
Wong Ling Shing
Faculty of Science and Technology, University Kebangsaan Malaysia, 43600 Bangi, Selangor, Malaysia
Salmijah Surif
Faculty of Science and Technology, University Kebangsaan Malaysia, 43600 Bangi, Selangor, Malaysia
Lee Yook Heng
Faculty of Science and Technology, University Kebangsaan Malaysia, 43600 Bangi, Selangor, Malaysia
Asian Journal of Biochemistry
Year: 2008 | Volume: 3 | Issue: 3 | Page No.: 162-168







ABSTRACT
A biosensor based on electrochemical transduction using an oxygen probe has been developed for the measurement of Cd2+ toxicity by using cyanobacteria (Anabaena torulosa), where the cells were immobilized on the surface of an oxygen probe. The biosensor responded to the changes in photosynthetic oxygen release under illumination by a light source. Exposure to Cd2+ at concentrations below approximately 8 mg L-1 did not demonstrate any inhibition but stimulation of oxygen production was observed. Inhibition occurred only when concentration of Cd2+ was above 8 mg L-1. Using the same concentration range of Cd2+ but increasing the exposure time of Cd2+ from 15 to 30 min, inhibition of oxygen release was mainly observed. This pattern of inhibition of A. torulosa by Cd2+ was different from that of Cu2+, which was also measured by the same toxicity biosensor, where no stimulation in oxygen release was observed. The use of toxicity biosensor has enabled the detail toxicity behaviour of A. torulosa towards Cd2+ to be evaluated.
PDF Abstract XML References Citation
How to cite this article
Wong Ling Shing, Salmijah Surif and Lee Yook Heng, 2008. Toxicity Biosensor for the Evaluation of Cadmium Toxicity Based on Photosynthetic Behavior of Cyanobacteria Anabaena torulosa. Asian Journal of Biochemistry, 3: 162-168.
DOI: 10.3923/ajb.2008.162.168
URL: https://scialert.net/abstract/?doi=ajb.2008.162.168
DOI: 10.3923/ajb.2008.162.168
URL: https://scialert.net/abstract/?doi=ajb.2008.162.168
INTRODUCTION
Whole cell based biosensors can respond to a wide range of changes in their environment and suitable for use in toxicity test and environmental monitoring where the sources and nature of the toxicant cannot be predicted (Bentley et al., 2001). The biosensors response to real physiological impact of active compounds present in the samples and act as broad band monitoring for toxins, which might not necessary discriminate between the different types of toxicants (Giardi et al., 2001; Mattiasson 1997; Roger, 1995; Carpentier et al., 1991; Evans et al., 1986). Biosensors can be configured to be sensitive and inexpensive to manufacture and can be significant in terms of reducing cost and increase the efficiency of certain environmental monitoring applications (Roger, 1995).
Anabaena torulosa is a type of filametous cyanobacteria (cyanophyceae) from Nostocaceae family, which formally known as blue-green algae (Linda, 2000). The organism contains chlorophyll a and undergoes photosynthesis and the physiology changes can be observed by oxygen evolution. Based on the research by Chay et al. (2005), the organism has shown potential as a good biosensor to copper (Cu), lead (Pb) and 2,4-Dichlorophenoxyacetic acid (2,4-D). However, the previous studies had not investigated the usefulness of cyanobacteria A. torulosa as a biosensor for Cd toxicity evaluation. In view of Cd is a toxicant, which is non-essential, non-beneficial and possesses high toxic potential (Giardi et al., 2001; Lockwood, 1976) and a common pollutant in water resources (Heever and Grobbelaar, 1998), the focus of this study is to assess the usefulness of A. torulosa as a biosensor for rapid Cd toxicity evaluation.
MATERIALS AND METHODS
Reagents
Copper (2) nitrate, Cu(NO3)2, Cadmium (2) nitrate, Cd(NO3)2 (Merck, Germany), poli-hydroxylethyl methacrylate, (pHEMA) (Sigma UK), 1,4-dioxane (Fisher, UK), cyanobacteria Anabaena torulosa (Carolina Biological Supply Co., US), Bold`s Basic Medium (James, 1979) as culture medium for Anabaena torulosa. All reagents were prepared in distilled water.
A. torulosa Culture
A. torulosa was cultured in Bold`s Basic Medium at 18.5°C, 1000 Watt unit-1 area white fluorescent illumination, with light and dark period maintained at 16 and 8 h, respectively in culture chamber (GC-500, Protech). Each liter of Bold`s Basic Medium prepared contains 10 mL of several macronutrients stocks and 1 mL of trace elements stocks respectively (Table 1). Aeration was carried out by manual shaking three times daily to avoid cells clumping.
