
Mitchell Wendt
Veterinary Medical Center and College of Veterinary Medicine, Chungbuk National University, 28644 Cheongju, Korea
Benthotage Chamara Jayasankha De Silva
Veterinary Medical Center and College of Veterinary Medicine, Chungbuk National University, 28644 Cheongju, Korea
Gang-Joon Heo
Veterinary Medical Center and College of Veterinary Medicine, Chungbuk National University, 28644 Cheongju, Korea
LiveDNA: 82.16116
Asian Journal of Animal and Veterinary Advances
Year: 2017 | Volume: 12 | Issue: 4 | Page No.: 205-211







ABSTRACT
Background and Objectives: A gram-negative zoonotic bacterium, Pseudomonas aeruginosa, is well-known for its antimicrobial resistance and ubiquity in nature. The current study aimed to examine pet turtle-borne P. aeruginosa for its virulence determinants and the antimicrobial resistance. Methodology: Twenty-four turtles purchased from pet shops and online markets in Korea were examined to determine whether they excreted P. aeruginosa. Presumptive P. aeruginosa was isolated from the fecal samples of pet turtles by selective media incubation and verified with biochemical testing. Seventeen isolates were genetically characterized by 16S rRNA sequencing and confirmed as P. aeruginosa. These strains were further tested for antimicrobial resistance by disk diffusion test and PCR assays were conducted to detect virulence genes. Results: All tested strains showed susceptibility to ciprofloxacin and ofloxacin but were completely resistant to amoxicillin, colistin, streptomycin, cephalothin, trimethoprim-sulfamethoxazole, chloramphenicol, imipenem, cefoxitin and nalidixic acid. Against gentamycin, tetracycline, ceftriaxone and amikacin, some strains showed complete resistance while some were intermediate resistant. The PCR assay detected the presence of virulence genes, toxA (100%), lasB (100%) and exoS (53%), which aid in pathogenicity against humans. Conclusion: All the results indicated that the pet turtles pose a potential public health risk due to prospective zoonotic P. aeruginosa infection.
PDF Abstract XML References Citation
Received: January 09, 2017;
Accepted: March 01, 2017;
Published: March 15, 2017
How to cite this article
Mitchell Wendt, Benthotage Chamara Jayasankha De Silva and Gang-Joon Heo, 2017. Virulence Factors and Antimicrobial Resistance of Pseudomonas aeruginosa Isolated from Pet Turtles. Asian Journal of Animal and Veterinary Advances, 12: 205-211.
DOI: 10.3923/ajava.2017.205.211
URL: https://scialert.net/abstract/?doi=ajava.2017.205.211
DOI: 10.3923/ajava.2017.205.211
URL: https://scialert.net/abstract/?doi=ajava.2017.205.211
INTRODUCTION
Pseudomonas aeruginosa is a gram-negative bacterium well-known for its multiple antimicrobial resistances and ubiquity1. It is an opportunistic pathogen that primarily infects cystic fibrosis patients, burn victims and immunocompromised patients such as young children, the elders, or the chronically ill2. Pseudomonas aeruginosa infections can involve any part of the body and cause a range of symptoms from minor gastrointestinal infections resulting in diarrhea, fever and headache to severe infections including meningitis, bacterial keratitis, endophthalmitis and necrotizing enterocolitis3. Moreover, P. aeruginosa is a leading cause of nosocomial infections which is well-characterized as a source of respiratory tract infections that contribute to morbidity and mortality among cystic fibrosis patients4.
Genetically, P. aeruginosa virulence is believed to be multifactorial in that several factors are required for overall virulence and combinatorial in that virulence can result from many combinations of virulence factors5. Thus, environmental stimuli, horizontal gene transfer and variations within pathogenic gene islands can alter the virulence of a given strain of P. aeruginosa. Moreover, the prevalence of specific P. aeruginosa virulence genes is not itself indicative of pathogenicity6. However, certain generally conserved features of the P. aeruginosa genome aid in virulence. These include the ability to secrete exotoxins, elastase and type III effector proteins. Genes encoding these factors include toxA, lasB and exoS, respectively7-9.
