
S. Mandhania
Department of Biochemistry, CCS Haryana Agricultural University, Hisar, India
V. Jain
Department of Biochemistry, CCS Haryana Agricultural University, Hisar, India
S.P. Malhotra
Department of Biochemistry, CCS Haryana Agricultural University, Hisar, India
Asian Journal of Biochemistry
Year: 2010 | Volume: 5 | Issue: 1 | Page No.: 12-22







ABSTRACT
The present investigations were undertaken with the objective to isolate PME producing microbial strain and to standardize conditions to have cost effective production of the enzyme. Based on the ability of pectinolysis, out of 20 microbial strains isolated, one fungal strain was selected and got identified from Institute of Microbial Technology (IMTECH), Chandigarh (India), as Aspergillus heteromorphus. The strain was added to its collection with identification number MTCC 9262. Various process variables viz., type and pH of the medium, temperature of growth, time of incubation, inoculum concentration and type and concentration of carbon and nitrogen sources were optimized as per the conventional method of one variable at a time approach which involves varying a single independent variable and maintaining others at a constant level. Modified pectin medium-A, in which both Fe++ and Cu++ were omitted, gave the maximum growth of A. heteromorphus. Maximum PME production was obtained when an inoculum size of 4x106 spores mL-1 was incubated in modified pectin medium-A for 144 h at pH 4.5 and at a temperature of 30°C under static conditions in submerged fermentation. Out of the various nitrogen and carbon sources tested, dibasic ammonium phosphate and pectin were found to be the best N and C source, respectively. After standaradizing various process variables, a three fold increase in PME production by A. heteromorphus was achieved. Orange peel, a waste byproduct of fruit industry could effectively replace pectin as C-source thus, making the process cost effective. The PME produced by A. heteromorphus had the pH optimum in acidic range-a characteristic which could be exploited for its use in fruit processing industry.
PDF Abstract XML References Citation
How to cite this article
S. Mandhania, V. Jain and S.P. Malhotra, 2010. Culture Optimization for Enhanced Production of Microbial Pectin Methylesterase under Submerged Conditions. Asian Journal of Biochemistry, 5: 12-22.
DOI: 10.3923/ajb.2010.12.22
URL: https://scialert.net/abstract/?doi=ajb.2010.12.22
DOI: 10.3923/ajb.2010.12.22
URL: https://scialert.net/abstract/?doi=ajb.2010.12.22
INTRODUCTION
Pectins, the complex high molecular weight heterogeneous and acidic structural polysaccharides are the major constituents of primary cell wall of cereals, vegetables, fruits and fibers. The main component of pectin backbone is galacturonic acid with neutral sugars such as arabinose, galactose and xylose present in the side chains, whereas rhamnose constitutes a minor component of pectin. Particularly, in fruits and vegetables, pectins are present in large amounts contributing to the strength of these tissues. They are one of the important functional food ingredients in jams, jellies, fruit juices, confectionery products, bakery fillings and are used for stabilization of acidified milk drinks and yoghurts. Pectinolytic enzymes, that profoundly affect the fine structure of pectin, find applications in the various industrial processes. The most commonly used industrial pectinases which account for about 10% of the global industrial enzymes (Stutzenberger, 1992), include polygalacturonases, pectin lyases and pectin methylesterases (PMEs). Acidic PME is used in extraction, clarification and liquification of fruit juices (Kaur et al., 2004), while alkaline PME finds potential applications in paper and textile industry (Ahlawat et al., 2009). In addition, these enzymes also have the potential of producing low degree methylated pectins. Biotechnological potential of microorganisms producing PME in textile, paper and fruit processing industry (Reid and Ricard, 2000) has drawn a great deal of attention worldwide. Though numbers of PME producing organisms such as Aspergillus niger (Bhat, 2000; Kashyap et al., 2001). Aspergillus awamori (Blandino et al., 2002), Aspergillus foetidus (Sebastian et al., 1996; Hours et al., 1988), Aspergillus japonicum (Semenova et al., 2003), Aspergillus carneus (El-Sheekh et al., 2009), Aspergillus flavus (Yadav et al., 2008) and Bacillus subtilis (Ahlawat et al., 2009) have been reported, selection of potential isolates remains a tedious task, especially, when physiologically potential strains are to be obtained to achieve maximum yield (Pandey et al., 1999). Several researchers have studied the optimization of culture conditions for enhanced PME production from various microorganisms (Maldonado et al., 1986; Maldonado and de-Saad, 1998; Joshi et al., 2006; Ahlawat et al., 2009; El-Sheekh et al., 2009). We report here, the standardization of process variables for the maximum production of PME in submerged fermentation of a physiologically potential strain of Aspergillus isolated from the soil debris of fruit and vegetable markets.