Determination of Cells Growth
The number of cells was determined by using a Microscope BX51 (Olympus, USA) and a Weber haemocytometer. Concentration of suspended cell can be estimated by using UV/VIS spectrophotometer (Perkin Elmer). The optical density at 700 nm can be used to estimate the cell density for immobilization as the two quantities were closely related. OD at 700 nm is the absorption peak of chlorophyll a in A. torulosa (Desikachary, 1959). The growth of the cyanobacteria was determined every 2 days for 14 days to establish the growth profile and cells at the most active phase of the growth were selected for toxicity biosensor studies (Chay et al., 2005).
| Table 1: | The preparation of stocks for Bold`s Basic Medium |
 | |
Cyanobacteria Immobilization
The pHEMA solution was prepared by dissolving the polymer in a mixture of water and dioxane with the ratio 4:1 and 1 mL of solution contained 60 μg of pHEMA. By using 50 μL of pHEMA solution, a certain amount of A. torulosa cells were coated on a Teflon gas permeable membrane. The cyanobacteria entrapped in the pHEMA film were left to dry at 18.5°C for 4-5 h. The membrane was then assembled onto the tip of an oxygen electrode (Orion, USA) by using a rubber O-ring. The oxygen evaluation can be measured by a change of dissolved oxygen when the immobilized cells were illuminated by a 100 W lamp (Phillip Classictone, Holland) compared to non-illuminated condition. The change of oxygen level was recorded using a PC (DO sensorlink PCM 800, Thermo Orion, USA).
Exposure of A. torulosa to Various Concentrations of Cadmium Ions
Cd toxicity tests were conducted for both immobilized and free cells of A. torulosa. The amount of pHEMA used for immobilization, cell density and cell culture age were optimized. Each biosensor was used only once for toxicity evaluation at each concentration of Cd. The experiment for immobilized cyanobacteria was performed in pH 7 and at room temperature, as stated by Chay et al. (2005). The biosensor was left in 10 mL of distilled water for 15 min until a stable baseline was obtained. The photosynthetic oxygen release of the immobilized A. torulosa cells was determined continuously for 5 min after light illumination began. The biosensor probe was then incubated in 10 mL of various concentrations of Cd2+ (0.1, 0.25, 0.5, 1.0, 5.0, 10.0, 20.0 mg L-1) solutions for 15 min at room temperature. After the period, the probe was then transferred to 10 mL of distilled water again and illuminated to allow photosynthetic activity for another 5 min. Any inhibition of the immobilized cells on the oxygen probe can be calculated as follows:
where,
| Io | = | Highest amount of oxygen evolved before exposure to the Cd ions within a fixed duration. |
| I | = | Highest amount of oxygen evolved after exposure to the Cd ions within a fixed duration. |
| The highest amount of oxygen | = | The highest oxygen value observed within a fixed duration - the baseline oxygen value |
In free cell (i.e., non-immobilized cells) toxicity evaluation for A. torulosa, 10 mL A. torulosa cells with an absorbance unit of 1.0 Abs at 700 nm was left for 15 min until a stable baseline was obtained. Further illumination for 5 to 10 min was performed to oxygen evolution. Various amount (<0.1 mL) of 100 μg L -1 of Cd solution was the added to the free cell solution to make the total Cd concentration range from 0.1-5.0 μg L-1. It was then incubated for 15 min. Illumination was conducted once again at 5 to 10 min to determine the oxygen evaluation. The percentage of photosynthetic inhibition was then calculated.
RESULTS AND DISCUSSION
Immobilization techniques in biosensor offer advantages in terms of controlling the number of cells, better cell stability, more reproducible response, enabling continuous monitoring and greater mechanical strength (Mattiasson, 1997; Trevan and Mak, 1998). The usage of a thin layer of gel to immobilize the cells provides a fast diffusion of gases and short analysis time (Philp et al., 2003).
Cd toxicity tests on immobilized A. torulosa used in the biosensor construction showed that at lower concentrations of Cd, i.e., below approximately 8 mg L-1 of Cd there was no inhibition on photosynthetic oxygen release. Beyond this value, the inhibition of oxygen production occurred (Fig. 1). In fact, stimulation of oxygen release was observed at low concentration of Cd and the response is varied as the oxygen release can increase by up to 100% in one instance and only < 20% in another.
In photosynthetic organisms, Cd2+ can affect several metabolic activities in different ways. Some examples are photosynthesis and growth inhibition, binding onto chlorophylls, chlorosis and decrease in nutrient and water uptake (Clijsters and Assche, 1985; Krupa, 1999; Nies, 1999; Prasad and Strzalka, 1999, Clemens et al., 2002). Several studies in isolated chloroplasts have confirmed that Cd2+ affected photosynthesis in both the PS 1 and PS 2 (Atal et al., 1991; Siedlecka and Krupa, 1996; Pagliano et al., 2006).
