
Nivedita Jaiswal
Department of Biochemistry, Faculty of Sciences, Banaras Hindu University, Varanasi-221005, U.P., India
Om Prakash
Department of Biochemistry, Faculty of Sciences, Banaras Hindu University, Varanasi-221005, U.P., India
Asian Journal of Biochemistry
Year: 2011 | Volume: 6 | Issue: 4 | Page No.: 337-346







ABSTRACT
The detailed kinetic study of the immobilized enzyme is usually important and pertinent for the better management of the enzyme preparation. Therefore, the suitable comparison of the various kinetic parameters of the immobilized α-amylase (on gelatin) with that of the soluble enzyme has been conducted. The purified α-amylase was immobilized on gelatin using glutaraldehyde as an organic hardener with an optimum immobilization of 82.5% followed by characterization with respect to pH, temperature, substrate concentration, effect of thiols and thiol inhibitors. The optimum pH shifted from 5.5 to 7.0 i.e., towards the basic side upon immobilization. The optimum temperature of gelatin immobilized α-amylase was 85°C as compared to soluble enzyme (75°C) showing an improvement in thermal stability. A decrease in Michaelis constant, Km from 2.427 to 1.870 mg mL-1 upon immobilization was observed. An increase in activity with all the thiols was observed. No marked inhibition with the thiol inhibitors (similar to the soluble enzyme) is an indication of the fact that there are no free thiol or sulfhydryl groups in soybean α-amylase which are necessary for catalysis. The reutilization capacity of the immobilized enzyme was up to seven cycles. The potential utility of the gelatin immobilized enzyme in removal of starch stain from cloths by various commercially available detergents and sodium lauryl sulphate was tested. The immobilized α-amylase could be considered as a potential candidate for use as a cleaning additive in detergents in order to facilitate the removal of starch stains due to which it may find potential application in laundry detergents.
PDF Abstract XML References Citation
Received: January 19, 2011;
Accepted: April 21, 2011;
Published: June 18, 2011
How to cite this article
Nivedita Jaiswal and Om Prakash, 2011. Immobilization of Soybean α-amylase on Gelatin and its Application as a Detergent Additive. Asian Journal of Biochemistry, 6: 337-346.
DOI: 10.3923/ajb.2011.337.346
URL: https://scialert.net/abstract/?doi=ajb.2011.337.346
DOI: 10.3923/ajb.2011.337.346
URL: https://scialert.net/abstract/?doi=ajb.2011.337.346
INTRODUCTION
The immobilization of enzyme on insoluble supports has been a topic of active research in enzyme technology and is essential for their application to individual processes, for example in food technology, biotechnology, biomedicine and analytical chemistry. Enzyme immobilization is preferred to enhance enzyme stability, multiple and repetitive use and to remove the enzyme from the reaction mixture easily (Hasirci et al., 2006; Pandya et al., 2005; Altun and Cetinus, 2007). Since amylases have many applications in the food and fermentation industries, it is not surprising that several methods have been developed for the preparation of immobilized α-amylase, each having its own advantages and disadvantages specific to the methods. Some examples of different supports used for α-amylase immobilization are: glass beads and glutaraldehyde fixation (De Cordt et al., 1994), polymeric microspheres (Tumturk et al., 2000), coconut fiber (Dey et al., 2002), cellulose fibers from bagasse (Varavinit et al., 2002), functionalized silica (Lim et al., 2003), Cu2+-chelated poly (ethylene glycol dimethacrylate-n-vinylimidaole) (Kara et al., 2005), poly(dimmer acid-co-alkyl polyamine) particles (Hasirci et al., 2006), UV-curable polymers (Kahraman et al., 2006), sodium alginate, chitin, sephadex and dowex (El-Banna et al., 2007), functionalized glass beads (Kahraman et al., 2007), calcium alginate (Ertan et al., 2007), adsorption on zirconia (Reshmi et al., 2007), cellulose fiber (Egwim and Oloyede, 2008), cyclic carbonate (Turunc et al., 2009), polyaniline (Pascoal et al., 2010), coconut coir (Borgio, 2011). The several procedures for the immobilization of an enzyme involve: (1) entrapment of the enzyme molecules in polymeric structures, (2) physical adsorption of the enzyme onto a solid support, (3) covalent binding to an activated support, (4) copolymerization of the enzyme molecules with the polymers and (5) cross linking between the enzyme molecules. Among these immobilization methods, covalent binding is usually advantageous as it: a) increases the thermal and storage stability of the enzyme, b) prevents leakage and c) decreases diffusion problems of the substrate and products in the reaction medium (Hasirci et al., 2006). Recently, α-amylase has been covalently immobilized onto poly (hydroxyethylmethacrylate) and poly (methyl methacrylate-acrylic acid) microspheres and zirconium membranes (Arica et al., 1995; Sun et al., 1999; Tien and Chiang, 1999).