MATERIALS AND METHODS
Micro-Organism and Inoculum
The present investigations were carried out in the Department of Biochemistry, CCS Haryana Agricultural University, Hisar (Haryana), India from Nov. 2008 to April, 2009. Microorganisms were isolated from soil samples collected from different fruit and vegetable markets of the city. Based on the ability of pectin hydrolysis and potential of PME production, a total of twenty isolates were screened. Finally, one fungal isolate was selected as the potential producer of PME which was got identified from the Institute of Microbial Technology (IMTECH), Chandigarh, India as Aspergillus heteromorphus. It had been included to their collection at IMTECH with identification mark MTCC 9262. The inoculum was prepared in Potato Dextrose Medium (PDM) containing pectin as the sole C-source, by harvesting spores from 120 h old cultures grown at 30°C.
Fermentation
Submerged fermentation was carried out in 250 mL Erlenmeyer flasks by taking 50 mL pectin medium containing (%, w/v). (NH4)2SO4-0.3; KH2PO4-1.0; MgSO4-0.2; Na2B4O7-0.07; (NH4)6MO7O24-0.05; Fe(SO4)3-0.001, CuSO4-0.0003; MnSO4-0.0001; ZnSO4-0.007; Pectin-0.3 (SRL). Flasks were inoculated with inoculum size of 1x105 spores mL-1 and incubated at 30°C for 144 h under static conditions in a BOD incubator (Remi, India).
For maximum production of PME, various culture conditions viz., type and concentration of carbon and nitrogen sources, type of medium, inoculum size, pH, temperature and incubation period were optimized by conventional method of standardization by one variable at a time approach which involves varying a single independent variable at a time while maintaining the others at a constant level. Fungal isolate was grown in different media, containing 0.3% pectin, at pH 4.2 for 120 h. Medium giving maximum growth was further used to standardize the inoculum size by inoculating different concentrations (1-5x106 spores mL-1) of inoculum. Similarly, the temperature and time of incubation were optimized by growing the fungal isolate at different temperatures (25, 30, 35, 40, 45 and 50°C) for different time periods (72-192 h), respectively. Different organic and inorganic nitrogen sources viz., yeast extract, peptone, ammonium chloride, ammonium hydrogen phosphate, ammonium sulfate and sodium nitrate were supplemented separately to a final concentration of 0.3% (w/v) to study the microbial growth and PME activity. Glucose, fructose, sorbitol, sucrose, maltose, pectin and citrus peel at a final concentration of 1% were used as the carbon sources.
Process Variables
Effect of carbon and nitrogen sources and their concentrations, type of medium, inoculum size, pH, incubation temperature and time period, was examined in submerged condition. Statistical analysis of standard deviations was done to determine the effect of variables on production of PME using software Statistical Package for Agricultural Scientists OPSTAT.
Enzyme Isolation and Assay
After 144 h of growth, the fungal broth was filtered through 4 layers of muslin cloth and centrifuged at 8000 rpm for 15 min at 4°C in a refrigerated centrifuge. The supernatant taken as enzyme extract containing extracellular PME, was used to assay the enzyme activity. Ten milliliter of 1% pectin in 0.15 M NaCl (pH 7.0) was incubated with 2 mL of enzyme extract in 100 mL flask at 35°C for 1 h. The reaction was stopped by keeping the flask in boiling water bath for 10 min. The contents were cooled and the free carboxyl groups released were determined by titrating against 0.02 N NaOH using phenolphthalein as indicator (Kertesz, 1955). One unit of PME was defined as the amount of enzymes required to release one micro equivalent of carboxyl group/mL/h.
