
Lamiaa Mostafa Radwan
Department of Poultry Production, Faculty of Agriculture, Ain Shams University, Cairo, Egypt
LiveDNA: 20.8095
Mahmoud Yousef Mahrous
Department of Poultry Production, Faculty of Agriculture, Ain Shams University, Cairo, Egypt
Neima Koutb Alsenosy
Department of Genetics, Faculty of Agriculture, Ain Shams University, Cairo, Egypt
LiveDNA: 20.21959
Mohamed Ibrahim El Sabry
Department of Animal Production, Faculty of Agriculture, Cairo University, Giza, Egypt
LiveDNA: 20.21958
Asian Journal of Animal and Veterinary Advances
Year: 2018 | Volume: 13 | Issue: 4 | Page No.: 309-315







ABSTRACT
Background and Objective: The global warming is a hot topic for poultry scientists because of its negative influence on the quantity and the quality of poultry production. This study was conducted to investigate the influence of heat stress on Myogenin and uncoupling protein (UCP) genes expression and some carcass traits in two strains of broilers. Materials and Methods: This study was divided into two separate experiments, which were conducted at the same time. In the both experiments, chicks were exposed to 24°C (control) or 2 h/day at 2-7 days of age and against exposed to 40°C or 3 h/day at 33-34 days of age (treated), while chicks were fed a starter diet contained 23 and 25% crude protein in first and second experiments, respectively. At days 4 and 35 of age, samples were collected to determine the gene expression using qRT-PCR. At slaughter, carcass traits were measured. The Myogenin gene which responsible for the formation of muscle tissues is severely affected by heat stress which resulted in a depression in carcass and muscles weight. So, from a practical point of view it’s an important to compensate this accursed depression by raising the protein level in broiler diets. Results: The results showed that body weight, carcass, breast muscle relative weights of Cobb were significantly heavier compared to Ross. In general, the gene expression of both Myogenin and UCP genes were negatively influenced by heat stress, however, the increase in the protein (percentage) mitigated delirious heat stress effect, in both strains. Conclusion: Increasing the protein percentage may be used to alleviate the influence of heat stress and maintain the high yield of broilers.
PDF Abstract XML References Citation
Received: October 11, 2017;
Accepted: December 20, 2017;
Published: June 15, 2018
How to cite this article
Lamiaa Mostafa Radwan, Mahmoud Yousef Mahrous, Neima Koutb Alsenosy and Mohamed Ibrahim El Sabry, 2018. Interaction Between Heat Stress and Early Age on Muscle Development and Related to Gene Expression in Two Strains of Broilers Chickens. Asian Journal of Animal and Veterinary Advances, 13: 309-315.
DOI: 10.3923/ajava.2018.309.315
URL: https://scialert.net/abstract/?doi=ajava.2018.309.315
DOI: 10.3923/ajava.2018.309.315
URL: https://scialert.net/abstract/?doi=ajava.2018.309.315
INTRODUCTION
The global warming is a hot topic for poultry scientists because of its negative influence on the quantity and the quality of poultry production. The poultry meat considered as a main source of high nutritive value protein for human. In 2014, many reports indicated that the rate of increase in poultry meat consumption was about 1.6% and the share of poultry meat expenditure was 27.2% of the total expenditure of meat. Between 2015 and 2019, the overall growth of poultry meat commodities was expected to be 8.5% in the UK1, however the global warming, the increase in the ambient temperature, considered as a serious challenge that facing poultry industry. Inappropriate ambient temperature could cause significant economic losses in poultry farms, due to high mortality rate and depression in growth rate of birds, especially broilers2. Khan et al.3 mentioned that the estimated economic losses of poultry industry due to heat stress were about US $ 165 million/year. Previous studied have been showed that there were differences in tolerance of heat stress between different breeds and strain of chicken4. Therefore, information about the genes that were responsible for heat tolerance and the productivity under heat stress is needed to support breeding programs for enhancing the productivity of commercial broiler strains, those were grown in tropical and sub-tropical zones with poor management