
Janine Donaldson
School of Physiology, Faculty of Health Sciences, University of the Witwatersrand, 7 York Road, 2193 Parktown, Johannesburg, South Africa
LiveDNA: 27.16892
Michael Taurai Madziva
School of Physiology, Faculty of Health Sciences, University of the Witwatersrand, 7 York Road, 2193 Parktown, Johannesburg, South Africa
Kennedy Honey Erlwanger
School of Physiology, Faculty of Health Sciences, University of the Witwatersrand, 7 York Road, 2193 Parktown, Johannesburg, South Africa
Asian Journal of Animal and Veterinary Advances
Year: 2017 | Volume: 12 | Issue: 6 | Page No.: 274-283







ABSTRACT
Background and Objective: Fats are often added to poultry diets to increase growth performance and productivity. The effects of increased dietary fat consumption in guinea fowl and muscovy ducks are unknown, which may have consequences for poultry production costs and the formulation of species-specific diets. The current study investigated the effects of a diet rich in saturated fatty acids on glucose tolerance, serum metabolic health parameters and the liver of guinea fowl and muscovy ducks. Materials and Methods: Twenty four guinea fowl and 24 muscovy ducks were divided into two groups/species and fed either a standard or high-fat diet (enriched with 20% palm oil and 2% lard) over 8 weeks. After 4 and 8 weeks on the diets, six birds from each group were fasted overnight, subjected to an oral glucose tolerance test, re-fed for 72 h and then euthanized. Blood and liver samples were collected. Means between birds in the standard and high-fat diet groups, within each bird species at each time point were compared with a student’s t-test (p<0.05), using Graphpad 5 prism software. Results: The high-fat diet had no significant effects on the glucose tolerance parameters investigated or on the liver lipid content in both bird species. Significantly increased serum cholesterol concentrations were observed in birds that received the high-fat diet versus those that received the standard diet, in both bird species. Conclusion: The metabolic health of the birds was maintained following consumption of the high-fat diet and thus the diet would be safe for use during production of these alternative poultry species.
PDF Abstract XML References Citation
Received: August 02, 2017;
Accepted: September 09, 2017;
Published: October 15, 2017
How to cite this article
Janine Donaldson, Michael Taurai Madziva and Kennedy Honey Erlwanger, 2017. Metabolic Health of Guinea Fowl and Muscovy Ducks Fed a High-Fat Diet. Asian Journal of Animal and Veterinary Advances, 12: 274-283.
DOI: 10.3923/ajava.2017.274.283
URL: https://scialert.net/abstract/?doi=ajava.2017.274.283
DOI: 10.3923/ajava.2017.274.283
URL: https://scialert.net/abstract/?doi=ajava.2017.274.283
INTRODUCTION
Consumption of a high-fat diet (HFD) results in the development of metabolic disturbances in mammals, including glucose intolerance, obesity, insulin resistance and fat accumulation within the liver1,2. There is, however, a scarcity of data regarding increased levels of dietary fat consumption in Avian species (specifically the guinea fowl and muscovy duck). Most of the research which has been done has focused on chickens, which may have consequences in terms of poultry production costs and the formulation of diets specific to other poultry species.
Besides chickens, poultry species such as guinea fowl and muscovy ducks have become popular as a source of protein for human consumption. Guinea fowl are advantageous over chickens in terms of poultry production and meat quality. Guinea fowl are less susceptible than chickens to various poultry diseases, are associated with lower production costs3 and are able to adapt to various environmental conditions, making them relatively easy to rear4. Guinea fowl meat has a high protein and low fat content, making it of high quality and attractive to consumers compared to chicken meat3. Duck meat has also become popular as a source of protein as it has some of the characteristics of red meat, such as the high levels of phospholipids, together with the nutritional properties of poultry meat, such as high levels of unsaturated fatty acids. Ducks are also relatively hardy species which allows them to adapt to different environments5.
Poultry diets are often supplemented with fat in the form of animal fat or vegetable oils, as a concentrated source of metabolizable energy, increasing the energy available to the birds in an attempt to increase growth performance and productivity6,7. Improving the consistency and tastiness of the feed8 and at the same time decreasing feed intake and increasing feed efficiency9 are further rationales for the use of a HFD in poultry production practices. Birds have a high capacity for lipid absorption and biosynthesis10 and exogenous lipids are absorbed directly from the small intestine into the portal blood and transported to the liver for further metabolism and subsequent tissue deposition11. The degree of fatty acid saturation and chain length of added dietary fatty acids have been shown to affect the digestibility of the fat7. Saturated fatty acids (SFA) are preferentially stored and polyunsaturated fatty acids (PUFA) are preferentially oxidised for energy12. In humans, rats, hamsters and chickens, increased consumption of fat has been shown to cause an increased flux of free fatty acids to the liver, which eventually leads to increased triglyceride synthesis, impaired β-oxidation and reduced clearance of very low density lipoproteins from the liver and ultimately, the development of hepatic steatosis13. In addition, consumption of a HFD has also been linked to the development of obesity, glucose intolerance and insulin resistance14. Following HFD consumption, increased circulating free-fatty acid levels impair insulin function, resulting in insulin resistance, which in turn affects glucose metabolism homeostasis15.