While P. aeruginosa is ubiquitous in inanimate environments such as soil and water, it is also commonly found in the oral cavity and intestinal tracts of reptiles 10,11. As an opportunistic pathogen, it can cause ulcerative stomatitis, pneumonia, dermatitis and septicemia in reptiles12. Digestive tract inhabitation of P. aeruginosa there by poses a threat to immunocompromised reptiles and causes reptiles to constitute a potential source of zoonotic disease. In addition, a reptile’s fecal matter containing P. aeruginosa can contaminate its environment and the adaptability of P. aeruginosa allows it to readily thrive in diverse environments, especially those are not cleaned thoroughly13,14.
Globalization and development of modern infrastructures have made importation of exotic pets such as insects, tropical fish and especially turtles easier and increasingly popular15. However, the popularity of exotic pets has not been accompanied by dissemination of the public health concerns that arise when raising them16. Common expressions of affection towards pets involve physical contact, yet turtle owners may not be aware of the risk of contracting a pathogen when handling their pet without adequate countermeasures17. Therefore, examining the turtle-borne pathogenic bacteria is vital to prevent potential infections.
The aim of this stuyd was to identify and characterize pathogenic P. aeruginosa isolated from pet turtles by detecting the susceptibility pattern of antibiotics and PCR screening of virulence factors in order to evaluate the potential risk of exposure to the zoonotic pathogen.
MATERIALS AND METHODS
Sample collection: Twenty-four turtles belonging to seven commercially popular species were purchased randomly through pet shops and online markets in Korea. The purchased turtles (n = 24) were of varying age, size and weight including 3 Chinese stripe-necked turtles (Ocadia sinensis), 5 yellow-bellied sliders (Trachemys scripta scripta), 6 river cooters (Pseudemys concinna concinna), 1 Northern Chinese softshell turtle (Pelodiscus maackii), 3 Western painted turtles (Chrysemys picta belli), 3 peninsula cooters (Pseudemys peninsularis) and 3 common musk turtles (Sternotherus odoratus). All turtles were clinically examined upon purchase and noted to be apparently and clinically healthy and without signs of infection.
Raising condition of pet turtles: Seven cages each containing turtles of the same species were managed; each cage contained a slope made from soil and pebbles, 2 L of sterilized water and a canister filter to maintain water quality. The turtles were raised following the general husbandry method18. Gammarus dried shrimp with calcium supplements (Samhotech Co., Ltd., Seoul, Korea) were fed twice a day while water temperature was kept within 26±2°C, pH 6.5-8.2, with 12 h of photoperiod per day.
Isolation and identification of P. aeruginosa: Each of the purchased turtles was placed in a separate 500 mL beaker with 5 mL of sterilized distilled water for 24 h. About 1 mL of the distilled water containing a turtle’s feces was taken as a fecal sample. In order to prevent bacterial transmission between individuals, poly-gloves were changed each time an animal was handled. Fecal samples were enriched in alkaline peptone water (pH 8.6±0.2) at 37°C for 24 h.
| Table 1: | Primers used for PCR amplification of virulence genes |
 | |
The enriched samples were streaked onto starch-ampicillin agar and incubated at 37°C for 24 h.
Pseudomonas aeruginosa was presumptively differentiated from potential A. hydrophila using starch hydrolysis, indole and glucose oxidation and fermentation tests. Subcultures were incubated at 42°C for 24 h to differentiate P. aeruginosa from other Pseudomonas spp. Further phenotypic verification was performed using gram staining, motility, citrate utilization and Kovac’s oxidase tests, all of which matched typical reactions for P. aeruginosa.