technologies. Breeding programs focused on productivity and meat quality parameters to produce a superior commercial broiler strains to meet the goal of producers (high revenue) and to reach consumers’ satisfaction. Multi genetic and environmental factors can affect poultry meat quality. For example, each breed or strain has its own genetic makeup that can affect the growth rate, feed efficacy and the amount and types of protein deposition into muscles5,6. Another factor affects poultry live performance and meat quality is the chicks house environment that could be manipulated by management practices such as choosing a proper type for housing, proper feeding program including level of protein, controlling ambient temperature and humidity. etc.7. Recently, gene expression became a reliable tool for poultry that could enhanced the efficiency of breeding programs8,9. For instant, gene expression can be involved in improving meat quality by giving information about the factors that affect protein and fat deposition10. Pinheiro et al.11 mentioned that muscle growth occurs through the process of hypertrophy myofiber which leading to an increase in fiber sizes through increasing protein synthesis. Ban et al.12 mentioned that there is a great importance to know the information about gene pathways to myogenic differentiation. It is recorded that both feeding programs and feed ingredients affected gene expression of Myogenin and Myostatin genes which responsible for improving broiler performance13-15. Its knowing that, the UCP gene was responsible for generating energy in muscles and hence its expected that, its function is affected by both heat stress and the level of protein in a fed ration. The Myogenin gene which controls the formation of muscle tissues was severely affected by heat stress that lead to a depression in carcass and muscles weight. Therefore, this study was conducted to investigate the impact of heat stress and related gene expression on muscle development and some carcass traits in two different strains of broilers.
MATERIALS AND METHODS
This study was carried out in Poultry Breeding Farm, Poultry Production Department, Faculty of Agriculture, Ain Shams University at summer season 2016. While, the lab work RNA extraction only was fulfilled at the Department of Genetic, Faculty of Agriculture, Ain Shams University, but the gene expression analysis by Real Time PCR was fulfilled at the Department of Poultry Production, Faculty of Agriculture, Ain Shams University.
A total of 1400 days-old chicks (half of them from Ross 308 strain and the another half from Cobb 500 strain). The chicks were reared in environmentally and hygienic controlled rooms at the Poultry Farm, Faculty of Agriculture, Ain Shams University. Feed and water were available ad libtium. At one day old, chicks from each strain were randomly divided into two separate experiments under the same environmental, managerial and hygienic conditions. In the 1st experiment, 700 chicks (half of them from Ross and Cobb strain) were fed a starter ration (23% protein and 3200 kcal kg–1), while in the 2nd experiment, chicks were fed a starter ration (25% protein and 3200 kcal kg–1) from day 1-7 and all chicks were fed grower ration (21% protein and 3000 kcal kg–1) from 8 days to till the end of the experiment (35 days of age). While, chicks in each experiment were divided into 2 groups. The control group was exposed to normal ambient temperature (brooded at 33°C at the birds' level during first 3 days of age and then the temperature was reduced gradually till room (24°C) temperature at 21st day of age until end experimental, while heat treated group (HT) treated group was exposed to 39°C or 2 h/day at 2-7 days of age and against exposed to 40°C or 3 h/day at 33-34 days of age.
Carcass measurement: At 35 days of age, all chicks were individually weighed and then slaughtered. Slaughtered chicks were re-weighed after bleeding, removed feather and viscera. The carcass was cut and weights were recorded for the breast, the percent breast cuts was calculated as follows:
Also thickness of major pectoral muscles was recorded (total number of 160 samples) using a digital micrometer and by recording the thickness of three pieces of major pectoral muscles from the equator and equidistant from each other on the same half of the major pectoral muscles.