Since it is common to formulate poultry diets with added fat and there is a dearth of literature concerning the effects of increased fat consumption on the development of metabolic disturbances such as glucose intolerance and hepatic steatosis, specifically in guinea fowl and muscovy duck (which are becoming increasingly popular sources of protein for human consumption), the objective of the current study was to investigate the long term effects of a HFD, rich in SFA, on glucose tolerance, serum metabolic health markers and the liver lipid content of guinea fowl and muscovy ducks.
MATERIALS AND METHODS
Ethical approval and statement of animal rights: The study was approved by the University of the Witwatersrand Animal Ethics Screening Committee (Ethics clearance number: 2011/08/2A). All applicable international, national and/or institutional guidelines for the care and use of animals were followed when carrying out the present research. All procedures performed in the present study were in accordance with the ethical standards of the University of the Witwatersrand animal ethics screening committee. Data collection was performed during 2012-2013.
Animals and housing: Twenty four (mixed group of males and females), 4 week old guinea fowl (Numida meleagris) (452.4±91.2 g) and twenty four (mixed group of males and females), 4 week old muscovy ducks (Cairina moschata) (671.4±194.0 g) were used in the study. Each species of bird was randomly divided into two groups of 12 birds each and allocated to either a standard diet (STD) group which received the standard, commercial poultry feed (Epol®, Centurion, Pretoria, South Africa) or a high-fat diet (HFD) group which received the commercial poultry feed enriched with palm oil (SupaCrisp, Super Olein Palm Oil, Felda Bridge Africa (PTY) Ltd, Johannesburg, South Africa) at 20% of the mass of the feed and lard (Norbert’s German butchery, Wilropark, Roodepoort, South Africa) at 2% of the mass of the feed (in total 22% added dietary fat, on a weight/weight basis). The birds were fed their respective diets ad libitum, with free access to water for the duration of the experimental period. All birds were handled for a few minutes, several times/day, so that they could get used to being handled in an attempt to prevent handling stress hyperglycaemia when the interventions took place16. All birds were housed in their respective groups in the animal unit of the Central Animal Service, Faculty of Health Sciences, University of the Witwatersrand. Lighting was restricted to 12 h in each 24 h period, lights on from 06:00. To provide environmental enrichment of the bird cages, the guinea fowl were provided with perching logs and the muscovy ducks were provided with paddle pools filled with water.
General experimental procedure: Following a 2 week adaptation period, to familiarise the birds with the housing, handling and feeding conditions, the birds were fed their respective diets for 8 weeks. Body mass was measured weekly and weight gain was calculated (growth performance data was presented in a previous publication by our lab) by Donaldson et al.17. Every four weeks, six birds from each group (STD and HFD) were fasted for approximately 15 h overnight and then subjected to an oral glucose tolerance test (OGTT) as detailed below. The OGTT served as a measure of the glucose absorption capacities and glucose homeostasis of the birds18. Following the OGTT, the birds were re-fed and allowed to recover for 72 h and then euthanized using an anaesthetic overdose of Pentobarbital (Eutha-naze, Centaur Labs, South Africa) (200 mg kg–1), administered via the wing vein. Blood samples were collected by cardiac puncture (using 20 G hypodermic needles) into vacutainer tubes (Vacuette, Greiner Bio-One, Kremsmunster, Austria), for determination of serum biochemistry. The livers were excised, weighed and samples processed for lipid content. Liver lipid yield (as a % of liver tissue sample mass) was calculated.
Specific experimental procedures
Oral glucose tolerance tests: Following an overnight fast, fasting blood glucose concentrations of blood samples collected via venipuncture of the wing vein, after 4 and 8 weeks of STD or HFD feeding, were measured using a glucometer (Ascensia Elite™ Blood glucose meter, Bayer Corporation, Mishawaka, USA). Prior to the venipuncture, the area surrounding the wing vein was clipped of its feathers and sterilized with alcohol-soaked gauze swabs. Following the fasting blood glucose level measurement, birds were then administered a single dose of glucose solution (5g kg–1, 50% w/v glucose solution: Sigma-Aldrich®, Seelze, Germany) orally, via orogastric intubation into the crop. Blood glucose concentrations (mmol L–1) were then determined (using a glucometer) at fixed time intervals (15, 30, 60 and 120 min following glucose administration)19. Glucose tolerance curves, for both species of birds in the STD and HFD groups were then constructed. Baseline blood glucose, peak blood glucose and blood glucose concentrations 2 h following oral glucose administration were then determined. Area under the glucose curve (AUC) was also determined.
Serum metabolic health markers: The blood samples collected at euthanasia, were centrifuged (Sorvall RT 6000 B, Du Pont, Hertfordshire, United kingdom) at 370 rpm and 22°C, for 15 min. The serum was then collected and serum uric acid, total protein, albumin, aspartate transaminase (AST), total bilirubin, cholesterol and triglyceride concentrations were determined using an IDEXX Vetlab Analysis Machine (IDEXX Laboratories, Westbrook, Maine) according to manufacturer’s instructions. Serum cholesterol and triglyceride concentration data were presented in a previous publication by lab, Donaldson et al.17.