RESULTS
Identification of PME Producing Microorganism
Out of twenty wild cultures of fungal isolates, only five isolates showing clear zone (Fig. 1a, b) on agar plates with pectin medium were selected as PME producers. These isolates were further screened for the maximum production of PME on two media viz., potato dextrose medium and pectin medium (Fig. 2) at pH 4.2 with 0.3% pectin as sole carbon source.
 | |
| Fig. 1: | Clear zone formation showing PME activity on pectin agar medium (a) after 3 days of incubation and (b) after 5 days of incubation |
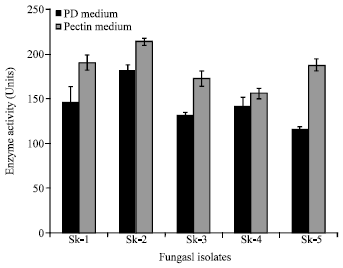 | |
| Fig. 2: | Production of PME by five fungal isolates on potato dextrose (PD) and pectin medium |
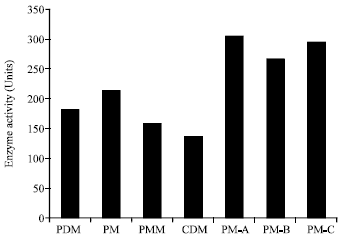 | |
| Fig. 3: | Effect of different media on PME production by Aspergillus heteromorphus MTCC 9262 |
As is clear from the results, isolate SK-2, exhibited highest PME production in both the media (160 and 213 units in potato dextrose medium and pectin medium, respectively) and was used for further studies for optimizing culture conditions to get maximum production of PME. This fungal isolate was got identified from IMTECH, Chandigarh as Aspergillus heteromorphus and was included to their collection at the centre with identification mark MTCC 9262.
Optimization of Culture Medium
The Aspergillus heteromorphus MTCC 9262 was initially grown on four different media (pH 4.2) viz., Potato Dextrose Medium (PDM), Pectin Medium (PM), Pectin Minimum Medium (PMM; it contained no micronutrients) and Czapeck Dox Medium (CDM). As is clear from the results presented in Fig. 3, the enzyme activity was maximum (213 units) in PM containing all the micronutrients and minimum in CDM (136 units). Since, pectin minimal medium which contained no micronutrients also showed 76% of the maximum activity, it was thought worthwhile to omit the micronutrients one by one and see the effect on growth and enzyme activity. When either Zn or Mn or B or Mo was omitted individually or in combination from pectin medium, the activity was same as observed in pectin minimal medium, indicating that these micronutrients were the absolute requirements for the growth of the A. heteromorphus and could not be omitted from the medium. However, when either Cu or Fe was omitted, there was rather improvement in PME activity over that of pectin medium. Keeping this in view, A. heteromorphus was grown in three modified pectin media viz., A (where, Fe and Cu both were omitted), B (where Cu alone was omitted) and C (where Fe alone was omitted). Results presented demonstrate that when grown in modified pectin medium-A, in which both Cu and Fe both were omitted, Aspergillus heteromorphus showed maximum PME producing activity (305 units) and the activity was even higher than that observed with pectin medium (213 units).
Optimization of Culture Conditions
Inoculum Concentration/Size
The size of inoculum seems to have a profound effect on growth of the microorganisms and enzyme production. PME production activity was studied by varying the inoculum size from 1mL (1x106spores) to 5 mL (5x106 spores). The maximum enzyme production (361 units) was observed with 4 mL inoculum i.e., 4x106 spores. When inoculum size was further increased to a size more than 4 mL, enzyme production declined (Fig. 4a-d).
pH
The production of PME by A. heteromorphus MTCC 9262 was studied by growing the fungus in PM-A medium with pH ranging from 4.0 to 6.5 in submerged fermentation. Results (Fig. 4) revealed that pH 4.5 was optimum for the maximum production of PME. On either side of the optimum pH (4.5), the enzyme production declined.
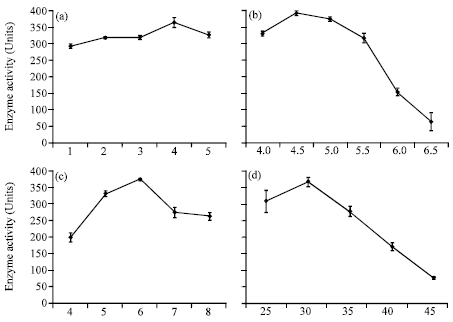 | |
| Fig. 4: | Effect of different cultural conditions on the PME production by Aspergillus heteromorphus MTCC 9262, (a) inoculum size (mL), (b) pH, (c) days of incubation and (d) incubation temperature |
Inoculation Time
The PME production was detected after three days i.e., after 72 h of incubation. After 72 h, the activity was measured at 24 h interval till 192 h. The results in Fig. 4 demonstrate maximum PME production to occur after 144 h of incubation with the yield of 377 units. After 144 h, the activity declined.
