.
Assay of carbonyl content: Protein carbonyl was measured by reaction with 2,4-dinitrophenyl hydrazine (DNPH) following the method of Levine et al. (1990) similar to that done before in our laboratory (Mukhopadhyay and Chatterjee, 1994). To solution of pure Bovine Serum Albumin (BSA) 1 mg 100 μL, micro volume of carbonated soft drink was added on a time dependent manner and was directly subjected to carbonyl formation assay. The dose of carbonated soft drink was chosen as described above. In time dependant experiments, both concentration and volume of BSA and volume of carbonated soft drink were kept fixed at 1 mg 100 μL and 50 μL, respectively and the experimental time after addition of carbonated soft drink to pure protein were varied between 10-160 min under incubation. Formation of carbonyl was measured at 390 nm spectrophotometrically (Hitachi model, Fig. 5a). The results were expressed as n moles of phenylhydrazones formed per milligram protein using a molar extinction coefficient of 22,000.
Assay of bityrosine formation: To solution of pure Bovine Serum Albumin (BSA) 1 mg 100 μL, 50 μL of carbonated soft drink was added on a time dependent manner and was directly used to bityrosine formation estimation (Kaplan et al., 2003). Here to the solution, 20 mM HEPES buffer, pH 7.5 was added and the fluorescence measured at 325 nm excitation and 415 nm emission using a Hitachi fluorescence spectrophotometer, model F-7000. The results were expressed as n moles of per milligram protein using a molar extinction coefficient of 13600. Here the experiment was done by varying time between 1 to 140 min, as presented Fig. 5b.
Measurement of tryptophan loss: To solution 1 mg 100 μL of pure Bovine Serum Albumin (BSA), 50 μL of carbonated soft drink was added on a time dependent manner and was directly used to tryptophan loss assay. Tryptophan loss was measured at 280 nm excitation and 330-360 nm emission (Davies et al., 1987) using a Hitachi fluorescence spectrophotometer, model F-7000. The results were expressed as pico moles of per milligram protein using a molar extinction coefficient of 55,00. Here the time dependent study was done varying time period 0-160 min and the effect observed has been presented in Fig. 5c.
Statistical analysis: All values were expressed as Mean±Standard Deviation (SD) of 3 different sets of experiments, each set consisting of 3 controls and 15 experimental animals. Data were analyzed using origin 61 data analysis and graphing software. All statements were significant on based on a probability p≤0.05.
RESULTS
Determination of protein oxidation by carbonated soft drink in vivo by oxyblot assay: Extensive oxidation of liver tissue proteins (Fig. 1a, b) and serum proteins (Fig. 2a, b) were observed compared to control through Oxyblot assay when guinea pigs were orally administered carbonated soft drink for 30 days (Fig. 1a) and thereafter in a time-dependent manner (Fig. 1b). Besides oxidative damage low levels of serum protein degradations were also observed apparently due to the observed oxidative damage when animals were treated with carbonated soft drink for 60 days (Fig. 2b-silver stain). These observations may be explained due to generation of free radicals, under the experimental conditions as discussed later.
Determination of carbonated soft drink induced cellular apoptosis in guinea pigs in vivo through TUNEL assay: Tissue sections from sacrificed animals fed with carbonated soft drink for 60 days showed discernible apoptosis of liver (Fig. 3 ) and more significantly in kidney (Fig. 4) cells compared to control as determined by TUNEL assay reconfirming that the observed increase in cell death in the liver and kidney of the carbonated soft drink treated animals was apparently associated with the observed oxidative damage induced by the consumption of such drink, due to presence of high amount of sugar in such drink.