Liver lipid content: Liver lipid content analysis was only performed using liver samples, from half of the birds after 4 and 8 weeks of either STD or HFD feeding. Lipid extraction was performed as described by Bligh and Dyer20. Briefly, liver samples (approximately five grams) were placed into a chloroform:methanol (2:1) solution, overnight at 4°C. The samples were then filtered through filter paper (Albert®, Pore 7-11, Size 185 mm) and 30 mL of 0.9% saline was added. The samples were then gently mixed by inverting and allowed to stand overnight at 4°C, to allow separation into two phases. The bottom (chloroform) phase was then collected and reduced to dryness under vacuum, at 37°C, using a water bath (Labex®, Krugersdorp, South Africa) and then made up to 20 mL with chloroform. An aliquot of 2 mL of the extracts was then placed into dried, pre-weighed vials and re-dried at 50°C for 30 min, cooled and then reweighed to determine the lipid content.
Data analysis: All data were expressed as mean (SD). The data were analysed and plotted using Graphpad 5 Prism software (Graph-pad Software Inc, San Diego, USA). p<0.05 was considered significant.
A student’s t-test was used to assess differences in OGTT parameters, serum parameters and liver lipid content between birds in the STD and HFD groups, within each bird species, at each time point (following 4 and 8 weeks of feeding). Bartlett’s test statistic for equal variance was used to define OGTT parameters with significantly different variance (peak glucose concentrations and glucose concentrations 2 h following administration of the glucose load). The transformation used to correct for the unequal variance was the inverse function.
One guinea fowl and one muscovy duck fell ill during the experimental period and both were excluded from the study. Thus, for most of the statistical analyses (unless otherwise stated), n = 6 for both species of bird, at all time points, except for the guinea fowl on the HFD at 8 weeks and the muscovy ducks on the STD at 8 weeks, where n = 5. For the analysis of the liver lipid content, n = 3 for both species of bird, at all time points, except for the muscovy ducks on the STD at 8 weeks, where n = 2.
RESULTS
Guinea fowl
Oral glucose tolerance tests: The glucose tolerance curves for guinea fowl after 4 (a) and 8 (b) weeks of STD or HFD feeding are shown in Fig. 1. Fasting blood glucose concentrations were not significantly different between STD and HFD groups after 4 and 8 weeks of feeding. There were also no significant differences in fasting blood glucose concentrations within the single dietary groups across the two different time points assessed (p>0.05). Blood glucose concentrations peaked at 15 min following gavage in guinea fowl on the STD versus 30 min for those on the HFD following 4 weeks of feeding and at 30 min following gavage for guinea fowl in the STD and HFD groups after 8 weeks of feeding. All peaks were significantly higher than fasting blood glucose concentrations (p<0.01) for both dietary groups at both time points, except for that of the guinea fowl on the HFD, following 8 weeks of feeding, which was not significantly different from fasting blood glucose concentrations. The peak blood glucose concentrations for both dietary groups were not significantly different from one another after 4 and 8 weeks of feeding. Peak blood glucose concentrations were also not significantly different across the two different time points assessed, within the single dietary groups (p>0.05). Blood glucose concentrations returned to normal 60 min after gavage in guinea fowl in both the STD and HFD groups at all time points assessed. There were no significant differences in AUC between birds in the STD and HFD groups, following either 4 or 8 weeks of STD or HFD feeding, neither were there any significant differences (p>0.05) observed in AUC across the two different time points assessed, within the individual STD and HFD groups (Fig. 2).
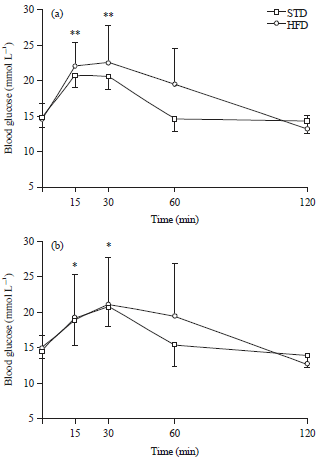 | |
| Fig. 1(a-b): | Glucose tolerance curves (Mean±SD) of guinea fowl after 4 (a) and 8 (b) weeks of standard diet (STD) or high-fat diet (HFD) (20% palm oil and 2% lard) feeding |
**p<0.01 significance when comparing blood glucose concentrations at 15 and 30 min post-gavage to fasting blood glucose concentrations in guinea fowl in the STD and HFD groups, following 4 weeks of feeding. *p<0.05 significance when comparing blood glucose concentrations at 15 and 30 min post-gavage to fasting blood glucose concentrations in guinea fowl in the STD and HFD group, following 8 weeks of feeding | |
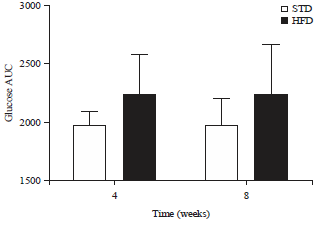 | |