A. torulosa is a photosynthetic organism. Although Cd can inhibit the activities of PS 1 and PS 2, results from this study showed that A. torulosa has mechanism to mediate the effect of Cd2+ especially at low concentration and after a short period of exposure. The mechanism appeared to accelerate the photosynthetic rate and produced more oxygen. The damage caused by low level of Cd2+ might initiate some biochemical processes that are related to the self-recovering mechanism from damage. The cyanobacteria cells used in the experiments were from the most active phase of growth and hence they may be capable of influence the Cd2+ transformation process in the cells (Krishnan et al., 2007).
However, the inhibition of photosynthesis began as soon as the Cd2+ concentration increased beyond the threshold value where the recovering mechanism no longer operational. Similar behaviour was also observed for the non-immobilized cells but in this case inhibition was observed until 1 mg L-1 of Cd after which stimulation occurred. At 5 mg L-1, the cells demonstrated inhibition again (Fig. 2).
The more susceptible of the non-immobilized cells to inhibition by Cd when compared with the immobilized cells is presumably attributed to the immobilized cells being protected from the pHEMA matrix that used to immobilize the organisms. This enables the organism to launch its self-recovery mechanism when Cd2+ is present.
The exposure duration of Cd2+ can have different effects on the toxicity behaviour. At short duration of exposure to Cd2+, an organism can adapt to higher Cd2+ concentrations by employing resistance mechanisms, such as expression of Cd2+-sequestering compounds (e.g., phytochelatins) or exporters capable of transporting Cd2+ out of cell (Nies, 1999). It has been observed that the cyanobacteria Trichodesmium erythraeum bloom when the Cd2+ concentration increased to 0.31 mg L-1 (Krishnan et al., 2007). For A. torulosa studied here, The IC50 for 30 min of exposure is 0.9 mg L-1. However, the IC50 for 15 min of exposure was not reached even with very high concentration of Cd2+ (Fig. 3).
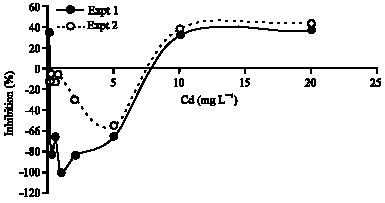 | |
| Fig. 1: | The response of the toxicity biosensor containing immobilized A. torulosa when exposed to various concentrations of Cd2+. Expt 1 and Expt 2 are duplicate experiments |
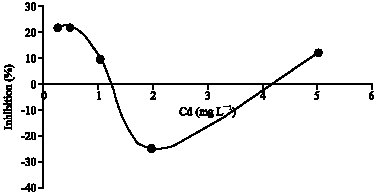 | |
| Fig. 2: | The response of non-immobilized (free) A. torulosa cells to various concentrations of Cd2+ demonstrate a response similar to the immobilized A. torulosa |
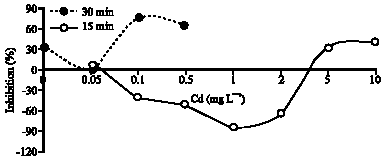 | |
| Fig. 3: | The effect of the length of exposure time on the inhibition behaviour of A. torulosa by Cd2+ |
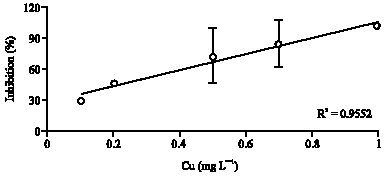 | |
| Fig. 4: | The inhibition pattern from various concentrations of Cu2+ on the toxicity biosensor containing immobilized A. torulosa. The reproducibility of the biosensor to Cu is indicated by the errors on the data |
The inhibition of oxygen observed throughout the Cd2+ concentrations used still demonstrated the cells attempted to recover after an initial inhibition by Cd2+ during the prolonged exposure to Cd2+ to 30 min.
The peculiar behaviour of Cd2+ toxicity on A. torulosa did not exhibited by other metal such as copper (Cu). Exposure of the biosensor to Cu2+ using the present experimental setup showed no increase of oxygen production by the cyanobacteria (Fig. 4). In fact inhibition of oxygen production was observed throughout when exposed to Cu2+and the response of the biosensor was linear with the concentrations of Cu2+. This is in agreement with previous finding by Chay et al. (2005) who used a same experimental setup to study Cu toxicity of A. torulosa.
In other species of algae or cyanobacteria, the presence of Cd2+ induced a significant decrease in activities of both PS 1 and PS 2, which led to inhibition of oxygen production. The level of inhibition on oxygen release yielded IC50 value of 5-10 mg L-1 in Chlamydomonas sp. and for Anabaena inaequalis, 1 mg L-1 of Cd2+ significantly inhibited the photosynthetic rate (Nagel and Voigt, 1995; Stratton and Corke, 1979). For Anabaena flos-aquae, the LD50 96 h for Cd2+ was reported to be 0.14 mg L-1 (Heng et al., 2004). Thus, even Anabaena species demonstrated different toxicity response to Cd2+. Based on the results observed here, some further studies may be useful to understand how the behaviour of this cyanobacteria species when under Cd2+ exposure.