α-amylase (EC 3.2.1.1; 1,4-α-d-glucanohydrolase) is one among the various starch hydrolyzing enzymes which is of considerable interest in terms of industrial applicability. It is particularly employed in starch processing industries for the hydrolysis of polysaccharides such as starch into simpler sugar constituents (Ikram-ul-Haq et al., 2002). It catalyzes the hydrolysis of glucosidic linkages as starch to produce smaller oligosaccharides. This property of α-amylase is exploited for its practical use in removal of starch stain from cloths as starch is a major polysaccharide used as a food ingredient such as pasta, chocolate, baby food, sauce and gravy. The removal of starch from surfaces is important as starch can be an attractant for dust and dirt as well (Hmidet et al., 2009). Usually, the enzymes used in detergents are unstable and are at the risk of being attacked and inhibited by the detergent action. Immobilization may be found as a better solution to this which not only protects the enzyme from inhibition by various detergent components but also facilitates its reuse (Maliks et al., 2000). Various literatures are available showing the washing efficiency of α-amylases from various sources (Hmidet et al., 2009; Rani et al., 2007; Dhingra et al., 2006; Mitidieri et al., 2006).
In the present study, effects of various immobilization parameters in order to obtain maximum yield of immobilized α-amylase was studied. The immobilized enzyme was characterized with respect to pH, temperature, substrate concentration and effect of thiols and thiol inhibitors. Besides, its application as a detergent additive in order to remove starch stain from cloth was checked.
MATERIALS AND METHODS
Materials: Soybean seeds were procured from the local market. Sodium alginate, Maltose, Gelatin and 3,5-dinitrosalicylic acid (DNS) was purchased from Sigma Chemicals Co. (St. Loius, MO, USA). Soluble starch was from Qualigens Fine Chemicals, Mumbai. All other reagents were analytical grade chemicals either from BDH or E. Merck, India. The study was conducted during January 2010 to December 2010.
Isolation and purification of α-amylase: α-amylase was isolated from soybean (Glycine max) seeds, procured from the local market and was purified to electrophoretic homogeneity as described Prakash and Jaiswal (2010).
Immobilization of α-amylase on gelatin: Gelatin solution of different concentrations was prepared by dissolving gelatin in distilled water by heating at 50°C and thereafter cooling to room temperature. One milliliter of the purified enzyme (0.44 mg protein) was added to 9.0 mL gelatin solution with thorough mixing. The suspension was casted on preassembled glass plates and when the suspension jellified, 10 mL of 10% glutaraldehyde solution was added and left for complete hardening. The gelatin was then cut into small beads (5x5 mm), washed thoroughly with distilled water to remove any enzyme attached to the surface of beads and stored in 25 mM sodium acetate buffer (pH 5.5) at 4°C.
α-amylase assay: α-amylase activity was estimated following the method as described by Bernfeld (1955).
Protein estimation: Protein was estimated by the method of Lowry et al. (1951) with Folin-ciocalteau reagent calibrated with crystalline bovine serum albumin.