| Fig. 2: | Area under the glucose curve (Mean±SD) for guinea fowl after 4 (a) and 8 (b) weeks of standard diet (STD) or high-fat diet (HFD) (20% palm oil and 2% lard) feeding |
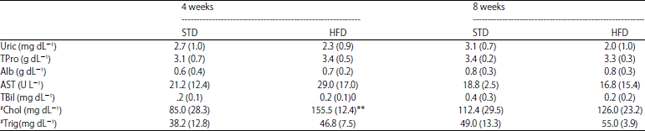
| Table 1: | Guinea fowl (Numida meleagris) serum parameters after 4 and 8 weeks of either standard diet or high-fat diet feeding |
 | |
Data represented as mean (SD), **p<0.01 HFD compared to STD at specific time point, STD = Standard diet, HFD = High-fat diet, Uric = Uric acid, TPro = Total protein, Alb = Albumin, AST = Aspartate aminotransferase, TBil = Total bilirubin, Chol= Cholesterol, Trig = Triglycerides, n = 6 for both STD and HFD groups at 4 weeks and n = 6 for STD group at 8 weeks and n = 5 for HFD group at 8 weeks, #Serum cholesterol and triglyceride data has been referred to in a previous publication by our lab, Donaldson et al.17 | |
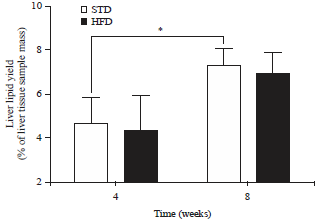 | |
| Fig. 3: | Liver lipid yield (expressed as a % of liver tissue sample mass, Mean±SD) of guinea fowl after 4 and 8 weeks of standard diet (STD) or high-fat diet (HFD) (20% palm oil and 2% lard) feeding |
| *p<0.05 significance when comparing liver lipid yield of guinea fowl on the STD after 4 weeks of feeding and guinea fowl on the STD after 8 weeks of feeding | |
Serum biochemistry: Serum parameters for guinea fowl in both the STD and HFD groups are presented in Table 1. No significant differences were observed in serum uric acid, total protein, albumin, AST, total bilirubin and triglyceride concentrations between birds on the STD and HFD, after either 4 or 8 weeks of feeding. Serum cholesterol concentrations were significantly higher (p<0.01) after 4 and 8 weeks of feeding in guinea fowl on the HFD compared to those on the STD diet. No significant differences were observed in serum uric acid, total protein, albumin, total bilirubin, triglyceride and cholesterol concentrations when compared across the two different time points, within the individual STD and HFD groups. Serum AST concentrations were significantly higher (p<0.05) following 4 weeks of STD feeding compared to that observed following 8 weeks of STD feeding. No significant differences in serum AST concentrations were observed across the two different time points in the HFD group.
Liver lipid content: No significant differences were observed between the liver lipid yield (as a % of liver tissue sample mass) of guinea fowl on the STD compared to those on the HFD, after 4 and 8 weeks feeding (Fig. 3). When comparing liver lipid yield between the two time points (i.e., after 4 and 8 weeks of feeding), within individual diet groups, the birds in the STD group had significantly increased (p<0.05) liver lipid yield after 8 weeks of feeding compared to after 4 weeks of STD feeding. There were no significant differences observed between birds in the HFD group after 4 weeks of feeding and those in the HFD group after 8 weeks of feeding.
Muscovy ducks
Oral glucose tolerance tests: The glucose tolerance curves for muscovy ducks after 4 (a) and 8 (b) weeks of STD or HFD feeding are shown in Fig. 4. Fasting blood glucose concentrations were not significantly different between STD or HFD groups or between the two different time points assessed, within a single dietary group. Peak blood glucose concentrations were reached 30 min after administration of the glucose load, following 4 and 8 weeks of either STD or HFD feeding. All peaks were significantly higher (p<0.01) than fasting blood glucose concentrations in both dietary groups, after 4 and 8 weeks of feeding and were not significantly different between the STD and HFD groups. Peak blood glucose concentrations were also not significantly different across the two time points assessed, within the single dietary groups. Blood glucose concentrations only returned to normal 120 min after gavage, at all time points assessed, for birds in both the STD and HFD groups. The AUC for muscovy ducks in the HFD group was significantly larger (p<0.05) than that of the birds in the STD group, after 4 weeks of feeding. When comparing the AUC across the two different time points assessed, within the individual diet groups, no significant differences were observed (Fig. 5).