CONCLUSION
The toxicity biosensor for the evaluation of Cd2+ toxicity on A. torulosa demonstrated that the Cd2+ toxicity behaviour of the cyanobacteria is peculiar where at lower concentration of Cd2+ (<8 mg L-1), stimulation of the photosynthetic activities occurred. This is in contrast to metal such Cu2+ toxicity on the same cyanobacteria where the stimulation phase was not observed even at much lower concentrations Cu2+ when compared to that of Cd2+. However, when the exposure time of the organism to Cd2+ was increased to 30 min, the biosensor registered a diminished effect of the stimulation phase and the inhibition effect became dominant.
REFERENCES
- Atal, N., P.P. Saradhi and P. Mohanty, 1991. Inhibition of the chloroplast photochemical reactions by treatment of wheat seedlings with low concentrations of cadmium: Analysis of electron transport activities and changes in fluorescence yield. Plant Cell Physiol., 32: 943-951.
CrossRefDirect Link - Bentley, A., A, Atkinson, J. Jezek and D.M. Rawson, 2001. Whole cell biosensors-electrochemical and optical approaches to ecotoxicity testing. Toxic. In Vitro, 15: 469-475.
Direct Link - Carpentier, R., C. Loranger, J. Chartrand and M. Purcell, 1991. Photoelectrochemical cell containing chloroplast membranes as a biosensor for phytotoxicity measurement. Anal. Chim. Acta, 249: 55-60.
CrossRef - Chay, T.C., S. Salmijah and L.Y. Heng, 2005. A copper toxicity biosensor using immobilized cyanobateria, Anabaena torulosa. Sensor Lett., 3: 49-54.
Direct Link - Clemens, S., M.G. Palmgren and U. Kramer, 2002. A long way ahead: Understanding and engineering plant metal accumulation. Trends Plant Sci., 7: 309-315.
CrossRef - Clijsters, H. and F. van Assche, 1985. Inhibition of photosynthesis by heavy metals. Photosynth. Res., 7: 31-40.
CrossRefDirect Link - Evans, G.P., M.G. Briers and D.M. Rawson, 1986. Can biosensors help to protect drinking water. Biosensors, 2: 287-300.
CrossRef - Heng, L.Y., K. Jusoh, C.H.M. Ling and M. Idris, 2004. Toxicity of single and combinations of lead and cadmium to the cyanobacteria Anabaena flos-aquae. Bull. Environ. Contam. Toxicol., 12: 373-379.
CrossRefDirect Link - Giardi, M.T., M. Kobilzek and J. Masojidek, 2001. Photosystem II-based biosensor for the detection of pollutants. Biosensors Bioelectron., 16: 1027-1033.
Direct Link - Krishnan, A.A., P.K. Krishnakumar and M. Rajagopalan, 2007. Trichodesmium erythraeum (Ehrenberg) bloom along the southwest coast of India (Arabic Sea) and its impact on trace metal concentration in seawater. Estuarine Coastal Shelf Sci., 71: 641-646.
Direct Link - Nies, D.H., 1999. Microbial heavy-metal resistance. Applied Microbiol. Biotechnol., 51: 730-750.
PubMed - Pagliano, C., M. Raviolo, F.D. Vecchia, R. Gabbrielli, C. Gonnelli, N. Rascio, R. Barbato and N.L. Rocca, 2006. Evidence for PSII donor-side damage and photoinhibition induced by cadmium treatment on rice (Oryza sativa L.). J. Photochem. Photobiol. B: Biol., 84: 70-78.
Direct Link - Philp, J.C., S. Balmand, E. Hajto, M.J. Bailey, S. Wiles, A.K. Lilley, J. Hajto and S.A. Dunbar, 2003. Whole cell immobilized biosensors for toxicity assessment of a wastewater treatment plant treating phenolics-containing waste. Anal. Chem. Acta, 487: 61-74.
Direct Link - Siedlecka, A. and Z. Krupa, 1996. Interaction between cadmium and iron and its effects on photosynthetic capacity of primary leaves of Phaseolus vulgaris. Plant Physiol. Biochem., 32: 833-841.
Direct Link - Van der Heever, J.A. and J.U. Grobbelaar, 1998. In vivo chlorophyll a fluorescence of Selenastrum capricornutum as a screening bioassay in toxicity studies. Arch. Environ. Contamination Toxicol., 35: 281-286.
CrossRef