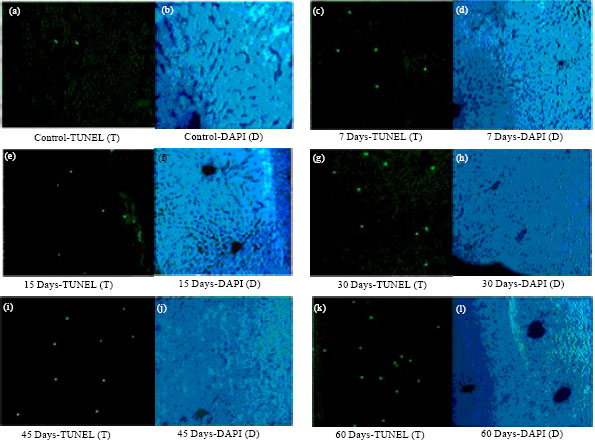 | |
| Fig. 3(a-l): | Detection of DNA strand breaks as cell death indicator by TUNEL assay in liver cells of guinea pigs orally administered carbonated soft drink (10x magnification), Uniform cross section, depict the DNA strand breaks in apoptotic cells as green fluorescent spots in the control and experimental liver tissue sections of the guinea pigs fed on water as control group while others were given carbonated soft drink in a time frame of 7, 15, 30, 45 and 60 days, respectively. T: TUNEL stained and DAPI stained by D: DAPI stained |
Determination of carbonated soft drink induced cellular apoptosis of guinea pig whole blood cells in vivo as measured by fluorescence-activated cell sorting (FACS) of annexin V and propidium iodide labeled cells: Whole blood cells collected from guinea pigs, which were orally administered with carbonated soft drink for definite experimental periods (as in legands) were subjected to flow cytometric analysis in order to assess any possible apoptosis of such cells. Results show that blood cells from treated guinea pigs undergo a dose-dependent increase in cellular apoptosis compared to control (Fig. 5). Early apoptosis appear between 30 to 60 days. It can be found Table 1 (4th column) that there is increase of apoptotic cell population percentage compared to control (0-day) from 0.04±0.005 to 30 days (6.75±0.42) which remained steady till 60th day (5.14±0.22).
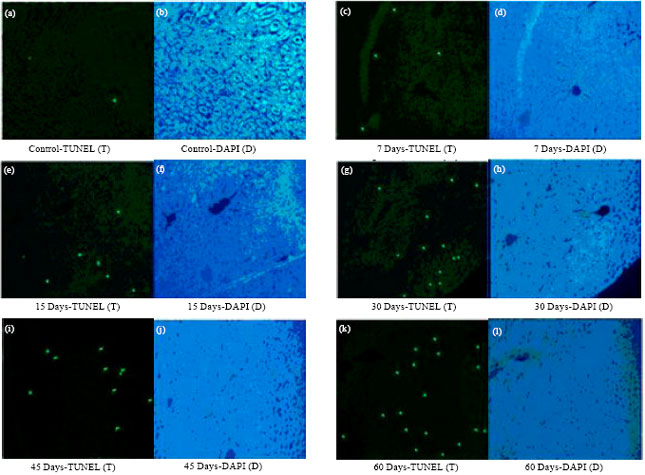 | |
| Fig. 4(a-l): | Detection of DNA strand breaks as cell death indicator by TUNEL assay in kidney cells of guinea pigs exposed to carbonated soft drink (10x magnification), Uniform cross section, depict the DNA strand breaks in apoptotic cells as green fluorescent spots in the control and experimental kidney tissue sections of the guinea pigs which have been given water while others were given carbonated soft drink for 7, 15, 30, 45 and 60 days, respectively, T: TUNEL stained and D: DAPI stained |
| Table 1: | Percentage distribution of normal, early apoptotic, late apoptotic and necrotic cells |
 | |
The annexin V-FITC/PI double staining assay was employed to classify blood cells in early apoptosis or late apoptosis stages, respectively.
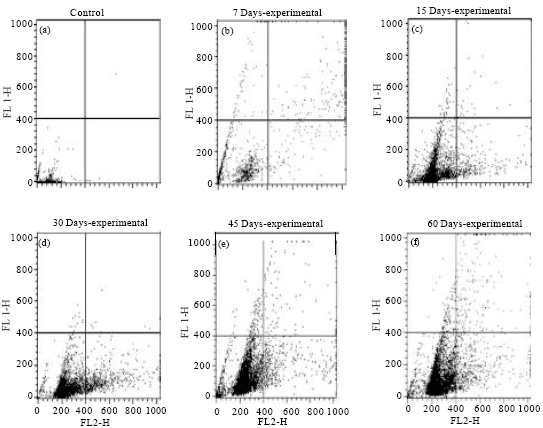 | |
| Fig. 5(a-f): | Determination of carbonated soft drink induced cellular apoptosis in guinea pig in vivo as measured by FACS of annexin V and propidium iodide labeled whole blood cells, Annexin V-Flurescein isothiocyanate/ Propidium Iodide (FITC/PI) stained blood cells was exposed to carbonated soft drink for a period of 0th, 7th, 15th, 30th, 45th and 60th day. Control: Blood cells after water intake carbonated soft drink, Experimental group: For a time frame of 7th, 15th, 30th, 45th and 60th day, respectively. The annexin V-FITC-/PI-population, Q3-lower left quadrant: Control cells, whereas annexin V-FITC+/PI- cells Q4-lower right quadrant: Early apoptosis, annexin V-FITC+/PI+ (Q2-upper right quadrant): late apoptosis and annexin V-FITC-/PI+ (Q1-upper left quadrant) as necrosis |
Figure 5 depicts the apoptotic profiles of blood cells isolated from the guinea pigs which were orally administered with carbonated soft drink for 0,7,15,30,45 and 60 days, respectively and clearly shows a time-dependent increase in early as well as late apoptosis in treated animals, with decrease at later stage.