| Table 2: | Muscovy ducks (Cairina moschata) serum parameters after 4 and 8 weeks of either standard diet or high-fat diet feeding |
 | |
Data represented as mean (SD), **p<0.01 HFD compared to STD at specific time point, STD = Standard diet, HFD = High-fat diet, Uric = Uric acid, Tpro = Total protein, Alb = Albumin, AST = Aspartate aminotransferase, TBil = Total bilirubin, Chol = Cholesterol, Trig = Triglycerides, n = 6 for both STD and HFD groups at 4 weeks (except for Trig, where n = 5 in STD group) and n = 5 for STD groups at 8 weeks (except for AST, where n = 4 in STD group) and n = 6 for HFD groups at 8 weeks (except for AST, where n = 4 in HFD group), #Serum cholesterol and triglyceride data has been referred to in a previous publication by our lab, Donaldson et al.17 | |
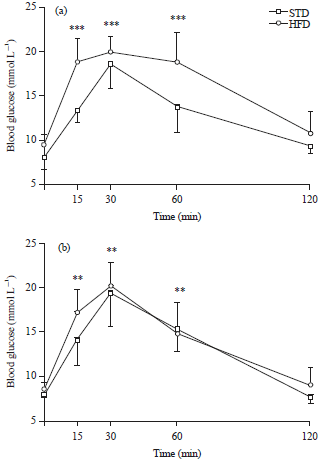 | |
| Fig. 4: | Glucose tolerance curves (Mean±SD) of muscovy ducks after 4 (a) and 8 (b) weeks of standard diet (STD) or high-fat diet (HFD) (20% palm oil and 2% lard) feeding |
***p<0.001 and **p<0.01 significance when comparing blood glucose concentrations at 15, 30 and 60 min post-gavage to fasting blood glucose concentrations in muscovy ducks in the STD and HFD groups, following 4 weeks of feeding and in muscovy ducks in the STD and HFD groups, following 8 weeks of feeding, respectively | |
Serum biochemistry: Serum parameters for muscovy duck in both the STD and HFD groups are presented in Table 2. No significant differences were observed in serum uric acid, total protein, albumin, AST, total bilirubin or triglyceride concentrations in the muscovy ducks in both the STD and HFD groups, following either 4 or 8 weeks of feeding.
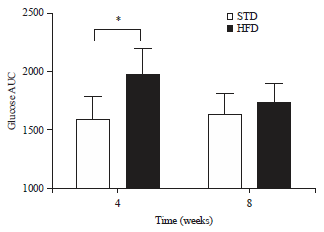 | |
| Fig. 5: | Area under the glucose curve (Mean±SD) for after 4 (a) and 8 (b) weeks of standard diet (STD) or high-fat diet (HFD) (20% palm oil and 2% lard) feeding |
| *p<0.05 significance when comparing AUC of muscovy ducks on the STD and HFD after 4 weeks of feeding | |
Serum cholesterol concentrations were significantly higher (p<0.001) in birds in the HFD group compared to those in the STD group after 4 weeks of feeding, however no significant differences in serum cholesterol concentrations were observed between the two dietary groups after 8 weeks of feeding. When comparing each serum parameter across the two different time points, within the individual STD and HFD groups, no significant differences were observed in serum uric acid, total protein, albumin, AST and total bilirubin concentrations. Serum triglyceride concentrations were significantly higher (p<0.05) following 8 weeks of HFD feeding compared to that observed following 4 weeks of HFD feeding. Serum cholesterol concentrations were significantly higher (p<0.05) following 4 weeks of HFD feeding compared to that observed following 8 weeks of HFD feeding. No significant differences in serum triglyceride and serum cholesterol concentrations were observed across the two different time points within the STD group.
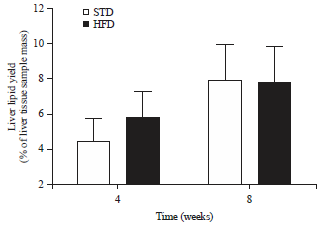 | |
| Fig. 6: | Liver lipid yield (expressed as a % of liver tissue sample mass, Mean±SD) of muscovy ducks after 4 and 8 weeks of standard diet (STD) or high-fat diet (HFD) (20% palm oil and 2% lard) feeding |
Liver lipid content: No significant differences were observed between the liver lipid yield (as a % of liver tissue sample mass) of birds on the STD compared to those on the HFD, after 4 and 8 weeks feeding (Fig. 6). No significant differences in liver lipid yield were observed within the individual STD or HFD groups, after either 4 or 8 weeks of STD or HFD feeding.
DISCUSSION
High-fat diet feeding in mammals21,22 and some birds has been associated with alterations in glucose tolerance. Consumption of saturated fatty acids (SFA) and monounsaturated fatty acids (MUFA), such as palm oil and lard, causes an imbalance in the plasma concentrations of glucose and insulin23,24. Crespo and Esteve-Garcia24 observed increased plasma insulin concentrations, that were not accompanied by a depletion of plasma glucose, in broilers fed diets containing 10% tallow or olive oil versus those fed diets containing 10% sunflower or linseed oil, suggesting the development of insulin resistance in the birds fed diets rich in SFA and MUFA compared to those fed diets rich in PUFA24. Since avian insulin plays a role in promoting glucose uptake from the bloodstream25, one would expect the HFD-induced changes in the insulin/glucose balance and the development of insulin resistance in the birds fed the diets rich in SFA and MUFA, to have an effect on glucose tolerance. Newman et al.23 observed no significant differences in plasma glucose responses to an intravenous glucose tolerance test in male broiler chickens fed a HFD composed of 80 g kg–1 tallow, fish oil or sunflower oil for 6 weeks. However, the broilers receiving the tallow had a markedly increased plasma insulin response. Improved clearance of blood glucose in the birds receiving the diets rich in PUFA was ascribed to an increase in PUFA within the tissue membranes of these birds, resulting in an increased number of insulin receptor binding sites and an increased affinity of the insulin receptor for binding23.
