
P.C. Chikezie
Department of Biochemistry, Imo State University, Owerri, Imo State, Nigeria
LiveDNA: 234.10425
ORCID: 0000-0001-5066-724X
Asian Journal of Biochemistry
Year: 2011 | Volume: 6 | Issue: 1 | Page No.: 55-64







ABSTRACT
The aim of the present in vitro study was to ascertain the tendency of two quinoline (quinine and chloroquine phosphate) drugs to interfere with osmotic stability of three human erythrocyte genotypes, namely, HbAA, HbAS and HbSS. Spectrophotometric method was used for determination of the capacity of the erythrocyte genotypes to withstand osmotic stress in the presence of separate increasing concentrations (0.2, 0.4, 0.6 and 0.8 mg%) of the two antimalarials. The Mean Corpuscular Fragility (MCF) index of the three genotypes was in the order: HbAA<HbAS<HbSS irrespective of the malaria status of the blood donors. Whereas there was no significant difference (p>0.05) between the MCF values of non-malarious blood samples of HbAA and HbAS erythrocytes, values between HbAA and HbSS erythrocytes showed significant difference (p<0.05). In addition, parasitized erythrocytes exhibited significant (p<0.05) increased MCF values. Furthermore, at relative low experimental concentrations (approx<0.4 mg %) of the two drugs, parasitized erythrocytes and those of non-malarious human origin of HbAA and HbAS genotypes showed variable levels of stability. The HbSS erythrocytes did not exhibit osmotic stability within the range of experimental concentrations of the two drugs. The implications of these findings are discussed.
PDF Abstract XML References Citation
How to cite this article
P.C. Chikezie, 2011. Comparative in vitro Osmotic Stability of Three Human Erythrocyte Genotypes in the Presence of Quinine and Chloroquine Phosphate. Asian Journal of Biochemistry, 6: 55-64.
DOI: 10.3923/ajb.2011.55.64
URL: https://scialert.net/abstract/?doi=ajb.2011.55.64
DOI: 10.3923/ajb.2011.55.64
URL: https://scialert.net/abstract/?doi=ajb.2011.55.64
INTRODUCTION
Quinine acts primarily as blood schizontocide; it has little or no effect on sporozoites or preerythrocytic forms of malaria parasites. The alkaloid is gametocidal for P. vivax and P. malariae but not for P. falciparum (Tracy and Webster, 2001). Chloroquine is one of a large series of 4-aminoquinolines and closely resembles the obsolete 8-aminoquinoline antimalarials, primaquine and pentaquine. The drug contains the same side chain as quinacrine but differs from these antimalarials in having a quinoline instead of an acridine nucleus and lacking the methoxy moiety (Tracy and Webster, 2001). The drug has no activity against latent tissue forms of P. vivax or P. ovale and thus cannot cure infections with these species (Tracy and Webster, 2001).
Osmotic fragility is a measure of the resistance of the erythrocytes to lysis by osmotic stress (Oyewale and Ajibade, 1990). The test involves exposing erythrocyte to decreasing concentrations of isotonic buffered saline solution and measuring the level of haemolysis spectrophotometrically at room temperature. The test is generally useful to ascertain the level of stability and functionality of plasma membrane (Krogmeier et al., 1993). Compounds that can significantly promote membrane integrity or destabilization effect their actions by direct chemical contact with biomolecules that constitute the architectural structure of plasma membrane (Champe et al., 2005). Other compounds may act in such a way that the activity of certain erythrocyte redox enzymes, such as glutathione reductase (Becker et al., 2004; Forchetti et al., 2006), glutathione peroxidase (Mayes, 1983) and glucose-6-phosphate dehydrogenase (Mayes, 1983; Champe et al., 2005; Ojo et al., 2006), that are required for membrane integrity are compromised.
Early researchers have proposed the pre-incubation of donor-blood samples with antimalarials for the prevention and eradication of transfusion-induced malaria infection (Ali and Kadaru, 2005). While the exercise may exterminate the parasites, efforts should ensure that the drugs did not interfere with and/or distort erythrocyte integrity and functionality. Therefore, the efforts of the present study are; to investigate the variability of osmotic fragility amongst the three erythrocyte genotypes and the capacities of the two mentioned antimalarials to interfere with and/or distort membrane stability and integrity. The findings of the present in vitro study may provide a subset of useful preliminary data for effective, successful and safe utilization of these antimalarials for in vitro blood processing exercise.