Gene expression analysis: The RNA was extracted from forty eight major pectoral muscle samples using Trizol (Invitrogen, Carlsbad, CA). Myogenin estimated at 4 days of age only, while, UCP estimated at 35 days of age only. The isolated RNA was reversing transcribed for a cDNA using Maloney murine leukemia virus reverse transcriptase (Promega, Madison, WI). Total RNA extraction16 was separated electrophoretically on agarose gel with denaturing conditions in order to confirmed the integrity of ribosomal RNA bands17. Gene expression of Myogenin and UCP were assessed by one-step quantitative real-time (Thermo scientific verso SyBR Green 1-step qRT-PCR Kit Plus Rox Vial). The gene expression of Myogenin estimated, UCP were determined using real-time quantitative Polymerase Chain Reaction (RT-PCR) as described in Liu et al.18. The values were normalized with mRNA expression were expressed as the ratio of the α-actin mRNA values in arbitrary units. The RT-PCR to measure the expression of each gene was performed using the DyNAmo Hot Start. The primers used for each gene are listed in Table 1. The final PCR products were analyzed on a 1.5% agarose gel to check for amplification specificity. The mRNA levels, analyses were performed using GraphPad Prism version 6.00 for Windows (GraphPad Software, La Jolla, CA).
Statistical analysis: Data onto each experiment were statistically analyzed by two-way analysis of variance (ANOVA) with strain and treatment (heat stress) and their interaction (this is done separately for each experiment), using the General Linear Models (GLM) procedure using JMP Ver. 11 (SAS19). Duncan’s multiple range tests were used to separate means when significance differences exist.
| Table 1: | Primers used for each gene |
 | |
The statistical models used in this study were as follows:
Yijk = μ+Si+Tj+(S*T)ij+eijk
Where:| μ | = | Overall mean |
| Si | = | Strain effect |
| Tj | = | Treatment effect |
| S*Tij | = | Interaction between strain and treatment |
| eijk | = | Experimental error |
RESULTS AND DISCUSSION
Live body weight: It’s well known that the high temperature caused a significant reduction in both live body and carcass weights. In experiments 1 and 2, the interaction between strain and heat treatment (HT) showed an effect (p = 0.003) on the body weight being that Cobb strain groups received either high or normal CP (%) had heavier body weights compared to Ross ones. However, the heat stress negatively affected the final body weight, the results showed that live body weight of Cobb 500 strain was significantly heavier compared to Ross 308 in both experiments (Table 2). These differences in both body and carcass weights may be due to the high potential of genetic composition of Cobb strain to be responded to HT compared to Ross strain. Hristakieva et al.20 and Olawumi et al.21 recorded that Cobb 500 had a rapid growth rate and better feed conversion ratio compared to other studied commercial broiler strains. The chicks subjected to HT and fed lower CP level showed the lowest carcass weight (Table 2). In addition, the results from a Table 2 indicated that when birds fed a higher level of protein, this level help them to overcome the negative effect of heat stress on both body and carcass weight. This agreed with Kamran et al.22, who reported that the broilers fed low CP level diets recorded a lower growth rate and carcass composition. Results from Table 2 showed the interaction between genetic composition of the strain and temperature, being high temperature that affected live body weight and carcass weight. The Cobb strain recorded a greater breast relative weight compared to those of Ross in both experiments. Results showed that the ambient temperature had a significant effect on (p = 0.001) the relative weight of breast in both experiments (Table 3). Mello et al.23 recorded a decline in the breast yield due to birds exposure to 24 h temperature. Furthermore, they suggested that this negative effect of heat stress on breast yielded might be more due to, than adequate intake of nutrients and energy and reducing the synthesis and storage of glycogen, which is the main source of energy for the breast attributed the lower breast muscle development to the increased respiratory frequency due to heat stress.