Steady-state kinetics and storage stability of immobilized enzyme: The optimum pH for the immobilized α-amylase was determined by varying the pH of assay buffer (Tris-acetate) from 4.0-9.0. The enzymatic activity was determined by the method described for enzyme assay. The optimal temperature was studied by varying the temperature of immobilized enzyme in assay buffer from 0-100°C. Km was determined by Lineweaver and Burk (1934) with variation in the substrate (starch) from 5 to 40 mg mL-1. In order to test the reusability of the immobilized α-amylase, the activity in the beads was assayed several times for the hydrolysis of soluble starch. The activities of the first batch were taken to be 100%. After each amylase activity assay of the beads, they were removed, washed thoroughly with distilled water at 4°C. Then, the beads were reassayed for α-amylase activity and the same process was repeated till the seventh use.
Effect of thiols and thiol inhibitors: The effect of different thiols and thiol inhibitors on enzyme activity was observed with the varying concentration of these reagents in the reaction mixture and then assaying the enzyme activity. The activity assayed in the absence of these reagents was taken as 100% and residual activity determined in their presence was calculated.
Application of immobilized α-amylase: Clean cotton cloth pieces of size 5x5 cm were stained with 0.2 mL aqueous starch solution. Stock solutions (2.0 g L-1) of commercially available detergents like Surf Excel, Ariel, Rin, Tide, Ghadi and sodium lauryl sulphate (a major component of detergent) were prepared in distilled water. Anthrone reagent (0.02%) was prepared in chilled concentrated sulphuric acid. The stained cloth pieces were subjected to wash treatments in separate flasks: (1) in distilled water only (2) in detergent only (3) in detergent and immobilized α-amylase. Each flask was incubated at 40°C for 20 min under continuous agitation (150 rpm). After incubation, the washing performance in each case was determined. The cloth piece from each flask was taken out and the volume of washout so left was made up to 25 mL with distilled water. A 1.0 mL of these washouts was mixed with 2 mL of anthrone reagent in tubes and then were placed in boiling water bath for 10 min and cooled to room temperature. The color obtained was determined spectrophotometrically at 630 nm. A standard curve was prepared by taking varying concentration of glucose in the range of 0-100 μg. The amount of glucose produced was extrapolated from the standard curve. The value of glucose was multiplied by 0.9 to convert glucose value to starch.
Statistical analysis: Data were analyzed using SPSS (version 16.0) package on a microcomputer. All values expressed were the means for three replicate experiments carried out with a fresh batch of purified enzyme.
RESULTS AND DISCUSSION
Immobilization of α-amylase on gelatin: In the present study, the enzyme was immobilized on gelatin matrix and covalently crosslinked by the organic hardener glutaraldehyde. The optimization of immobilization of α-amylase in gelatin was done by varying the concentration of gelatin from 5 to 30% (w/v). As it is evident from the Table 1, maximum immobilization was achieved at 20% gelatin concentration. The results revealed that at low gelatin concentration, unstable and fragile gelatin beads were obtained which led to poor immobilization which may be due to the larger pore size in the gel which probably caused leaching of the enzyme. At concentrations above 20%, the percent immobilization was low which may be the result of steric hindrance due to high concentration of gelatin.
The glutaraldehyde may be said to act as a hardening agent as it was not easy to handle the matrix (with the enzyme entrapped within) in its absence. Moreover, the glutaraldehyde concentration above 10% led to a decline in percent immobilization which again may be the result of the steric hindrance caused by the presence of the glutaraldehyde which may have affected the accessibility of the substrate to amylase (Table1). Thus, the optimum immobilization of 82.5% was observed with 20% gelatin and 10% glutaraldehyde concentration.
Steady-state kinetics: An enzyme in solution can have altered pH optima upon immobilization. Depending upon the surface and residual charges on the solid matrix and the nature of the enzyme bound, the pH value in the immediate vicinity of the enzyme molecule may change and thus cause a shift in the pH of the enzyme activity.