MATERIALS AND METHODS
Anti-malarial drugs: The two antimalarials, quinine (BDH, UK) and chloroquine phosphate (May and Baker, Pharmaceutical Company, Nigeria Plc) were purchased on 15th July, 2009 from Cimpok Pharmaceuticals, Amakhohia, Owerri, Nigeria.
Collection of blood and preparation of erythrocyte samples: Five milliliters (5.0 mL) of human venous blood of HbAA, HbAS and HbSS genotypes obtained from subjects/volunteers by venipuncture was stored in EDTA anticoagulant tubes. Blood of HbSS genotype and malarious blood samples (density of asexual stage P. falciparum was between 20000 and 80000 parasite per μL of blood volume) were from patients attending clinics at the Federal Medical Center (FMC), Imo State University Teaching Hospital (IMSUTH), Orlu, St.John Clinic/Medical Diagnostic Laboratories, Avigram Medical Diagnostic Laboratories and Qualitech Medical Diagnostic Laboratories. These centers are located in Owerri, Imo State, Nigeria.
The erythrocytes were washed by centrifugation method as described by Tsakiris et al. (2005). Within 2 h of collection of blood samples, portions of 1.0 mL of the samples were introduced into centrifuge test tubes containing 3.0 mL of buffer solution pH = 7.4: 250 mM tris (hydroxyl methyl) amino ethane-HCl(Tris-HCl)/140 mM NaCl/I.0 mM MgCl2/10 mM glucose). The suspension was centrifuged at 1200x g for 10 min to separate the erythrocytes from the liquid phase. After centrifugation, the supernatant was carefully withdrawn with a pastuer pipette and decanted. The sediment constituted harvested erythrocytes. The erythrocytes were re-suspended in the buffer and washed three times by similar centrifugation technique. The erythrocytes were finally suspended in 1.0 mL of this buffer solution.
Determination of erythrocyte osmotic fragility: Osmotic fragility of human three erythrocyte genotypes, HbAA, HbAS and HbSS was determined by a measure of haemoglobin released from erythrocytes when placed in an environment containing serial dilutions of Phosphate Buffer Saline (PBS) solution as described by Oyewale (1993), with minor modifications (Mafuvadze et al., 2008).
Control analysis: Twenty microliters portion of erythrocytes suspended in 1.0 mL buffer solution: pH = 7.4 (Tris HCl/140 Mm NaCl/1.0 mM MgCl2/10.0 mM glucose), was added to test tube containing 5.0 mL of PBS solution, pH = 7.4 - {NaCl (9.0 g)/Na2HPO4.2H2O (1.71 g)/ NaH2PO4.2H2PO4.2H2O (2.43 g) per 1litre of distilled water}, of serial concentrations in the order of 0.9, 0.7, 0.6, 0.4, 0.3 and 0.2 g/100 mL. The seventh test tube contained distilled water. The test tubes were allowed to stand for 30 min at room temperature (24°C). Subsequently, the contents of test tubes were centrifuged at 1200 g for 10 min. The supernatant was decanted and haemoglobin content determined spectrophotometrically at λmax= 540 nm using PBS (0.9 g/100 mL) solution as blank. Haemolysis in each test tube was expressed as a percentage, taking as 100% the maximum value of absorbance of the test tube that contained erythrocytes suspended in distilled water (0.0 g/100 mL).
Test analysis: A 0.5 mL of the human three erythrocyte genotypes were incubated for 30 min, at room temperature (24°C), in 1.0 mL buffer solution pH = 7.4 (Tris HCl/140 mM NaCl/1.0 mM MgCl2/10.0 mM glucose), in the presence of 0.5 mL of increasing concentrations (0.2, 0.4, 0.6 and 0.8 mg %) of the separate 2 antimalarial drugs. A portion of 20 μL of erythrocytes suspension was used for determination of osmotic fragility.