| Table 2: | Effect of strain, treatment and their interaction on final body weight and carcass weight of the studied broiler strains |
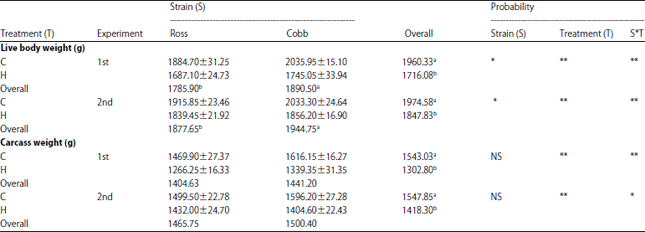 | |
C: Normal temperature, H: High temperature, 1st: Normal level protein (23%), 2nd: High level protein (25%). Means, within the same parameter, within the row, followed by different superscripts letters differ significantly (p<0.05). **p<0.01, *p<0.05 and NS: Not significant | |
| Table 3: | Effect of strain, treatment and their interaction on breast muscles characters of ross and cobb broiler strains |
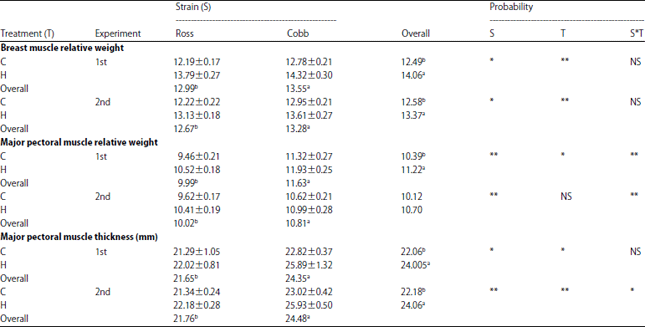 | |
C: Normal temperature, H: High temperature, 1st: Normal level protein (23%), 2nd: High level protein (25%). Means, within the same parameter, within the row, followed by different superscripts letters differ significantly (p<0.05). **p<0.01, *p<0.05 and NS: Not significant | |
This increase in respiration provides a higher activity of the breast muscles, which its development was hampered by the limited reserves of glycogen in the animal body. But these studies used intermittent heat stress from hatching to make an adaptation of negative high ambient temperature, the breast relative weighted and thickness had increased make more source storage of glycogen was used when exposed to high ambient temperatures. The strain had a significant effect on major pectoral muscle relative weight, being Cobb strain had greater relative weights compared to those of Ross (Table 3). The strain significantly impacted the relative weight of major pectoral muscles in both experiments. While, the effect of high temperature was significantly on the major pectoral muscle only in experiments to diet to feed 23% protein (experiment 1).
| Table 4: | The mRNA abundance of the Myogenin gene in Ross (R) and Cobb (C) lines subjected to normal and heat treatments in experiment 1 and 2, respectively significant (p<0.05). **p<0.01 and NS: Not significant |
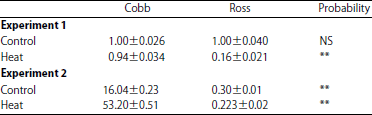 | |
| Table 5: | The mRNA abundance of the UCP gene in Ross (R) and Cobb (C) lines subjected to normal and heat treatments in experiment 1 and 2, respectively significant (p<0.05). **p<0.01 and NS: Not significant |
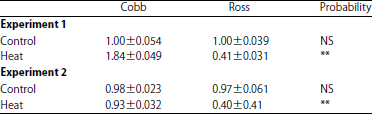 | |
Hence, it might be suggested that the gene composition could affect the muscle characteristics. Broiler strain significantly affected major muscles’ thickness, being Cobb strain had a thicker muscle compared to Ross one. Only the major pectoral muscle thickness only was affected in both experiments (Table 3). Also, there is an increase in thickness major pectoral muscle for birds raised under the effect of high ambient temperature. During the heat stress, the birds try to loss the excess heat through a limited pathway. Panting was considered as the main quick and efficient pathway to loss excess heat. Exposing chicks to ambient high temperature during the first 3 days of age caused an increase in insulin like growth factor-1 (IGF-1), number of satellite cells and thickness of myofibrils, in consequence weight and thickness of breast muscle were increased24. The results of experiment 1 showed that the percent of muscle protein was higher significantly for Cobb strain compared to Ross one. The results of experiment 2 showed that no significant differences due to strain, treatment and interaction. Meat quality and its chemical composition have high heritability and high response to selection25.