Evaluation of in vitro oxidative potency of carbonated soft drink: This study was aimed to determine whether carbonated soft drink contains oxidants within it or these are generated by secondary metabolic pathway, when consumed, to cause oxidative damages. A pure protein Bovine Serum Albumin (BSA) was treated directly with fixed volume of carbonated soft drink and evaluated for oxidation due to this treatment. Protein carbonyl formation was found to increase in a time-dependent manner compared to control on addition of carbonated soft drink to such pure BSA solution (Fig. 6a).
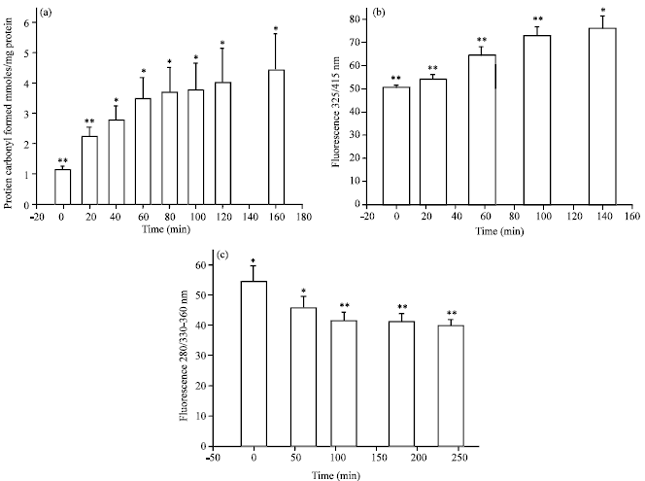 | |
| Fig. 6(a-c): | Time dependent in vitro assay of protein carbonyl formation by carbonated soft drink using Bovine Serum Albumin BSA, (a) Protien carbory formation, (b) Protien bityrosine formation and (c) Protien tryptophan loss was assessed by standand protocols protein |
Bityrosine formation (Fig. 6b) and tryptophan loss (Fig. 6c) which are both dependable indicators of protein oxidation also showed similar time-dependent increase compared to control during incubation of BSA with fresh carbonated soft drink. This confirmed that carbonated soft drink contains oxidants which are capable of direct oxidations of proteins.
DISCUSSION
India is a country with variety of population with different food habits and genetic variations. Many people may not have proper nutritional diets which are more prone in rural areas, but are fond of carbonated soft drink particularly due to hot and humid weather in most calendar months. The data on physiological and biochemical effects of carbonated soft drinks in animals are scanty (Amato et al., 1998; Milei et al., 2011; Jeroh et al., 2012; Awhin, 2012). Most data implicating such drinks to adverse health effects are largely based on epidemiological studies. The present study was thus aimed to obtain comprehensive data on the possible health hazards especially with respect to physiological and biochemical effects of such drink using carbonated soft drink, available in Indian market on suitable animal model, like guinea pigs, which represent humans in many metabolic aspects (Stith and Das, 1982) and cannot synthesize Vitamin C like humans and thus very suitable to study oxidative damages of tissue. Present study confirms that carbonated soft drink contain oxidants those are capable of oxidative modification of proteins which can partly explain the oxidative damage of liver, kidney and serum proteins as have been observed in this study in guinea pigs which were orally administered with such drink in measured doses.
In oxyblot assay, as described in the present study, extensive oxidation of liver tissue proteins and serum proteins have been observed under the experimental conditions in guinea pigs. Beside oxidative damage, protein degradation also takes place in serum, due to carbonated soft drink consumption. It has been reported that excess sugar consumption increases production of destructive free radicals (Mohanty et al., 2000). The high concentration of sugar as is present in carbonated soft drink under experimental condition as reported here may be responsible for such oxidation of tissue proteins in vivo due to generation of free radicals. Carbonated soft drink contains high sugar. Glucose has been shown to modify the amino group of proteins by a process of non-enzymatic glycation leading to potentially deleterious effects on structure and function of proteins (Cussimanio et al., 2003).