Evaluation of percentage haemolysis and stabilization of erythrocytes: The quotient of absorbance of the content of each test tube (1st-6th) and the seventh test tube was multiplied by a factor of 100. The range of values represented the percentage of erythrocyte lysis at each corresponding PBS concentration (0.9-0.2 g/100 mL).
Where:
| 0.DA | = | Absorbance of test tube (1st-6th) supernatant |
| 0.DB | = | Absorbance of 7th test tube supernatant |
The corresponding concentration of PBS solution that caused 50% lysis of erythrocytes defined the MCF index (Dewey et al., 1982; Krogmeier et al., 1993). The erythrocyte osmotic fragility curve; the plot of percentage of erythrocyte lysis versus concentrations of PBS solution was used to obtain the MCF values.
The relative capacity of the five antimalarial drugs to stabilize or destabilize erythrocyte membrane was evaluated as percentage of the quotient of the difference between MCF values of test and control samples to the control sample (Parpart et al., 1947; Chikezie, 2007). Thus:
Statistical analyses: The experiments were designed in a completely randomized method and data collected were analyzed by the analysis of variance procedure while treatment means were separated by the Least Significance Difference (LSD) incorporated in the Statistical Analysis System, package of 9.1 versions (SAS, 2006).
RESULTS
The MCF index represented and interpreted level of erythrocyte membrane stability. The mean (±SD) MCF values of the three-erythrocyte genotypes (HbAA, HbAS and HbSS) of blood samples obtained from non-malarious and malarious blood donors is presented in Table 1.
| Table 1: | Osmotic fragility: mean corpuscular fragility (MCF) index of erythrocyte of non-malarious and malarious blood donors: |
 | |
| *M and NM = No. of malarious and non-malaroius blood samples respectively. *Means in the column with the same letter are not significantly different at p<0.05 according to LSD | |
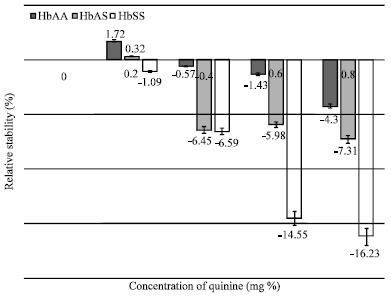 | |
| Fig. 1: | Comparative in vitro stability of three erythrocyte genotypes (HbAA; n = 25, HbAS; n = 25 and HbSS; n = 20) of non-malarious human blood samples in the presence of quinine |
The mean (±SD) MCF values of the three genotypes were in the order: HbAA<HbAS<HbSS irrespective of the malarial status of the blood donors. Whereas there was no significant difference (p>0.05) between the MCF values of non-malarious blood samples of HbAA and HbAS erythrocytes, the values between HbAA and HbSS erythrocytes exhibited significant difference (p<0.05).
In addition, parasitized erythrocytes exhibited significantly (p<0.05) increased MCF values. This was an obvious reflection of higher fragility index of these P. falciparum infected erythrocytes. Specifically, whereas MCF values of erythrocytes obtained from non-malarious blood donors ranged between 0.352+/-0.06 and 0.422+/-1.49 g/100 mL, parasitized erythrocytes presented values within the range of 0.445+/-1.01 and 0.497+/-1.11 g/100 mL.
Non-malarious blood samples (HbAA Genotype): Within the concentration range of (0.6-0.8) mg percentage, Quinine promoted erythrocyte membrane destabilization. However, at relatively low concentration, specifically at 0.2 mgpercentage, the drug exhibited membrane stabilizing effect ([Quinine] = 0.2 mg percentage; MCF = 0.343±0.05 g/100 mL; stability = 2.28%; p<0.05) (Fig. 1).
The results also showed Chloroquine Phosphate as agents of erythrocyte stabilization. Chloroquine Phosphate at 0.2 mg percentage apparently promoted membrane stability (MCF = 0.307+/-0.03 g/100 mL; stability = 12.54%; p<0.05). However, the capacity of the drug to stabilize the erythrocyte membrane diminished in a concentration dependent manner (Fig. 3).