Gene expression: Myogenesis is a complicated and necessary process for muscles proliferation, which controlled by a group of genes such as: MyoD, Myogenin and Myostatin12. In the experiment 1, results showed that the Myogenin gene expression was similar in both strains under normal ambient temperature. However, a decrease in the mRNA transcript abundance of Myogenin gene was found in both Cobb and Ross strains due to exposure to high ambient temperature (Table 4). Gene expression of Cobb strain was highly significant (p = 0.0001) under high ambient temperature (Experiment 1) when compared with those of Ross one. It is noticed that there was a reduction in myogenin mRNA expression (which responsible for the formation of birds muscles) under high ambient temperature for both strains but the reduction was less in Cobb strain. In experiment 2, when birds fed a diet of 25% crude protein, its noticed that there were no any reduction occurred in Myogenin mRNA expression of Cobb strain. This trend was not the same for Ross strain, although, this reduction was lined when compared with their counterparts experimented 1. These findings might be explaining the occurred increase in the relative weight of breast muscle, either for its thickness and its protein contented in Cobb strain compared to Ross one (Table 3). This result was in strain on the finding of Gabriel et al.26, who found that a decreases in the abundance of myogenesis-related gene transcripts, which affected the skeletal muscle tissue formation of adverse temperature conditions during the embryonic stage. The uncoupling protein gene (UCP) was responsible for a transporter of the inner mitochondrial membrane27, which is known to uncouple respiration from ATP synthesis by short circuit the inward proton flow, resulting in heat production4. Under high temperature the chicken loss the excess heat of panting. Panting process requires more energy ATP to make pectoral muscle increased respiratory rate so under high temperature the UCP mRNA expression increase. As for the UCP gene expression, the results showed that there were no significant differences detected between the two strains under the circumstances of experiment 1. However, an increase in mRNA transcription abundance of UCP gene was found in Cobb and a depression in the mRNA transcription abundance of UCP gene was found only in the Ross strain due to exposure to high ambient temperature. However, the mRNA transcription abundance of Myogenin gene was significantly lower (p<0.05) in Ross strain compared to Copp one (Table 5). In experiment 2, the results of UCP mRNA expression were significant for Cobb and Ross strains reared under normal temperature were similar, whereas Cobb strain showed higher gene expression than Ross strain (Table 5). Feng et al.28 found an increase in the mitochondrial H2O2 production and MDA concentrations on breast muscle had under high temperature conditions compared to the normal ambient temperature control group, in chicken. Tan et al.29 observed that mitochondrial superoxide radical production of chicken skeletal muscle increases in acute heat stress, have indicated that reactive oxygen species (ROS) influence the calcium released channel or Ca2+-ATPase activity. The UCP gene expression results may played an important role in adjusting metabolic heat production to maintain stable body temperatures under different environments30, which is may be important to the chicken productivity and meat quality. Broiler under heat stress was significantly enhanced superoxide production in heat stress-treated skeletal muscle mitochondria and changes DNA, proteins, lipid and other biological molecular31. The UCP gene plays roles in important β-oxidation and thermogenesis this gene was specifically expressed in chicken muscle, the main site of thermogenesis and whole body β-oxidation4.
CONCLUSION
The modern broiler strains may show different performances due to their genetic composition. Both experiments appear that Cobb strain may have better productive performance under heat stress compared to the Ross strain. While the second experiment refer to that high ambient temperature negatively affect gene expression of Myogenin but this deleterious effect could be mitigated by increasing the protein level in chicken diets.
SIGNIFICANCE STATEMENT
This study discovers the influence of heat stress on genes expression for Myogenin and uncoupling protein (UCP) and some carcass traits in two strains of broilers that can be beneficial for avoiding negative influence for hot ambient temperature on the quantity and the quality of poultry production. The modern broiler strains may show different performances due to their genetic composition. Cobb strain may have better productive performance under heat stress compared to the Ross strain. Thus a new theory on increasing the protein percentage may be used to alleviate the influence of heat stress and maintain the high yield of broilers.