The effect of pH on the activity of gelatin immobilized amylase is shown in Fig. 1. The pH optima of soluble and immobilized amylase were 5.5 (Prakash and Jaiswal, 2010) and 7.0, respectively, i.e., a shift of 1.5 Units towards the basic side was resulted from immobilization of α-amylase on gelatin. A similar trend of shift in pH of 2.4 U towards the basic side (from 5.6 to 7.0) was observed by Tripathi et al. (2007) from immobilization of α-amylase on both chitosan and amberlite.
| Table 1: | Percent immobilization with varying gelatin concentration |
 | |
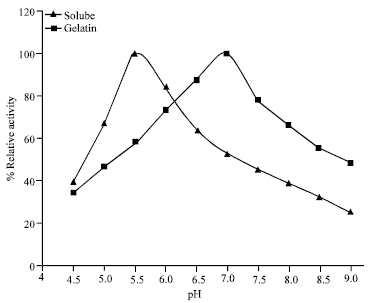 | |
| Fig. 1: | Effect of pH on soluble and gelatin immobilized α-amylase activity |
Similarly, El-Shora and Youssef (2008) also found a shift of pH optimum from 7.5 to 8.0 on immobilization of glutamate dehydrogenase on gelatin beads via cross-linking with glutaraldehyde. No change in optimum pH was reported in case of commercial α-amylase (Bacillus sp.) immobilized onto pHEMA microspheres and to a composite temperature sensitive membrane, respectively (Tien and Chiang, 1999; Chen et al., 1998). A shift towards acidic region has been observed when α-amylase was immobilized on poly(methylacrylate-acrylic acid) microspheres (Aksoy et al., 1998) and on zirconium dynamic membranes (Tien and Chiang, 1999). The alteration in the pH optimum of the enzyme in solution and upon immobilization to a solid matrix is largely dependent on the residual surface charges on the solid matrix and the nature of the bound enzyme. The effect of temperature on the activity of soluble and immobilized enzyme is shown in Fig. 2. The soluble enzyme exhibited a temperature optimum of 75°C (Prakash and Jaiswal, 2010) and this value shifted to 85°C for the immobilized enzyme. This increase in the temperature optima may be due to the conformational integrity of the enzyme structure upon covalent binding to the support material (Tien and Chiang, 1999; Bayramoglu et al., 1992; Demircioglu et al., 1995; Lee et al., 1993).
On comparison of the kinetic constants of soluble and immobilized amylase, a decrease in Km [from 2.427 (soluble) to 1.87 (gelatin immobilized)] was observed (data not shown). A similar decrease in Km was observed upon immobilization of α-amylase onto cyclic carbonate (Turunc et al., 2009). The Km value is known as the affinity of the enzymes to substrates (Cosulich et al., 2000; Park et al., 2005) and the lower value of Km emphasize the higher affinity between enzymes and substrates. The present results show that affinity of the enzyme to its substrate was increased by immobilization.
The reusability of the gelatin immobilized enzyme was upto seven cycles. Almost 90% activity was retained upto three cycles, but with subsequent runs, there was a decline in the activity of the immobilized enzyme. The activity loss upon reuse could be due to weakening in the strength of binding between the matrix and enzyme on repeated use and hence the enzyme might leach out from the matrix, therefore yielding a loss in activity. Besides, the frequent encountering of the substrate into the same active site might distort it which would dwindle the catalytic efficiency either partially or fully.
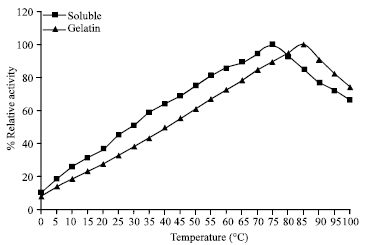 | |
| Fig. 2: | Effect of temperature on soluble and gelatin immobilized α-amylase activity |
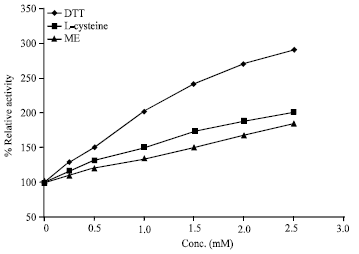 | |
| Fig. 3: | Effect of thiols on gelatin immobilized α-amylase activity |
Effect of thiols and thiol inhibitors: The immobilized α-amylase showed an increase in activity by all the thiols (DTT, L-cysteine and ME) studied (Fig. 3). The activation by DTT was much more pronounced showing about 2.9 fold increase at 2.5 mM concentration. In comparison, L-cysteine and ME showed only 2 fold and 1.84 fold increase in activity, respectively. This observed activation of the soybean α-amylase may be due to the protection of the-SH groups of the enzyme by the added thiol as the thiol reagents itself is consumed by oxygen, leaving the-SH groups of the enzyme intact for the catalytic functions. A similar activation was reported in case of watermelon urease (Prakash and Upadhyay, 2003).