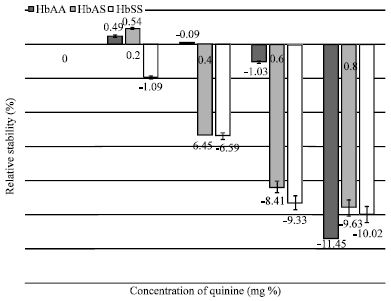 | |
| Fig. 2: | Comparative in vitro stability of three erythrocyte genotypes (HbAA; n = 24, HbAS; n = 25 and HbSS; n = 12) of malarious human blood samples in the presence of quinine |
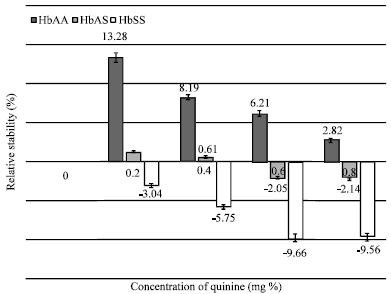 | |
| Fig. 3: | Comparative in vitro stability of three erythrocyte genotypes (HbAA; n = 25, HbAS; n = 25 and HbSS; n = 20) of non-malarious human blood samples in the presence of chloroquine phosphate |
Non-malarious blood samples (HbAS Genotype): A 0.2 mg percentage of Quinine and between ranges of 0.2-0.4 mg percentage of Chloroquine Phosphate promoted erythrocyte stabilization. The capacities of these drugs at the specified concentrations to stabilized the erythrocytes was not significantly different (p>0.05) from the control/reference blood samples. Corresponding higher concentrations of these drugs engendered membrane destabilization. However, Fig. 3 showed that 0.6 mg percentage concentration of Chloroquine Phosphate did not significantly (p>0.05) promote erythrocyte membrane distabilization. Malarious Blood Samples (HbAA Genotype): Quinine at relatively low concentrations, specifically at 0.2 mg percentage and 0.4 mg percentage, stabilized the erythrocytes at levels of 0.49 and 0.09%, respectively. The capacity of the drug to stabilize erythrocytes was however not significantly different (p>0.05) compared to the control samples.
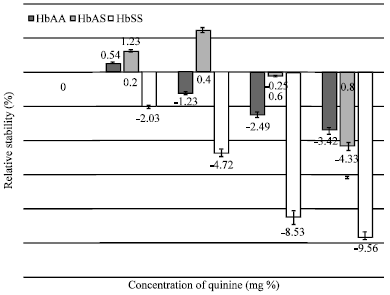 | |
| Fig. 4: | Comparative in vitro stability of three erythrocyte genotypes (HbAA; n = 24, HbAS; n = 25 and HbSS; n = 12) of malarious human blood samples in the presence of chloroquine phosphate |
Whereas Chloroquine Phosphate at 0.2mgpercentage stabilized the erythrocytes (stabilization = 0.54%), (Fig. 4) higher concentrations disrupted erythrocyte membrane integrity.
Malarious blood samples (HbAS genotype): Figure 2 showed that Quinine at 0.2 mg percentage stabilized erythrocytes by 0.94%. Likewise, 0.2 and 0.4 mg percentage concentrations of Chloroquine Phosphate caused erythrocyte stabilization by 1.23 and 2.49%, respectively. Further corresponding increase in the concentrations of these two drugs promoted membrane destabilization. Concentration of Chloroquine Phosphate at 0.6 mg percentage did not show significant (p>0.05) capacity to destabilize the erythrocytes. HbSS Blood Samples: In concentration dependent manner, the four experimental concentrations of Quinine and Chloroquine Phosphate engendered increasing erythrocyte osmotic fragility.
A general overview of the results showed that the two drugs exhibited similar patterns in influencing erythrocyte membrane integrity within the four experimental concentrations. However, Quinine caused higher overall destabilizing effect on the three-erythrocyte genotypes compared to Chloroquine phosphate.