Genomic DNA was extracted from presumptively identified P. aeruginosa isolates by Chelex-100 extraction method and PCR for 16S rRNA was performed using universal primers 12F and 1492R. Amplicons were sequenced and tested for the similarity using BLAST algorithm of NCBI database to confirm the species status.
Detection of virulence genes by PCR: The presence of toxA, lasB elastase and exoS genes among seventeen identified P. aeruginosa strains was detected by PCR using the primers proposed by Lanotte et al.19 (Table 1). Each PCR mixture was 20 μL total volume comprising of 10 μL of Quick Taq® HS DyeMix (TOYOBO, Japan) 1 μL of 10 pmol μL–1 each primer and 1 μL of template. Each gene was amplified separately in a thermal cycler using the following protocol: 94°C for 3 min, 30 cycles of 94°C for 30 sec, 55°C for 1 min and 72°C for 1 min 30 sec and 72°C for 5 min. The PCR products were checked in 1% agarose gel at 100 V using gel loading buffer with DNA stain (Jena Bioscience, Germany).
Antimicrobial susceptibility testing: Pseudomonas aeruginosa strains were subjected to disk diffusion test with 15 common antimicrobials which are used in veterinary medicine against gram-negative bacteria. Pseudomonas aeruginosa was cultured on Tryptic Soy Agar (TSA) and generated colonies were adjusted to a turbidity of 0.5 McFarland units (1.5×106 CFU mL–1) with saline. The bacterial suspension was then spread on Mueller Hinton Agar (MHA). Disks containing amikacin (30 μg), amoxicillin (30 μg), cefoxitin (30 μg), ceftriaxone (30 μg), cephalothin (30 μg), chloramphenicol (30 μg), ciprofloxacin (5 μg), colistin sulfate (10 μg), gentamycin (10 μg), imipenem (10 μg), nalidixic acid (30 μg), ofloxacin (5 μg), streptomycin (10 μg), tetracycline (30 μg) and trimethoprim-sulfamethoxazole (25 μg) purchased from Kisan Biotech Co., Ltd. (Seoul, Korea) were placed on inoculated MHA plates and incubated for 18 h at 37°C. After incubation, antimicrobial activity was determined by measuring the diameter of the clear zones formed on the MHA plates. Clinical and Laboratory Standards Institute (CLSI) standards were used to determine the efficacy of each antimicrobial20.
RESULTS
Isolation and identification of P. aeruginosa: Suspicious colonies were isolated from 17 of 24 turtle fecal samples and no growth was noted on the other plates (n = 7). Positive samples were from 2/3 (67%) Chinese stripe-necked turtles, 4/5 (80%) yellow-bellied sliders, 4/6 (67%) river cooters, 1/1 (100%) Chinese softshell turtle, 3/3 (100%) Western painted turtles, 2/3 (67%) peninsula cooters and 1/3 (33%) common musk turtles. The colonies emitted a distinctive "grape-like" odor which is this characteristic of P. aeruginosa21. Furthermore, biochemical testing and incubation at 42°C presumptively differentiated the colonies from A. hydrophila and other Pseudomonas spp. and indicated P. aeruginosa. In addition, 16S rRNA sequencing results confirmed the isolates as P. aeruginosa.
Detection of virulence genes by PCR: As shown in Fig. 1, PCR results clearly indicated the presence of each virulence gene among studied strains. All tested isolates (100%) were positive for toxA and lasB while 9 out of 17 strains (53%) were positive for exoS gene.