As observed from the effect of the thiol inhibitors; NEM, IAA and pHMB (Fig. 4), there was no marked inhibition maintaining more or less about 70% residual activity at 10 mM indicating that there are not free sulfhydryl groups available in soybean α-amylase to be directly involved in catalysis.
Application of the gelatin immobilized α-amylase in starch stain removal: The ability of the gelatin immobilized α-amylase for starch stain removal was assessed by washing starch stained cotton cloth pieces in detergents alone and in presence of immobilized α-amylase. The starch content removed from the cloth as washout was determined. The washing performance of the detergents is shown in Table 2. The higher amount of starch content indicates better washing. In distilled water, all gave almost similar washing.
| Table 2: | Colorimetric determination of starch content in the washout after washing |
 | |
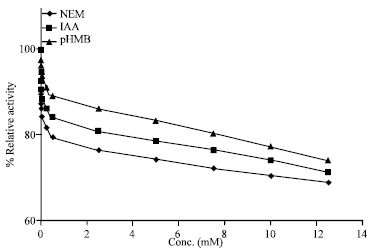 | |
| Fig. 4: | Effect of thiol inhibitors on gelatin immobilized α-amylase activity |
The washing performance of the detergents supplemented with immobilized α-amylase was better as compared to its performance with detergent only. This is based on the property of amylases which are known to hydrolyze the starch and thus removes the starch stain from clothes freely and rapidly. Thus, the use of immobilized α-amylase in starch stain removal by detergents has increased their washing efficiency without being affected by the components of detergents (like surfactants, builders, co-builders, bleach, inhibitors, etc.
CONCLUSION
The results revealed that gelatin immobilized α-amylase formed irreversible and stable aldemine bonds that led the immobilized enzyme to acquire higher temperature optima, increased tolerance to higher pH range and could be reused for starch hydrolysis. All these criteria, could, therefore be successfully utilized in continuous production of maltose. It was found that immobilized α-amylase improved the washing efficiency of detergents and removed starch stains from clothes which are difficult to remove under normal washing conditions. Thus, the immobilized α-amylase could be considered as a potential candidate for use as a cleaning additive in detergents in order to facilitate the removal of starch stains. Due to this promising feature it may find potential application in laundry detergents.
REFERENCES
- Hasirci, N., S. Aksoy and H. Tumturk, 2006. Activation of poly(dimmer acid-co-alkyl polyamine) particles for covalent immobilization of α-amylase. React. Funct. Polym., 66: 1546-1551.
CrossRefDirect Link - Pandya, P.H., R.V. Jasra, B.L. Newalkar and P.N. Bhatt, 2005. Studies on the activity and stability of immobilized α-amylase in ordered mesoporous silicas. Microporous Mesoporous Mater., 77: 67-77.
CrossRef - Altun, G.D. and S.A. Cetinus, 2007. Immobilization of pepsin on chitosan beads. Food Chem., 100: 964-971.
CrossRefDirect Link - De Cordt, S., M. Hendrickx, G. Maesmans and P. Tobback, 1994. The influence of polyalcohols and carbohydrates on the thermostability of α-amylase. Biotechnol. Bioeng., 43: 107-114.
CrossRef - Tumturk, H., S. Aksoy and N. Hasirci, 2000. Covalent immobilization of α-amylase onto poly(2-hydroxyethyl methacrylate) and poly(styrene-2-hydroxyethyl methacrylate) microspheres and the effect of Ca2+ ions on the enzyme activity. Food Chem., 68: 259-266.
CrossRef - Kahraman, M.V., N. Kayaman-Apohan, A. Ogan and A. Gungor, 2006. Soybean oil based resin: A new tool for improved immobilization of α-amylase. J. Applied Polym. Sci., 100: 4757-4761.