DISCUSSION
From comparative investigations, the results presented in Table 1 showed that human erythrocyte of HbSS genotype exhibited the least stability that was in the order HbAA>HbAS>HbSS. In agreement with these results, Dewey et al. (1982) asserted that differences in erythrocyte osmotic fragility are under the control of the individual genotype of the erythrocytes. Thus, during erythrocyte formation, any of the number of erythrocyte properties such as membrane structure, cell shape or internal salt balance, responsible for variant erythrocyte behavior occurred according to the dictate of genetic makeup of corresponding erythrocytes (Dewey et al., 1982). From a similar perspective, it is probable that variations in some physicochemical properties and oxidant levels of the three erythrocyte genotypes contributed to the differences in mechanical stabilities and capacities of the erythrocytes to withstand osmotic stress (Senturk et al., 2001; Richards et al., 2007; Chikezie et al., 2009). Decreasing stability and deformability of HbSS erythrocytes demonstrated cross-linking of spectrin, a erythrocyte membrane protein of major structural importance, caused by oxidative damage of sulphydryl groups (Lubin and Chiu, 1982; Palek and Liu, 1979). In addition, oxidized haemoglobin in the form of Heinz bodies attached to the interior of membrane surface is a common phenomenon in sickle cell disease. This property causes redistribution of major membrane components such as anion channel, ankyrin and glycophorin (Chiu and Lubin, 1989). These redistributions enhance IgG binding and reduced deformability and fragility (Chiu and Lubin, 1989).
Moreover, increasing evidence suggest that in vivo lipid peroxidation may be an important factor in sickle cell anemia (Stone et al., 1990). Sickle erythrocytes and their membranes are susceptible to endogenous free radical-mediated oxidative damage that correlates with the proportion of irreversibly sickled cell (Rice-Evans et al., 1986). In agreement with these lines of reasoning, Tamer et al. (2000) reported that higher superoxide generation in human HbSS erythrocytes was associated with increased tendency of diminished mechanical and osmotic stability compared with human HbAA erythrocytes. Furthermore, erythrocytes generate superoxide species under normal physiological conditions, but drastically increase in sickle cell disease. Unstable hemoglobin produced under this condition generates free radicals and further induce erythrocyte hemolysis (Chan, 1996). Therefore, accumulation of oxidant contributes to accelerated damage of sickle erythrocyte membranes and senescence of these cells. From another perspective, comparative osmotic stability of human erythrocytes showed connection with the relative tendency of the cells to retain more sodium ion (Na+) intracellularly with a concomitant loss of potassium ion (K+) (Dunham and Hoffmann, 1971).
Over four decades ago, Herman (1969) reported that osmotic fragility of normal duck erythrocytes significantly increased after exposure in vitro to cell-free extracts of P. lophurae or P. lophurae-infected duck erythrocytes. He further averred that plasma from infected ducklings could also produce an increased osmotic fragility of normal duck erythrocytes in vitro. The present investigations have also demonstrated that the osmotic fragility index of parasitized cells in P. falciparum infections was significantly increased. These observations also, were in agreement with the reports of Fogel et al. (1966). They demonstrated that the osmotic fragility of parasitized cells in Plasmodium berghei, P. knowlesi, P. gallinaceum and P. falciparum infections was significantly increased. In addition, their findings showed that the increased fragility of erythrocytes was not solely limited to the parasite-containing cells. In a closely related study, Dubey et al. (2004) observed that P. falciparum infected erythrocytes exhibited gradual increase in osmotic fragility as the parasites mature from ring to schizont stage. Furthermore, Clark et al. (1991) proposed that Nitric oxide production increase in any generalized infection particularly in acute malaria. Higher levels of nitric oxide produce poor deformability of erythrocytes by inhibiting Na+/K+ ATPase in the erythrocyte membrane and oxidizing the membrane lipids through generation of peroxiynitrate.
Previous in vitro studies by Soforawa (1975), Dean and Schechter (1978), Uwakwe and Ezeh (2000) and Ali and Kadaru (2005) reported the capability of xenobiotics to interfere with erythrocyte membrane integrity and stability. In addition, many authors have cited a large number of drugs that cause alterations on the shape and physiology of the erythrocytes (Ammus and Yunis, 1989; Braga et al., 2000). Evidence that drugs can interfere with osmotic resilience of erythrocytes have been demonstrated with various natural products (Chikezie and Ibegbulem, 2004; Chikezie, 2007; De Souza Fontes et al., 2007). In concord with these reports, our present studies have shown the two antimalarial drugs interfered with erythrocyte membrane stability.