REFERENCES
- Dalolio, F.S., L.F.T. Albino, H.J. Lima, J.N.D. Silva and J. Moreira, 2015. Heat stress and vitamin E in diets for broilers as a mitigating measure. Acta Scientiarum. Anim. Sci., 37: 419-427.
Direct Link - Khan, R.U., S. Naz, Z. Nikousefat, V. Tufarelli, M. Javdani, N. Rana and V. Laudadio, 2011. Effect of vitamin E in heat-stressed poultry. World's Poult. Sci. J., 67: 469-478.
CrossRefDirect Link - Dridi, S., E. Decuypere and J. Buyse, 2013. Cerulenin upregulates heat shock protein-70 gene expression in chicken muscle. Poult. Sci., 92: 2745-2753.
CrossRefDirect Link - Malila, Y., Y. Srimarut and U. Juthawut, 2015. Monitoring of chicken RNA integrity as a function of prolonged postmortem duration. Asian-Aust. J. Anim. Sci., 28: 1649-1656.
Direct Link - Koppelkamm, A., B. Vennemann, S. Lutz-Bonengel, T. Fracasso and M. Vennemann, 2011. RNA integrity in post-mortem samples: Influencing parameters and implications on RT-qPCR assays. Int. J. Legal Med., 125: 573-580.
CrossRefDirect Link - Wang, Q., L. Ning, D. Xuemei, L. Zhengxing, L. Hui and W. Changxin, 2001. Single nucleotide polymorphism analysis on chicken extracelluar fatty acid binding protein gene and its associations with fattiness trait. Sci. China, 4: 429-434.
CrossRef - Lorentz, L.H., L.D.G. Gaya, R. Lunedo, J.B.S. Ferraz, F.M.D. Rezende and T.M. Filho, 2011. Production and body composition traits of broilers in relation to breast weight evaluated by path analysis. Scientia Agricola, 68: 320-325.
Direct Link - De Genova Gaya, L., 2013. Genetic variability in ultrasound records of breast muscle in a broiler breeding program. Natural Sci., Vol. 5.
CrossRefDirect Link - Malila, Y., R.J. Tempelman, K.R.B. Sporer, C.W. Ernst, S.G. Velleman, K.M. Reed and G.M. Strasburg, 2013. Differential gene expression between normal and pale, soft and exudative turkey meat. Poult. Sci., 92: 1621-1633.
CrossRefDirect Link - Pinheiro, C.H.d.J., L.R. Silveira, R.T. Nachbar, K.F. Vitzel and R. Curi, 2010. Regulation of glycolysis and expression of glucose metabolism-related genes by reactive oxygen species in contracting skeletal muscle cells. Free Radic. Biol. Med., 48: 953-960.
CrossRef - Ban, Q., Y. Liang, Z. Zhao, X. Liu and Q. Li, 2013. Differential expression levels of genes related to myogenesis during embryogenesis of quail and chicken. Pak. Vet. J., 33: 317-320.
Direct Link - Yasin, M., A. Asghar, F.M. Anjum, M.S. Butt and M.I. Khan et al., 2012. Oxidative stability enhancement of broiler bird meats with α-lipoic acid and α-tocopherol acetate supplemented feed. Food Chem., 131: 768-773.
CrossRefDirect Link - Demonty, I., R.T. Ras, H.C. van der Knaap, L. Meijer, P.L. Zock, J.M. Geleijnse and E.A. Trautwein, 2013. The effect of plant sterols on serum triglyceride concentrations is dependent on baseline concentrations: A pooled analysis of 12 randomised controlled trials. Eur. J. Nutr., 52: 153-160.
CrossRefDirect Link - Naji, T.A.A., I. Amadou, R.Y. Zhao, X. Tang, Y.H. Shi and G.W. Le, 2014. Effects of phytosterol in feed on growth and related gene expression in muscles of broiler chickens. Trop. J. Pharm. Res., 13: 9-16.