Temperature
To find out the optimum temperature of growth and enzyme production, the organism was grown at different temperatures ranging from 25 to 45°C. Optimum temperature observed in this study was 30°C (Fig. 4) for the maximum production of PME by A. heteromorphus in submerged conditions.
Nitrogen Source
When different inorganic and organic nitrogen sources were supplemented in the medium to achieve maximum PME production by A. heteromorphus, peptone and dihydrogen ammonium phosphate were observed to be better as they enhanced the PME production to 480 (28% of control) and 520 (39% of control) units, respectively (Fig. 5). Among the various nitrogen sources tested, ammonium sulphate was at number three.
Carbon Source
The carbon source in the medium appears to exert a profound effect on the enzyme production behaviour of the fungus. Pectin at 1% concentration gave the maximum PME production (613 units). Citrus peel at a concentration of 1% showed almost identical PME production (543 units) which was next to the activity observed with 1% pectin (Fig. 5).
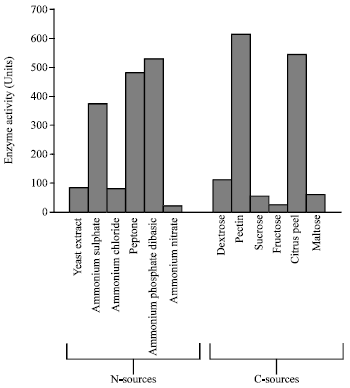 | |
| Fig. 5: | Effect of different nitrogen and carbon sources on PME production by Aspergillus heteromorphus MTCC 9262 |
| Table 1: | Comparison of PME production by Aspergillus heteromorphus MTCC 9262 under initial and optimized culture conditions |
Based on these observations, citrus peel being a readily available waste by product of citrus fruit industry, could be suggested as an efficient carbon source for PME production by Aspergillus heteromorphus. No PME activity could be observed when pectin was replaced by sucrose, maltose, dextrose and fructose as the carbon sources suggesting that PME in A. heteromorphus is substrate inducible.
Finally, the fungal strain of Aspergillus heteromorphus MTCC 9262 was grown in 250 mL Erlenmeyer flask containing 50 mL of modified pectin medium-A with 0.3% citrus peel as the C-source and 0.2% dihydrogen ammonium phosphate as the N-source at pH 4.5 and 30°C for 144 h inoculated with 4x106 spores mL-1. Results presented in Table 1 reveal that optimization of process variables resulted in 3-fold increase in PME production.
DISCUSSION
In recent years, considerable attention has been paid to the use of micro-organisms in industrial fermentation processes, especially enzyme production There has been a surge of interest in the production of fungi having the ability to produce acidic pectinases which can be used in the fruit processing industry where they play a crucial role in the extraction and clarification of fruit juices. In order to obtain concentrates of high quality, it is essential to degrade pectin and clarify the juices with the help of pectolytic enzymes. It is very well reported that pectolytic enzymes produced by certain sprouting strains like Aspergillus niger, Aspergillus oryzae, Rhizopus oryzae etc., have long been used in fruit juice clarification because they have been observed to have desirable characteristics to be utilized for that purpose. However, these fungal strains have not been exploited to scale up the production of these enzymes for the purpose of their purification and immobilization. The search for specific enzymatic activities is still increasing due to their demand for clarification of new types of fruit juices. For this reason, the present investigations were undertaken with the objective to harness the valuable product namely fungal/microbial isolate and to examine its potential to produce pectin methylesterase. During the present investigations, twenty isolates were screened on the basis of their ability of pectin hydrolysis on agar plates containing pectin and the dye ruthidium red. The appearance of clear zone was taken as the growth of fungus. Similar strategy of screening of fungal cultures was adapted by Boccas et al. (1994). Out of twenty, five isolates giving appreciably good amount of PME were grown on potato dextrose and pectin media. The isolate-2 gave maximum activity in both the media and was selected for further studies. Microbial cultures, regardless of the nature of their end product and the type of bioprocess have certain specific requirements for their growth which have to be optimized for their maximum production. The growth conditions optimized for the fungal strain selected in the present investigations included type of medium, its temperature, pH, inoculum concentration, incubation time and type and concentration of C- and N-source. When grown on different media, it was observed that the modified pectin medium A in which both Fe and Cu were omitted gave the maximum PME production. This may be due to inhibitory effect of Cu and Fe on PME activity (Celestino et al., 2006). Cu has been reported to inhibit the growth of the microorganisms while the Fe has been found to interfere with the color of ruthidium red while counting the microbial colonies. Maximum production of PME when inoculated with 4x106 spores of A. heterolorphus and decrease in its production on increasing the inoculum size (Fig. 4) could be due to competition between microorganism colonies for nutrients and probably the non-availability of nutrients for the large population limits the fungal growth. Similar observations have been made by Maldonado and Callieri (1989) where inoculum size of 4x106 spores has been shown to give maximum growth. However, there are several reports showing the need of either less (Maldonado and de-Saad, 1998; Bai et al., 2004) or more (Cavalitto et al., 1996) size of inoculum for maximum production of PME from different sources. This might be due to differences in the nutrient requirements by different microorganisms.