In TUNEL assay under the experimental conditions tissue sections of sacrificed animals fed with carbonated soft drink showed distinct apoptosis of liver and kidney. It has been reported that damages to proteins and DNA results from excess sugar consumption (Tappy and Le, 2010). The observed effect under experimental condition as reported here might be due to high amount of sugar present in carbonated soft drink.
In this study, it has been found that blood cells from experimental guinea pigs undergo a dose dependent increase in cellular apoptosis. The increase is though small but was noted in early apoptosis period between 30-60 days. The cellular apoptosis decreases at late stage with no significant necrosis of the cells. Early apoptosis appearance was noted between 30 to 60 days, compared to control. Reactive oxygen species may cause immediate apoptosis but due to feeding of subclinical dose of vitamin C (2 mg day-1) to animals during experimental period to prevent scurvy, such immediate apoptosis might not occurred. Alternatively, there may be cumulative depletion of other antioxidant present in the system between 30 to 60 days, which prevented immediate apoptosis. Certain polysaccharides have shown induced apoptosis of lymphoma cell line (Hattori et al., 2004). Observations of cellular apoptosis as reported here might also be due to high sugar content of such drink, in experimental animal like guinea pigs which are unable to synthesize vitamin C and are susceptible to oxidative damages and degradations.
In vitro studies as have been presented here show that carbonated soft drink directly furnish the oxidants, which are partly or wholly responsible for the physiological oxidative damage.
In India carbonated soft drink is very popular. But a large number of Indian populations do not get adequate nutrition from their daily diet and may suffer for insufficient antioxidant intake. This population could carry a risk of biochemical damages as observed here due to consumption of such drink in excess.
CONCLUSION
Carbonated soft drink has been used in this study, obtained from Indian market brings about extensive oxidation of liver and kidney tissue proteins and serum protein in vivo when fed to guinea pigs during the experimental period. Such consumption of carbonated soft drink make discernible apoptosis of liver and kidney tissues in guinea pigs under experimental conditions with early apoptosis of blood cells in a small proportion. In in vitro studies also direct oxidations of proteins have been noted.
REFERENCES
- Teerasong, S., S. Chan-Eam, K. Sereenonchai, N. Amornthammarong, N. Ratanawimarnwong and D. Nacapricha, 2010. A reagent-free SIA module for monitoring of sugar, color and dissolved CO2 content in soft drinks. Analytica Chimica Acta, 668: 47-53.
CrossRef - Malik, V.S., M.B. Schulze and F.B. Hu, 2006. Intake of sugar-sweetened beverages and weight gain: A systematic review. Am. J. Clin. Nutr., 84: 274-288.
PubMedDirect Link - Vartaman, L.R., M.B. Schwartz and K.O. Brownwell, 2007. Effects of soft drink consumption on nutrition and health: A systematic review and meta-analysis. Am. J. Public Health, 97: 667-675.
CrossRefPubMedDirect Link - Raben, A., T.H. Vasilaras, A.C. Moller and A. Astrup, 2002. Sucrose compared with artificial sweeteners: different effects on ad libitum food intake and body weight after 10 wk of supplementation in overweight subjects. Am. J. Clin. Nutr., 76: 721-729.
Direct Link - Heaney, R.P. and K. Rafferty, 2001. Carbonated beverages and urinary calcium excretion. Am. J. Clin. Nutr., 74: 343-347.
Direct Link - Tsimihodimos, V., V. Kakaidi and M. Elisaf, 2009. Cola-induced hypokalaemia: Pathophysiological mechanisms and clinical implications. Int. J. Clin. Pract., 63: 900-902.
CrossRef - Mohanty, P., W. Hamouda, R. Garg, A. Aljada, H. Ghanim and P. Dandona, 2000. Glucose challenge stimulates Reactive Oxygen Species (ROS) generation by leucocytes. J. Clin. Endocrinol. Metab, 85: 2970-2973.
CrossRefPubMedDirect Link - Tappy, L. and K.A. Le, 2010. Metabolic effects of fructose and the worldwide increase in obesity. Physiol. Rev., 90: 23-46.
CrossRefDirect Link - Glushakova, O., T. Kosugi, C. Roncal, W. Mu and M. Heinig et al., 2008. Fructose induces the inflammatory molecule ICAM-1 in endothelial cells. J. Am. Soc. Nephrol., 19: 1712-1720.
CrossRef - Al-Awwadi, N.A., C. Araiz, A. Bornet, S. Delbosc and J.P. Cristol et al., 2005. Extracts enriched in different polyphenolic families normalize increased cardiac NADPH oxidase expression while having differential effects on insulin resistance, hypertension and cardiac hypertrophy in high-fructose-fed rats. J. Agric. Food Chem., 53: 151-157.