Antimicrobial resistance testing: The disk diffusion test results are shown in Table 2. All strains completely resisted amoxicillin, colistin, streptomycin, cephalothin, trimethoprim-sulfamethoxazole, chloramphenicol, imipenem, cefoxitin and nalidixic acid. In partial, 82, 59, 65 and 59% of the isolates were resistant against gentamycin, ceftriaxone, tetracycline and amikacin, respectively. Intermediate resistance was detected for gentamycin, tetracycline, ceftriaxone and amikacin among 6, 35, 41 and 35% of the strains, respectively. However, all tested isolates were susceptible for tested fluoroquinolones; ciprofloxacin and ofloxacin.
 | |
| Fig. 1(a-c): | PCR results of (a) toxA, (b) lasB and (c) exoS virulence genes in P. aeruginosa strains (lanes 3-19) |
| Lane 1: Negative control, Lane 2: Positive control; ATCC reference strain and Lane 20: 100 bp DNA ladder | |
DISCUSSION
In this study, P. aeruginosa was isolated from fecal samples of 17 (71%) turtles out of 24 turtles and were genetically confirmed it as P. aeruginosa. The high percentage of fecal samples containing P. aeruginosa could be attributed to environmental contamination from fecal samples of one turtle leading to horizontal transmission to others, which suggested that careless management of pet shops and inadequate cleaning of turtle raising environments could increase P. aeruginosa contamination. This result indicated that raising and distributing pet turtles carries an associated risk of P. aeruginosa contamination.
PCR assays found that the isolates contained the virulence genes toxA, lasB and exoS. These genes, while not inherently indicative of pathogenicity, encode pathogenic factors that aid in cytotoxicity7–9. Moreover, the presence of both lasB and exoS increases P. aeruginosa infiltration of epithelial cells due to a regulatory effect of lasB on exoS22. The absence of a consistent virulence gene profile in P. aeruginosa infections displays the bacteria’s ability to adapt and infiltrate humans with its conserved core genome6. Therefore, the isolated strains of P. aeruginosa were potentially pathogenic or could become pathogenic given the requisite environmental factors to induce activation of necessary combinations of genes.
As exotic pet ownership becomes increasingly popular, the need for awareness about associated public health concerns grows. Reptiles are known as a potential reservoir for zoonotic pathogens and they often excrete bacteria as normal flora12. They are therefore considered a high-risk pet group23. Pseudomonas aeruginosa has previously been isolated from healthy wild tortoises and sea turtles24, 25. However, this is the first study examining virulence factors and antimicrobial resistance simultaneously focusing on turtles sold in pet shops. Pet turtles are known as a source of salmonellosis and this motivated the 1975 FDA ban on the sale of turtles less than 10.2 cm (4 inches) of carapace size in the US23. Nevertheless, these regulations do not exist in other first-world countries such as Korea. Furthermore, the environmental adaptability and mechanisms of antimicrobial resistance exhibited by P. aeruginosa necessitate prevention of infections to avoid stimulating further evolution of antimicrobial resistance1,26. Pet shops must inform their customers about infection risks and emphasize how to maintain environmental cleanliness. Moreover, health care providers should educate patients and the public about the risks of pet turtles and proper sanitation techniques.
As an opportunistic pathogen with a high degree of adaptability, P. aeruginosa commonly causes nosocomial infections, especially among patients with cystic fibrosis or compromised epithelia and those requiring ventilators27-29. Pseudomonas aeruginosa nosocomial outbreaks have stemmed from contaminated tap water, hand lotion and children’s bath toys, so preventing the introduction of P. aeruginosa into hospitals is critical13,14,30. Therefore, even healthy individuals who own pet turtles should take sanitary precautions before visiting hospital patients.
Antimicrobial resistance testing of the strains revealed resistance not only to many common antimicrobials that were frequently resisted by P. aeruginosa but also to amikacin and gentamicin antimicrobials usually effective against P. aeruginosa31.