When grown in medium with different pH values, maximum production of enzyme was observed at pH 4.5. On either side of this optimum pH, the PME producing ability declined. The variation in PME production due to change in pH may be because of maximum availability of nutrients at that particular pH, as has been reported by Joshi et al. (2006). Semenova et al. (2003) also reported that the production of pectinase by A. japonicum was maximum in acidic medium. The influence of wide range of pH from 2.3 (Sebastian et al., 1996) to 7.2 (Parley and Page, 1971) on the production of microbial pectinase from different substrates has been reported. Wide range of pH of the medium during upstream bioprocesses makes the end product either acidic or alkaline, which tends to have varied applications (Hoondol et al., 2002). Acidic pectinases are useful in extraction, clarification and liquifaction of fruit juices (Kaur et al., 2004) and wines (Favela et al., 2005), while alkaline pectinases are widely used in fabric industries for setting of plant fibers such as flax, hemp and jute, in biopreparation of cotton fabrics, in enzymatic polishing of jute/cotton blended fabrics in the paper industry to solve the retention problems in mechanical pulp bleaching, in the treatment of pulp and paper effluents and for improving the flavour of black tea (Sharma and Satyanarayana, 2004; Favela et al., 2005). Aspergillus heteromorphus, studied during the present investigations, is suggested to be the producer of acidic PME and may find its applications in fruit juice industry.
Maximum production of PME at 144 h of incubation as has been observed in the present investigations has also been reported by Sunnotel and Nigam (2002). However, in their case, prolonged fermentation up to 288 h resulted in comparatively very small decline in enzyme activity, whereas in the present case, the activity declined significantly after 144 h of incubation suggesting that enzyme produced by A. heteromorphus was more unstable than the one described by Sunnotel and Nigam (2002). Temperature is another very important factor which is known to influence the metabolic rate of the organism involved in the process, which in turn determines the amount of the end product. Temperature has been observed to be one of the major process variables affecting the production of pectinases in submerged conditions. Optimum temperature of 30°C for maximum production observed in present case falls within the range of 21°C (Joshi et al., 2006) to 35°C (Soni and Bhatia, 1981) already reported.
When different inorganic and organic nitrogen sources were supplemented in the medium to achieve maximum PME production by A. heteromorphus, peptone and dihydrogen ammonium phosphate were observed to be better as they enhanced the PME production to 480 (28% of control) and 520 (39% of control) units, respectively (Fig. 5). Ammonium phosphate (dibasic) has been reported to be the best nitrogen source for the growth of A. niger (Joshi et al., 2006). Ammonium chloride, ammonium nitrate and yeast extract inhibited PME production. Kashyap and Soni (2003) have also reported ammonium nitrate to be inhibitory for pectinase production by Bacillus. To find out its optimum concentration, the fungal isolate was grown in different concentrations (ranging from 0.1-0.4%) of dibasic ammonium phosphate and it was observed that 0.2% gave the maximum PME production. The present results are in accordance with those reported by Joshi et al. (2006) where A. niger has been observed to produce maximum PME with 0.2% dibasic ammonium phosphate.