A significantly increased glucose AUC was observed in the muscovy ducks in the HFD group compared to those in the STD group, after 4 weeks of feeding. This could be a sign of glucose intolerance as a result of the HFD consumption. However, the same effect was absent after 8 weeks of HFD feeding, indicating an adaptation to the HFD by the muscovy duck, the mechanisms of which should be further investigated. Given the HFD-induced changes in the insulin function observed in chickens, the lack of any observable changes in the glucose tolerance test parameters assessed in the present study may suggest some inter-species differences in the regulation of lipid metabolism amongst avian species.
Liver lipid content was similar between birds on the STD and HFD, for both bird species. Our results agree with those of Magubane et al.26, who observed no HFD-induced effects on liver lipid content in Japanese quail supplemented with 10% canola oil (rich in MUFA) for a period of 7 weeks. Tissue lipid deposition is the net result of the absorption of dietary fatty acids, the oxidation of fatty acids for energy and de novo fatty acid synthesis27. The fatty acid profile of the diet consumed impacts the resulting lipid deposition within various body tissues. Increased lipid deposition is generally observed following consumption of SFA, whereas PUFA tend to reduce lipid deposition27-29. Results from previous studies assessing liver lipid deposition using predominantly chickens, are not in agreement with those of the current study. The contradictory results may be due to inter-avian species differences in lipid metabolism. Genetic dissimilarities in lipid metabolism between different avian species and even between different breeds of the same avian species have been observed in studies involving the induction of liver lipid accumulation by overfeeding. Hermier et al.30 observed less hepatic lipid accumulation following overfeeding in common ducks compared to muscovy ducks, which was ascribed to more efficient routing of liver lipids towards secretion via VLDLs, thus promoting the storage of lipids in extrahepatic tissues30. An earlier study by Hermier et al.31 demonstrated increased susceptibility to hepatic steatosis in the landes geese compared to the rhine geese, which was accredited to the reduced ability of this breed to export VLDLs from the liver, thus favouring hepatic lipid accumulation31. Fournier et al.32 observed that compared to the landes goose, the Poland goose reacted to overfeeding by increasing exportation of all liver lipids, resulting in increased extrahepatic fattening as opposed to hepatic steatosis32. Since differences in hepatic lipid metabolism following overfeeding have been observed between different avian species/breeds, it is possible that essential differences exist in the handling of dietary lipids and their subsequent fate following HFD feeding as well. This could explain the discrepancies observed between the liver lipid deposition in the birds used in the present study and those reported on in previous studies.
Significantly increased serum cholesterol levels were observed in the guinea fowl and muscovy ducks following the HFD feeding in the current study. The hypercholesterolemia was well tolerated by the birds as they exhibited no signs of poor health, suggesting an effective coping mechanism in terms of managing potential modifications in lipid metabolism and lipid portioning, as a result of changes in dietary lipid content17. The intake of diets rich in SFA have been linked to the development of hypercholesterolemia33. Monfaredi et al.34 observed significantly increased serum cholesterol concentrations in broilers receiving beef tallow at either 20 or 40 g kg–1 compared to broilers on the control diet. Similarly, Celebi and Utlu35 observed a significant increase in serum cholesterol concentrations in laying hens supplemented with 4% beef tallow compared to those not receiving additional lipids. Ayala et al.10observed significantly increased serum cholesterol concentrations in chickens, following the consumption of a diet enriched with 20% cholesterol and 2% palm oil (rich in SFA). Increased serum cholesterol concentrations following consumption of SFA have been attributed to decreases in hepatic low-density lipoprotein receptor expression and activity and increases in the production of low-density lipoproteins36-39. In addition, certain saturated fatty acids such as palmitic acid, myristic acid and lauric acid are more hypercholesterolemic than others36, thus the fatty acid composition of the animal fats and vegetable oils added to poultry diets needs to be carefully considered, with regards to maintaining the overall health status of the birds during production.
CONCLUSION
The present study showed that feeding a HFD (rich in saturated fatty acids) to guinea fowl and muscovy duck for up to 8 weeks did not significantly affect any of the glucose tolerance parameters examined or the liver lipid content, despite the resulting hypercholesterolemia. Thus, in a poultry production setting, the diet would in fact be safe in terms of maintaining the overall health status of these alternative poultry species. The mechanisms involved in the metabolism of various lipid types (i.e. saturated vs. unsaturated) and subsequent handling of the lipid metabolites in these bird species needs to be further investigated in order to establish the physiological mechanisms behind the resistance of these alternative poultry species to the metabolic effects of a HFD.
SIGNIFICANCE STATEMENTS
The current study demonstrates that the use of a high-fat diet, rich in saturated fatty acids, in a poultry production setting would in fact be safe in terms of maintaining the overall health status of these alternative poultry species, since the diet was well-tolerated by both the guinea fowl and muscovy ducks, with no obvious adverse health effects observed. The study provides insight into the effects of high-fat diet consumption in these alternative poultry species which are becoming popular table birds, information which is important with regards to poultry production practices and the development of species-specific poultry diets.