The similarity in the pattern of membrane stabilization/destabilization in the presence of the two drugs suggested common mode of action on erythrocyte integrity. Numerous membrane destabilizing agents may act by direct interaction with architectural membrane proteins and enzymes, thereby modifying their structure/function relationship that is necessary and required for membrane integrity (Bazzoni and Rasia, 1998). Chloroquine and Quinine have been described to act by modifying certain protozoan membrane proteins (Tracy and Webster, 2001).
CONCLUSION
One of the properties of an ideal antimalarial drug, applicable for in vitro blood processing procedure for the prevention of transfusion induced malarial is one that exhibit minimum or insignificant destabilizing effect on erythrocyte membrane. The present study showed the critical concentrations of Quinine and Chloroquine phosphate that engendered membrane destabilization of three human erythrocyte genotypes. However, further investigation is necessary to ascertain whether concentrations below the corresponding critical values of the two antimalarials are capable to eradicate the parasite in vitro. Finally, the erythrocyte genotype (HbSS) did not exhibit stability within the range of experimental concentrations of the two antimalarials. Therefore, the sickle cell erythrocyte may not be suitable for in vitro blood processing procedure.
REFERENCES
- Ali, M.S.M. and A.G.M. Kadaru, 2005. In vitro processing of donor blood with sulphadoxine/pyrimethamine for eradication of transfusion induced malaria. Am. J. Trop. Med. Hyg., 73: 1119-1123.
Direct Link - Becker, K., L. Tilley, J.L. Vennerstrom, D. Roberts, S. Rogerson and H. Ginsburg, 2004. Oxidative stress in malaria parasite infected-erythrocytes: Host-parasite interactions. Int. J. Parasitol., 34: 163-189.
CrossRefDirect Link - Bazzoni, G. and M. Rasia, 1998. Effects of an amphipathic drug on the rheological properties of the cell membrane. Blood Cells Mol. Dis., 24: 552-559.
Direct Link - Braga, A.C.S., M.B.N. Oliveira, G.D. Feliciano, I.W. Reninger, J.F. Oliveira, C.R. Silva and M. Bernardo-Filho, 2000. The effect of drugs on the labeling of blood elements with technetium-99m. Curr. Pharm. Des., 6: 1179-1191.
CrossRefDirect Link - Mafuvadze, B., M. Nyanungo, H. Saina, B. Gorejena, T. Mashayamombe and K.H. Erlwanger, 2008. Deprivation of drinking water for up to 48 hours does not affect the osmotic fragility of erythrocytes from captive helmeted guinea fowl (Numida meleagris). Int. J. Poult. Sci., 7: 59-63.
CrossRefDirect Link - Oyewale, J.O., 1993. Effect of storage of blood on the osmotic fragility of mammalian erythrocytes. J. Vet. Med., 40: 258-264.
PubMedDirect Link - Palek, J. and S.C. Liu, 1979. Dependence of spectrin organization in erythrocyte membranes on cell metabolism: implications for control of erythrocyte shape, deformability and surface area. Semin Hematol., 16: 75-93.
PubMed - Parpart, A.K., P.B. Lorenz, E.R. Parpart, J.R. Gregg and A.M. Chase, 1947. The osmotic resistance (fragility) of human red cells. J. Clin. Invest., 26: 636-640.
CrossRefDirect Link - Rice-Evans, C., S.C. Omorphos and E. Baysal, 1986. Sickle cell membranes and oxidative damage. Biochem. J., 237: 265-269.
CrossRefPubMedDirect Link - Senturk, U.K., F. Gunduz, O. Kuru, M.R. Aktekin and D. Kipmen et al., 2001. Exercise-induced oxidative stress affects erythrocytes in sedentary rats but not exercise-trained rats. J. Applied Physiol., 91: 1999-2004.
Direct Link - Tsakiris, S., A. Giannoulia-Karantana, I. Simintzi and K.H. Schulpis, 2005. The effect of aspartame metabolites on human erythrocyte membrane acetylcholinesterase activity. Pharmacol. Res., 53: 1-5.
CrossRefPubMedDirect Link - Stone, W.L., P.H. Payne and F.O. Adebonojo, 1990. Plasma vitamin E and low plasma lipoprotein levels in sickled cell anemia patients. J. Assoc. Acad. Minor Phys., 1: 12-16.
PubMed