| Table 2: | Disk diffusion test results of P. aeruginosa isolated from pet turtles |
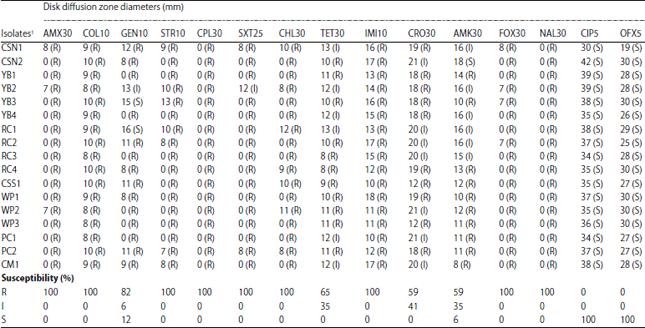 | |
AMX30: Amoxicillin (30 μg), COL10: Colistin sulfate (10 μg), GEN10: Gentamycin (10 μg), STR10: Streptomycin (10 μg), CPL30: Cephalothin (30 μg), SXT25: Trimethoprim-sulfamethoxazole (25 μg), CHL30: Chloramphenicol (30 μg), TET30: Tetracycline (30 μg), IMI10: Imipenem (10 μg), CRO30: Ceftriaxone (30 μg), AMK30: Amikacin (30 μg), FOX30: Cefoxitin (30 μg), NAL30: Nalidixic acid (30 μg), CIP5: Ciprofloxacin (5 μg) and OFX5: Ofloxacin (5 μg), S: Susceptible, I: Intermediate, R: Resistant, 1Isolate number is given according to the turtle species from which the strains were isolated; CSN: Chinese stripe-necked turtle, YB: Yellow-bellied slider, RC: River cooter, CSS: Chinese softshell turtle, WP: Western painted turtle, PC: Peninsula cooter, CM: Common musk turtle | |
Besides, all the strains were completely resistant to colistin, which was previously noted as an effective treatment for P. aeruginosa that developed multidrug-resistance32. Although, all the strains were still susceptible to ciprofloxacin and ofloxacin, possession of non-intrinsic resistance to amikacin and gentamicin among majority showed its ability to adapt and evolve antimicrobial resistance. On the other hand, it indicated the potential use of fluoroquinolones for the treatment of P. aeruginosa as previously reported33. However, public awareness of the risk of contracting or transferring P. aeruginosa from pet turtles is integral to prevent further evolution of antimicrobial resistance.
CONCLUSION
It was concluded that the pet turtles are vessels for multidrug resistant P. aeruginosa which are harboring virulence determinants; toxA, lasB and exoS. Hence, those bacteria contracted from contact with pet turtles are potentially pathogenic posing a significant public health risk.
SIGNIFICANCE STATEMENT
This is the first study to characterize simultaneously the virulence determinants and antimicrobial resistance of P. aeruginosa isolated from several species of pet turtles. The outcomes of this study revealed that studied strains of P. aeruginosa were multidrug resistant, possess virulence factors thereby, pose a potential public health risk.
ACKNOWLEDGMENT
The author would like to thank Basic Science Research Program who supported this study throught the National Research Foundation of Korea. We also thank to the Ministry of Education (NRF-2015R1D1A1A01060638) for their funding.
REFERENCES
- Livermore, D.M., 2002. Multiple mechanisms of antimicrobial resistance in Pseudomonas aeruginosa: Our worst nightmare? Clin. Infect. Dis., 34: 634-640.
CrossRefDirect Link - Vonberg, R.P. and P. Gastmeier, 2005. Isolation of infectious cystic fibrosis patients: Results of a systematic review. Infect. Control Hosp. Epidemiol., 4: 401-409.
CrossRefDirect Link - Lee, D.G., J.M. Urbach, G. Wu, N.T. Liberati and R.L. Feinbaum et al., 2006. Genomic analysis reveals that Pseudomonas aeruginosa virulence is combinatorial. Genome Biol., Vol. 7.
CrossRefDirect Link - Bradbury, R.S., L.F. Roddam, A. Merritt, D.W. Reid and A.C. Champion, 2010. Virulence gene distribution in clinical, nosocomial and environmental isolates of Pseudomonas aeruginosa. J. Med. Microbiol., 59: 881-890.
CrossRefPubMedDirect Link - Iglewski, B.H. and D. Kabat, 1975. NAD-dependent inhibition of protein synthesis by Pseudomonas aeruginosa toxin. Proc. Nat. Acad. Sci., 72: 2284-2288.