CrossRef - Olcina, G.J., D. Munoz, R. Timon, M.J. Caballero, J.I. Maynar, A. Cordova and M. Maynar, 2006. Effect of caffeine on oxidative stress during maximum incremental exercise. J. Sports Sci. Med., 5: 621-628.
Direct Link - Ohnishi, M. and M.S. Raijaque, 2010. Dietary and genetic evidence for phosphate toxicity accelerating mammalian aging. FASEB J., 24: 3562-3571.
CrossRef - Kapicloglu, S., A.H. Baki, Y. Tekelioglu and K. Araz, 2000. The effect of cola consumption on oral mucosa in rats. Dis. Esophagus, 13: 69-71.
CrossRef - Imai, A., S. Ichigo, H. Takagi, K. Matsunami, K. Suzuki and A. Yamamoto, 2010. Effects of cola intake on fertility: A review. Health, 2: 997-1001.
CrossRefDirect Link - Amato, D., A. Maravilla, C. Montoya, O. Gaja, C. Revilla, R. Guerra and R. Paniagua, 1998. Acute effects of soft drink intake on calcium and phosphate metabolism in immature and adult rats. Rev. Invest. Clin., 50: 185-189.
PubMedDirect Link - Milei, J., L.M. Otero, L.H. Gomez, D.R. Grana, D. Suarez, F. Azzato and G. Ambrosio, 2011. Chronic cola drinking induces metabolic and cardiac alterations in rats. World J. Cardiol., 3: 111-116.
CrossRefDirect Link - Jeroh, E., E.P. Awhin, L. Osademe and E.I. Awire, 2012. Effect of carbonated drinks on the activities of alanine aminotransferase (ALT) and aspartate aminotransferase (AST) in serum and kidney in Rattus novergicus. Asian J. Biochem., 7 : 59-62.
CrossRefDirect Link - Awhin, E.P., 2012. The effect of coke consumption on lipid peroxidase and alkaline phosphatase activity in the serum and liver of Rattus novergicus. Cont. J. Med. Res., 6: 16-20.
Direct Link - Stith, I.E. and S.K. Das, 1982. Development of cholinephosphotransferase in guinea pig lung mitochondria and microsomes. Biochim. Biophys. Acta., 714: 250-256.
PubMed - Misra, A., R. Chattopadhyay, S. Banerjee, D.J. Chattopadhyay and I.B. Chatterjee, 2003. Black tea prevents cigarette smoke-induced oxidative damage of proteins in guinea pigs. J. Nutr., 133: 2622-2628.
Direct Link - Banerjee, S., R. Chattopadhyay, A. Ghosh, H. Koley and K. Panda et al., 2008. Cellular and molecular mechanisms of cigarette smoke-induced lung damage and prevention by vitamin C. J. Inflamm., Vol. 5.
CrossRefDirect Link - Das, A., N. Dey, A. Ghosh, T. Das and I.B. Chatterjee, 2011. NAD(P)H: Quinone oxidoreductase 1 deficiency conjoint with marginal vitamin C deficiency causes cigarette smoke induced myelodysplastic syndromes. PLoS One, Vol. 6, No. 5.
CrossRefDirect Link - Levine, R.L., D. Garland, C.N. Oliver, A. Amici and I. Climent et al., 1990. Determination of carbonyl content in oxidatively modified proteins. Meth. Enzymol., 186: 464-478.
CrossRefPubMedDirect Link - Mukhopadhyay, C.K and I.B. Chatterjee, 1994. NADPH-initiated cytochrome P450-mediated free metal ion-independent oxidative damage of microsomal proteins. Exclusive prevention by ascorbic acid. J. Biol. Chem., 269: 13390-13397.
PubMedDirect Link - Kaplan, P., E. Babusikova, J. Lehotsky and D. Dobrota, 2003. Free radical-induced protein modification and inhibition of Ca2+-ATPase of cardiac sarcoplasmic reticulum. Mol. Cell. Biochem., 248: 41-47.
CrossRef - Cussimanio, B.L., A.A. Booth, P. Todd, B.G. Hudson and R.G. Khalifah, 2003. Unusual susceptibility of heme proteins to damage by glucose during non-enzymatic glycation. Biophys. Chem., 105: 743-755.
CrossRef - Hattori, T.S., N. Komatsu, S. Shichijo and K. Itoh, 2004. Protein-bound polysaccharide K induced apoptosis of the human Burkitt lymphoma cell line, Namalwa. Biomed. Pharmacother., 58: 226-230.
CrossRefDirect Link