ACKNOWLEDGMENTS
This study was supported by grants obtained from the Faculty Research Committee Funds of the University of the Witwatersrand (PHSL 013), the National Research Foundation (IFR2011040300007) and the Oppenheimer Memorial Trust Fund (OMT 19586/01).The authors would like to thank Margaret Badenhorst, Busisani Lembede, Mhlengi Magubane and Tanusha Dukhan from the School of Physiology, University of the Witwatersrand, South Africa, for their technical assistance.
REFERENCES
- Tierney, A.C., J. McMonagle, D.I. Shaw, H.L. Gulseth and O. Helal et al., 2011. Effects of dietary fat modification on insulin sensitivity and on other risk factors of the metabolic syndrome-LIPGENE: A European randomized dietary intervention study. Int. J. Obesity, 35: 800-809.
Direct Link - Roche, H.M., 2005. Fatty acids and the metabolic syndrome. Proc. Nutr. Soc., 64: 23-29.
CrossRefDirect Link - Moreki, J.C. and D. Seabo, 2012. Guinea fowl production in Botswana. J. World's Poult. Res., 2: 1-4.
Direct Link - Tjetjoo, S.U., J.C. Moreki, S.J. Nsoso and O.R. Madibela, 2013. Growth performance of Guinea fowl fed diets containing yellow maize, millet and white sorghum as energy sources and raised under intensive system. Pak. J. Nutr., 12: 306-312.
CrossRefDirect Link - Meng, X., B.A. Slominski and W. Guenter, 2004. The effect of fat type, carbohydrase and lipase addition on growth performance and nutrient utilization of young broilers fed wheat-based diets. Poult. Sci., 83: 1718-1727.
PubMedDirect Link - Sanz, M., A. Flores and C.J. Lopez-Bote, 2000. The metabolic use of energy from dietary fat in broilers is affected by fatty acid saturation. Br. Poult. Sci., 41: 61-68.
CrossRefPubMedDirect Link - Burlikowska, K., A. Piotrowska and R. Szymeczko, 2010. Effect of dietary fat type on performance, biochemical indices and fatty acids profile in the blood serum of broiler chickens. J. Anim. Feed Sci., 19: 440-451.
CrossRefDirect Link - Firman, J.D., A. Kamyab and H. Leigh, 2008. Comparison of fat sources in rations of broilers from hatch to market. Int. J. Poult. Sci., 7: 1152-1155.
CrossRefDirect Link - Ayala, I., A.M. Castillo, G. Adanez, A. Fernandez-Rufette, B.G. Perez and M.T. Castells, 2009. Hyperlipidemic chicken as a model of non-alcoholic steatohepatitis. Exp. Biol. Med., 234: 10-16.
CrossRefPubMedDirect Link - Noyan, A., W.J. Lossow, N. Brot and I.L. Chaikoff, 1964. Pathway and form of absorption of palmitic acid in the chicken. J. Lipid Res., 5: 538-541.
Direct Link - Leyton, J., P.J. Drury and M.A. Crawford, 1987. Differential oxidation of saturated and unsaturated fatty acids in vivo in the rat. Br. J. Nutr., 57: 383-393.
CrossRefPubMedDirect Link - Honors, M.A., S.L. Hargrave and K.P. Kinzig, 2012. Glucose tolerance in response to a high‐fat diet is improved by a high‐protein diet. Obesity, 20: 1859-1865.
CrossRefDirect Link - Figueiredo-Silva, A.C., S. Panserat, S. Kaushik, I. Geurden and S. Polakof, 2012. High levels of dietary fat impair glucose homeostasis in rainbow trout. J. Exp. Biol., 215: 169-178.
CrossRefDirect Link - Simon, J. and G. Rosselin, 1979. Effect of intermittent feeding on glucose-insulin relationship in the chicken. J. Nutr., 109: 631-641.
PubMedDirect Link - Donaldson, J., R. Dangarembizi, B. Mtetwa, M.T. Madziva and K.H. Erlwanger, 2014. The progressive effects of a high-fat diet on erythrocyte osmotic fragility, growth performance and serum triglyceride and cholesterol levels in Guinea fowl (Numida meleagris) and Muscovy duck (Cairina moschata). J. Anim. Physiol. Anim. Nutr., 98: 867-874.
CrossRefDirect Link - Weideman, M.A., E. Chivandi and K.H. Erlwanger, 2012. Glucose tolerance and lipid absorption in Guinea fowl (Numida meleagris) and domestic fowl (Gallus gallus var. domesticus). Asian J. Anim. Vet. Adv., 7: 653-663.
CrossRefDirect Link - Loxham, S.J.G., J. Teague, S.M. Poucher, J. de Schoolmester, A.V. Turnbull and F. Carey, 2007. Glucagon challenge in the rat: A robust method for the in vivo assessment of Glycogen phosphorlyase inhibitor efficacy. J. Pharm. Toxicol. Methods, 55: 71-77.
CrossRefPubMedDirect Link - Bligh, E.G. and W.J. Dyer, 1959. A rapid method of total lipid extraction and purification. Can. J. Biochem. Physiol., 37: 911-917.
CrossRefPubMedDirect Link - Lomax, M.A., G. Karamanlidis, J. Laws, S.G. Cremers, P.D. Weinberg and L. Clarke, 2013. Pigs fed saturated fat/cholesterol have a blunted hypothalamic-pituitary-adrenal function, are insulin resistant and have decreased expression of IRS-1, PGC1α and PPARα. J. Nutr. Biochem., 24: 656-663.