CrossRef - El-Banna, T.E., A.A. Abd-Aziz, M.I. Abou-Dobara and R.I. Ibrahim, 2007. Production and immobilization of α-amylase from Bacillus subtilis. Pak. J. Biol. Sci., 10: 2039-2047.
CrossRefPubMedDirect Link - Borgio, J.F., 2011. Immoblization of microbial (wild and mutant strains) amylase on coconut fiber and alginate matrix for enhanced activity. Am. J. Biochem. Mol. Biol.
Direct Link - Egwim, E.C. and O.B. Oloyede, 2008. Immobilization of α-Amylase from Acha (Digiteria exilis) on different cellulose fibre materials. Asian J. Biochem., 3: 169-175.
CrossRefDirect Link - Dey, G., V. Nagpal and R. Banerjee, 2002. Immobilization of alpha-amylase from Bacillus circulans GRS 313 on coconut fiber. Applied Biochem. Biotechnol., 102-103: 303-313.
PubMed - Varavinit, S., N. Chaokasem and S. Shobsngob, 2002. Immobilization of a thermostable alpha-amylase. Science Asia, 28: 247-251.
Direct Link - Kahraman, M.V., G. Bayramoglu, N. Kayaman-Apohan and A. Gungor, 2007. Alpha-amylase immobilization on functionalized glass beads by covalent attachment. Food Chem., 104: 1385-1392.
CrossRef - Turunc, O., M.V. Kahraman, Z.S. Akdemir, N. Kayaman-Apohan and A. Gungor, 2009. Immobilization of α-amylase onto cyclic carbonate bearing hybrid material. Food Chem., 112: 992-997.
CrossRef - Pascoal, A.M., S. Mitidieri and K.F. Fernandes, 2010. Immobilisation of α-amylase from Aspergillus niger on to polyaniline. Food Bioprod. Process.
CrossRef - Ertan, F., H. Yagar and B. Balkan, 2007. Optimization of α-amylase immobilization in calcium alginate beads. Prep. Biochem. Biotechnol., 37: 195-204.
PubMed - Lineweaver, H. and D. Burk, 1934. The determination of enzyme dissociation constants. J. Am. Chem. Soc., 56: 658-666.
CrossRefDirect Link - Lim, L.H., D.G. Macdonald and G.A. Hill, 2003. Hydrolysis of starch particles using immobilized barley α-amylase. Biochem. Eng. J., 13: 53-62.
CrossRef - Reshmi, R., G. Sanjay and S. Sugunan, 2007. Immobilization of α-amylase on zirconia: A heterogeneous biocatalyst for starch hydrolysis. Catal. Commun., 8: 393-399.
CrossRef - Arica, M.Y., V. Hasirci and N.G. Alaeddinoglu, 1995. Covalent immobilization of α-amylase onto pHEMA microspheres: Preparation and application to fixed bed reactor. Biomaterials, 15: 761-768.
CrossRef - Sun, Y.M., J.P. Chen and D.H. Chu, 1999. Preparation and characterization of α-amylase-immobilized thermal-responsive composite hydrogel membranes. J. Biomed. Mater. Res., 45: 125-132.
CrossRef - Tien, C.J. and B.H. Chiang, 1999. Immobilization of α-amylase on a zirconium dynamic membrane. Process Biochem., 35: 377-383.
CrossRefDirect Link - Ikram-ul-Haq, R. Abdullah, H. Ashraf and A.H. Shah, 2002. Isolation and screening of fungi for the biosynthesis of alpha amylase. Biotechnology, 2: 61-66.
CrossRefDirect Link - Hmidet, N., N.E.H. Ali, A. Hadder, S. Kanoun, S.K. Alya and M. Nasri, 2009. Alkaline proteases and thermostable α-amylase co-produced by Bacillus licheniformis NH1: Characterization and potential application as detergent additive. Biochem. Eng. J., 47: 71-79.
CrossRef - Maliks, S.V., V. Kalia and C.S. Pundir, 2000. Immobilization of porcine pancreas lipase on zirconia coated alkylamine glass using glutaraldehyde. Ind. J. Chem. Technol., 7: 64-67.