Direct Link - Chomczynski, P. and N. Sacchi, 1987. Single-step method of RNA isolation by acid guanidium tiocyanate phenol chloroform extraction. Anal. Biochem., 162: 156-159.
Direct Link - Liu, H.H., J.W. Wang, X. Chen, R.P. Zhang and H.Y. Yu et al., 2011. In ovo administration of rhIGF-1 to duck eggs affects the expression of myogenic transcription factors and muscle mass during late embryo development. J. Applied Physiol., 111: 1789-1797.
CrossRef - Hristakieva, P., N. Mincheva, M. Oblakova, M. Lalev and I. Ivanova, 2014. Effect of genotype on production traits in broiler chickens. Slovak J. Anim. Sci., 47: 19-24.
Direct Link - Olawumi, S.O., S. Fajemilehin and S.S. Fagbuaro, 2012. Genotype × sex interaction effects on carcass traits of three strains of commercial broiler chickens. J. World's Poult. Res., 2: 21-24.
Direct Link - Kamran, Z., M. Sarwar, M. Nisa, M.A. Nadeem, S. Mahmood, M.E. Babar and S. Ahmed, 2008. Effect of low-protein diets having constant energy-to-protein ratio on performance and carcass characteristics of broiler chickens from one to thirty-five days of age. Poult. Sci., 87: 468-474.
CrossRefDirect Link - Mello, J.L.M., M.M. Boiago, A. Giampietro-Ganeco, M.P. Berton and L.D.C. Vieira et al., 2015. Periods of heat stress during the growing affects negatively the performance and carcass yield of broilers. Arch. Zoot., 64: 339-345.
Direct Link - Fresard, L., M. Morisson, J.M. Brun, A. Collin, B. Pain, F. Minvielle and F. Pitel, 2013. Epigenetics and phenotypic variability: Some interesting insights from birds. Genet. Select. Evol., Vol. 45.
CrossRefDirect Link - Le Bihan-Duval, E., M. Debut, C. Berri, N. Sellier, V. Sante-Lhoutellier, Y. Jego and C. Beaumont, 2008. Chicken meat quality: Genetic variability and relationship with growth and muscle characteristics. BMC Genet., Vol. 9.
CrossRefDirect Link - Gabriel, J.E., H.J. Alves, M.F. Do Rosario, A. Secatto, L.L. Coutinho and M. Macari, 2011. Abundance of MyoD and myostatin transcripts in chicken embryos submitted to distinct incubation temperatures and timing exposures. Braz. J. Biol., 71: 563-564.
PubMedDirect Link - Rey, B., D. Roussel, C. Romestaing, M. Belouze and J.L. Rouanet et al., 2010. Up-regulation of avian uncoupling protein in cold-acclimated and hyperthyroid ducklings prevents reactive oxygen species production by skeletal muscle mitochondria. BMC Physiol., Vol. 10.
CrossRefDirect Link - Feng, J., M. Zhang, S. Zheng, P. Xie and A. Ma, 2008. Effects of high temperature on multiple parameters of broilers in vitro and in vivo. Poult. Sci., 87: 2133-2139.
CrossRefDirect Link - Tan, G.Y., L. Yang, Y.Q. Fu, J.H. Feng and M.H. Zhang, 2010. Effects of different acute high ambient temperatures on function of hepatic mitochondrial respiration, antioxidative enzymes and oxidative injury in broiler chickens. Poult. Sci., 89: 115-122.
CrossRefDirect Link - Del Vesco, A.P. and E. Gasparino, 2013. Production of reactive oxygen species, gene expression, and enzymatic activity in quail subjected to acute heat stress. J. Anim. Sci., 9: 582-587.
PubMedDirect Link - Ali, M.S., G.H. Kang and S.T. Joo, 2008. A review: Influences of pre-slaughter stress on poultry meat quality. Asian-Aust. J. Anim. Sci., 21: 912-916.
CrossRefDirect Link