CrossRefDirect Link - Kaiyala, K.J., R.L. Prigeon, S.E. Kahn, S.C. Woods, D. Porte and M.W. Schwartz, 1999. Reduced β-cell function contributes to impaired glucose tolerance in dogs made obese by high-fat feeding. Am. J. Physiol.-Endocrinol. Metab., 277: E659-E667.
Direct Link - Newman, R.E., W.L. Bryden, A.C. Kirby, L.H. Storlien and J.A. Downing, 2005. Dietary n-3 and n-6 fatty acids alter avian glucose metabolism. Br. Poult. Sci., 46: 104-113.
CrossRefDirect Link - Crespo, N. and E. Esteve-Garcia, 2003. Polyunsaturated fatty acids reduce insulin and very low density lipoprotein levels in broiler chickens. Poult. Sci., 82: 1134-1139.
CrossRefDirect Link - Scanes, C.G. and E. Braun, 2013. Avian metabolism: Its control and evolution. Front. Biol., 8: 134-159.
CrossRefDirect Link - Magubane, M.M., B.W. Lembede, K.H. Erlwanger, E. Chivandi and J. Donaldson, 2013. Fat absorption and deposition in Japanese quail (Coturnix coturnix japonica) fed a high fat diet. J. S. Afr. Vet. Assoc., Vol. 84.
CrossRef - Smink, W., W.J.J. Gerrits, R. Hovenier, M.J.H. Geelen, M.W.A. Verstegen and A.C. Beynen, 2010. Effect of dietary fat sources on fatty acid deposition and lipid metabolism in broiler chickens. Poult. Sci., 11: 2432-2440.
CrossRefPubMedDirect Link - Crespo, N. and E. Esteve-Garcia, 2002. Nutrient and fatty acid deposition in broilers fed different dietary fatty acid profiles. Poult. Sci., 81: 1533-1542.
CrossRefDirect Link - Velasco, S., L.T. Ortiz, C. Alzueta, A. Rebole, J. Trevino and M.L. Rodriguez, 2010. Effect of inulin supplementation and dietary fat source on performance, blood serum metabolites, liver lipids, abdominal fat deposition and tissue fatty acid composition in broiler chickens. Poult. Sci., 89: 1651-1662.
CrossRefPubMedDirect Link - Hermier, D., G. Guy, S. Guillaumin, S. Davail, J.M. Andre and R. Hoo-Paris, 2003. Differential channelling of liver lipids in relation to susceptibility to hepatic steatosis in two species of ducks. Comp. Biochem. Physiol. Part B: Biochem. Mol. Biol., 135: 663-675.
CrossRefDirect Link - Hermier, D., A. Saadoun, M.R. Salichon, N. Sellier and D. Rousselot-Pailley, 1991. Plasma lipoproteins and liver lipids in two breeds of geese with different susceptibility to hepatic steatosis: Changes induced by development and force-feeding. Lipids, 26: 331-339.
PubMedDirect Link - Fournier, E., R. Peresson, G. Guy and D. Hermier, 1997. Relationships between storage and secretion of hepatic lipids in two breeds of geese with different susceptibility to liver steatosis. Poult. Sci., 76: 599-607.
PubMedDirect Link - Zulet, M.A., A. Barber, H. Garcin, P. Higueret and J.A. Martinez, 1999. Alterations in carbohydrate and lipid metabolism induced by a diet rich in coconut oil and cholesterol in a rat model. J. Am. Coll. Nutr., 18: 36-42.
CrossRefPubMedDirect Link - Monfaredi, A., M. Rezaei and H. Sayyahzadeh, 2011. Effect of supplemental fat in low energy diets on some blood parameters and carcass characteristics of broiler chicks. S. Afr. J. Anim. Sci., 41: 24-32.
Direct Link - Celebi, S. and N. Utlu, 2006. Influence of animal and vegetable oil in layer diets on performance and serum lipid profile. Int. J. Poult. Sci., 5: 370-373.
CrossRefDirect Link - Fernandez, M.L. and K.L. West, 2005. Mechanisms by which dietary fatty acids modulate plasma lipids 1. J. Nutr., 135: 2075-2078.
Direct Link - Mustad, V.A., J.L. Ellsworth, A.D. Cooper, P.M. Kris-Etherton and T.D. Etherton, 1996. Dietary linoleic acid increases and palmitic acid decreases hepatic LDL receptor protein and mRNA abundance in young pigs. J. Lipid Res., 37: 2310-2323.
PubMedDirect Link - Tripodi, A., P. Loria, M.A. Dilengite and N. Carulli, 1991. Effect of fish oil and coconut oil diet on the LDL receptor activity of rat liver plasma membranes. Biochim. Biophys. Acta (BBA)-Lipids Lipid Metabol., 1083: 298-304.
CrossRefDirect Link - Kuo, P., M. Weinfeld and J. Loscalzo, 1990. Effect of membrane fatty acyl composition on LDL metabolism in Hep G2 hepatocytes. Biochemistry, 29: 6626-6632.
CrossRefDirect Link