Direct Link - Rani, P., M. Sharma, V. Kumar and C.S. Pundir, 2007. Immobilization of amylase onto arylamine glass beads affixed inside a plastic beaker: Kinetic properties and application. Indian J. Biotechnol., 6: 230-233.
Direct Link - Dhingra, S., M. Khanna and C.S. Pundir, 2006. Immobilization of α-amylase onto alkylamine glass beads affixed inside a plastic beaker: Kinetic properties and application. Indian J. Chem. Technol., 13: 119-121.
Direct Link - Mitidieri, S., A.H.S. Martinelli, A. Schrank and M.H. Vainstein, 2006. Enzymatic detergent formulation containing amylase from Aspergillus niger: A comparative study with commercial detergent formulations. Bioresour. Technol., 97: 1217-1224.
CrossRef - Prakash, O. and N. Jaiswal, 2010. A highly efficient and thermostable α-amylase from soybean seeds. Biotechnol. Applied Biochem., 57: 105-110.
PubMed - Lowry, O.H., N.J. Rosebrough, A.L. Farr and R.J. Randall, 1951. Protein measurement with the folin phenol reagent. J. Biol. Chem., 193: 265-275.
CrossRefPubMedDirect Link - Tripathi, P., L.L. Leggio, J. Mansfeld, R. Ulbrich-Hofmann and A.M. Kayastha, 2007. α-Amylase from mung beans (Vigna radiata)-correlation of biochemical properties and tertiary structure by homology modelling. Phytochemistry, 68: 1623-1631.
CrossRef - El-Shora, H.M. and M.M. Youssef, 2008. Identification of catalytically essential amino acid residues and immobilization of glutamate dehydrogenase from Rumex cotyledons. Asian J. Biochem., 3: 320-329.
CrossRefDirect Link - Chen, J.P., Y.M., Sun and D.H. Chu, 1998. Immobilization of α-amylase to a composite temperature-sensitive membrane for starch hydrolysis. Biotechnol. Prog., 14: 473-478.
CrossRef - Aksoy, S., H. Tumturk and N. Hasirci, 1998. Stability of α-amylase immobilized on poly(methyl methacrylate-acrylic acid) microspheres. J. Biotechnol., 60: 37-46.
CrossRef - Bayramoglu, Z., U. Akbulut and S. Sungur, 1992. Immobilization of α-amylase into photographic gelatin by chemical cross-linking. Biomaterials, 13: 704-708.
CrossRef - Demircioglu, H., H. Beyenal, A. Tanyolac and N. Hasirci, 1995. Entrapment of urease in glycol containing polymeric matrices and estimation of effective diffusion coefficients of urea. Polymer, 36: 4091-4096.
CrossRef - Lee, P.M., K.H. Lee and S.Y. Siaw, 1993. Covalent immobilization of aminoacrylase to alginate for L-phenulalanine production. J. Chem. Technol. Biotechnol., 58: 65-70.
CrossRef - Cosulich, M.E., S. Russo, S. Pasquale and A. Mariani, 2000. Performance evaluation of hyperbranched aramids as potential supports for protein immobilization. Polymer, 41: 4951-4956.
CrossRef - Park, D., S. Haam, K. Jang, I.S. Ahn and W.S. Kim, 2005. Immobilization of starch-converting enzymes on surface-modified carriers using single and co-immobilized systems: Properties and application to starch hydrolysis. Process Biochem., 40: 53-61.
CrossRef - Prakash, O. and L.S.B. Upadhyay, 2003. Effect of thiols on the activity of urease from dehusked seeds of watermelon (Citrullus vulgaris). Plant Sci., 164: 189-194.
CrossRef - Kara, A., B. Osman, H. Yavuz, N. Besirilana and A. Denizli, 2005. Immobilization of α-amylase on Cu+2 chelated poly(ethylene glycol dimethacrylate-n-vinyl imidazole) matrix via adsorption. Reactive Func. Polym., 62: 61-68.
CrossRefDirect Link