In the present investigations, the PME production activity observed with citrus peel at a concentration of 1% was almost identical to the activity observed with 1% pectin thus, making the process cost effective. Similarly, very recently (El-Sheekh et al., 2009), orange peel and pulp scrap excluded as waste in orange juice and canning industry, have been successfully used for the production of pectinase by Aspergillus carneus NRC 1. No PME activity could be observed when pectin was replaced by sucrose, maltose, dextrose and fructose as the carbon sources suggesting that PME in A. heteromorphus is substrate inducible. Many researchers have used pectin as a sole carbon source for PME production and found the enzyme to be inducible one (Oretga, 1994; Maldonado and de-Saad, 1998; Jaffar and Oommen, 1993). Kashyap and Soni (2003) however, reported that carbon sources such as polygalacturonic acid, pectin and lactose enhanced pectinase production by 11-44% while glucose, galactose, sucrose, maltose and xylose inhibited pectinase production by 25-60% in Bacillus.
After optimization of process variables, a 3-fold increase in PME production was observed. A similar increase of 2.5 fold has been obtained in the activity of tyrosine phenol lyase by Citrobacter freundii MTCC 2424 after optimizing the culture conditions (Chandel and Azmi, 2008). Sharma and Satyanarayana (2006) reported about 34-41 fold increase in pectinases production after optimization of process variables by statistical method of Response Surface Methodology (RSM).
CONCLUSION
Isolation and multi-step screening method employed in the study led to the isolation of one efficient strain of Aspergillus heteromorphus for production of PME. Results obtained on optimization of process variables under submerged fermentation revealed that citrus peel could be effectively used for PME production. About 3 fold enhancement in acidic PME production by A. heteromorphus MTCC 9262 was achieved when an inoculum size of 4x106 spores mL-1 was incubated in modified pectin medium-A at pH 4.5 and 30°C for 144 h. The low cost of its production and acidic nature of enzyme may further broaden the scopes for its use in fruit juice industries.
ACKNOWLEDGMENT
Researchers are thankful to Haryana State Council of Science and Technology, Chandigarh, India for the financial assistance in the form of the research project entitled Production and immobilization of microbial enzymes for fruit processing industry.
REFERENCES
- Bai, Z.H., H.X. Zhang, H.Y. Qi, X.W. Peng and B.J. Li, 2004. Pectinase production by Aspergillus niger using wastewater in solid state fermentation for eliciting plant disease resistance. Bioresour. Technol., 95: 49-52.
CrossRefDirect Link - Bhat, M.K., 2000. Cellulases and related enzymes in biotechnology. Biotechnol. Adv., 18: 355-383.
Direct Link - Blandino, A., T. lqbalsyah, S.S. Pandiella, D. Cantero and C. Webb, 2002. Polygalacturonase production by Aspergillus awamori on wheat in solid-state fermentation. Appl. Microbiol. Biotechnol., 58: 164-169.
Direct Link - Boccas, F., S. Roussos, M. Gutierrez, L. Serrano and G.G. Viniegra, 1994. Production of pectinase from coffee pulp in solid-state fermentation system: selection of wild fungal isolate of high potency by a simple three-step screening technique. J. Food Sci. Technol., 31: 22-26.
Direct Link - Cavalitto, S.F., J.A. Arcas and R.A. Hours, 1996. Pectinase production profile of Aspergillus foetidus in solid state culture at different acidities. Biotechnol. Lett., 18: 251-256.
Direct Link - Celestino, S.M.C., S.M.D. Freitas, F.J. Medrano, M.V.D. Sousa and E.X.F. Filho, 2006. Purification and characterization of a novel pectinase from Acrophialophora nainiana with emphasis on its physicochemical properties. J. Biotechnol., 123: 33-42.
Direct Link - Chandel, M. and W. Azmi, 2008. Optimization of process parameters for the production of tyrosine phenol lyase by Citrobacter freundii MTCC 2424. Bioresour. Technol., 100: 1840-1846.
Direct Link - El-Sheekh, M.M., A.S. Ismail, M.A. El-Ab, E.M. Hegazy and A.I. El-Diwany, 2009. Effective technological pectinases by Aspergillus carneus NRC1 utilizing the Egyptian orange juice industry scraps. Int. Biodeterioration Biodegradation, 63: 12-18.
CrossRef - Hoondal, G., R. Tiwari, R. Tewari, N. Dahiya and Q. Beg, 2002. Microbial alkaline pectinases and their industrial applications: A review. Applied Microbiol. Biotechnol., 59: 409-418.
CrossRefDirect Link - Hours, R.A., C.E. Voget and R.J. Ertola, 1988. Some factors affecting pectin-ase production from apple pomace in solid-state-cultures. Biol. Wastes, 24: 147-157.
Direct Link - Jaffar, M.B. and S. Oommen, 1993. Production, purification and characterization of pectin methylesterase (PME) from Arthrobotrys oligospora. J. Food Biochem., 17: 53-65.
Direct Link - Joshi, V.K., M. Parmar and N.S. Rana, 2006. Pectin esterase production from apple pomace in solid-state and submerged fermentations. Food Technol. Biotechnol., 44: 253-256.
Direct Link - Kashyap, D.R., P.K. Vohra, S. Chopra and R. Tewari, 2001. Applications of pectinases in the commercial sector: A review. Bioresour. Technol., 77: 215-227.
CrossRefPubMedDirect Link - Kashyap, D.R., K.S. Soni and R. Tewari, 2003. Enhanced production of pectinase by Bacillus sp. DT7 using solid-state fermentation. Bioresour. Technol., 88: 251-254.
CrossRef - Kaur, G., S. Kumar and T. Satyanarayana, 2004. Production, charac-terization and application of a thermostable polygalacturonase of a thermophilic mould Sporotrichum thermophile apinis. Bioresour. Technol., 94: 239-243.
CrossRef - Maldonado, M.C. and D.A.S. Callieri, 1989. Influence of environmental conditions on the production of pectinesterase and polygalacturonase by Aspergillus niger. Mircen. J., 5: 327-333.
Direct Link - Maldonado, M.C. and S.A.M. de Saad, 1998. Production of pectinesterase and polygalacturonase by Aspergillus niger in submerged and solid-state systems. J. Ind. Microbiol. Biotechnol., 20: 34-38.
Direct Link - Maldonado, M.C., A. Antonio Navarro and D.A.S. Callieri, 1986. Production of pectinases by Aspergillus sp. using differently pretreated lemon peel as the carbon source. Biotechnol. Lett., 8: 501-504.
Direct Link - Reid, I. and M. Ricard, 2000. Pectinase in papermaking: solving retention problems in mechanical pulps bleached with hydrogen peroxide. Enzyme Microbial Technol., 26: 115-123.
CrossRef - Sebastian, F.C., A.A. Jorge and A.H. Roque, 1996. Pectinase production pro-file of Aspergillus foetidus in solid-state cultures at different acidities. Biotechnol. Lett., 18: 251-256.
Direct Link - Semenova, M.V., S.G. Grishutin, A.V. Gusakov, O.N. Okunev and A.P. Sinitsyn, 2003. Isolation and properties of pectinases from the fungus Aspergillus japonicus. Biochemistry, 68: 559-569.
CrossRefDirect Link - Sharma, D.C. and T. Satyanarayana, 2004. Production and Application of Pectinolytic Enzymes of Sporotrichurm thermophile and Bacillus pumilus. In: Biotechnological Approaches for Sustainable Development, Reddy, M.S. and S. Khanna (Eds.). Allied Publishers Pvt. Ltd., India, ISBN: 81-7764-669-9, pp: 164-169.
- Sharma, D.C. and T. Satyanarayana, 2006. A marked enhancement in the production of a highly alkaline and thermostable pectinase by Bacillus pumilus dcsr1 in submerged fermentation by using statistical methods. Biores. Technol., 97: 727-733.
Direct Link - Sunnotel, O. and P. Nigam, 2002. Pectinolytic activity of bacteria isolated from soil and two fungal strain during submerged fermentation. World J. Microbiol. Biotechnol., 18: 835-839.
Direct Link - Yadav, S., P.K. Yadav, D. Yadav and K.D.S. Yadav, 2008. Purification and characterization of an alkaline pectin lyase from Aspergillus flavus. Process Biochem., 43: 547-552.
CrossRef - Ahlawat, S., S.S. Dhiman, B. Battan, R.P. Mandhan and J. Sharma, 2009. Pectinase production by Bacillus subtilis and its potential application in biopreparation of cotton and micropoly fabric. Process Biochem., 44: 521-526.
CrossRef