Direct Link - Rust, L., E.C. Pesci and B.H. Iglewski, 1996. Analysis of the Pseudomonas aeruginosa elastase (lasB) regulatory region. J. Bacteriol., 178: 1134-1140.
CrossRefDirect Link - Van Delden, C. and B.H. Iglewski, 1998. Cell-to-cell signaling and Pseudomonas aeruginosa infections. Emerg. Infect. Dis., 4: 451-460.
Direct Link - Warwick, C., P.C. Arena, C. Steedman and M. Jessop, 2012. A review of captive exotic animal-linked zoonoses. J. Environ. Health Res., 12: 9-24.
Direct Link - Becks, V.E. and N.M. Lorenzoni, 1995. Pseudomonas aeruginosa outbreak in a neonatal intensive care unit: A possible link to contaminated hand lotion. Am. J. Infect. Control, 23: 396-398.
CrossRefDirect Link - Bert, F., E. Maubec, B. Bruneau, P. Berry and N. Lambert-Zechovsky, 1998. Multi-resistant Pseudomonas aeruginosa outbreak associated with contaminated tap water in a neurosurgery intensive care unit. J. Hosp. Infect., 39: 53-62.
CrossRefDirect Link - Di Ianni, F., P.L. Dodi, C.S. Cabassi, I. Pelizzone and A. Sala et al., 2015. Conjunctival flora of clinically normal and diseased turtles and tortoises. BMC Vet. Res., Vol. 11.
CrossRef - Lanotte, P., S. Watt, L. Mereghetti, N. Dartiguelongue, A. Rastegar-Lari, A. Goudeau and R. Quentin, 2004. Genetic features of Pseudomonas aeruginosa isolates from cystic fibrosis patients compared with those of isolates from other origins. J. Med. Microbiol., 53: 73-81.
CrossRefDirect Link - Cox, C.D. and J. Parker, 1979. Use of 2-aminoacetophenone production in identification of Pseudomonas aeruginosa. J. Clin. Microbiol., 9: 479-484.
Direct Link - Cowell, B.A., S.S. Twining, J.A. Hobden, M.S.F. Kwong and S.M.J. Fleiszig, 2003. Mutation of lasA and lasB reduces Pseudomonas aeruginosa invasion of epithelial cells. Microbiology, 149: 2291-2299.
CrossRefDirect Link - Warwick, C., P.C. Arena and C. Steedman, 2013. Health implications associated with exposure to farmed and wild sea turtles. JRSM Short Rep., Vol. 4.
CrossRefDirect Link - Dickinson, V.M., T. Duck, C.R. Schwalbe, J.L. Jarchow and M.H. Trueblood, 2001. Nasal and cloacal bacteria in free-ranging desert tortoises from the Western United States. J. Wildlife Dis., 37: 252-257.
CrossRefDirect Link - Govan, J.R. and V. Deretic, 1996. Microbial pathogenesis in cystic fibrosis: Mucoid Pseudomonas aeruginosa and Burkholderia cepacia. Microbiol. Rev., 60: 539-574.
Direct Link - Douglas, M.W., K. Mulholland, V. Denyer and T. Gottlieb, 2001. Multi-drug resistant Pseudomonas aeruginosa outbreak in a burns unit-An infection control study. Burns, 72: 131-135.
CrossRefDirect Link - Giantsou, E. and K. Manolas, 2011. Superinfections in Pseudomonas aeruginosa ventilator-associated pneumonia. Minerva Anestesiol., 77: 964-970.
Direct Link - Buttery, J.P., S.J. Alabaster, R.G. Heine, S.M. Scott, R.A. Crutchfield and S.M. Garland, 1998. Multiresistant Pseudomonas aeruginosa outbreak in a pediatric oncology ward related to bath toys. Pediatric Infect. Dis. J., 17: 509-513.
Direct Link - Poole, K., 2005. Aminoglycoside resistance in Pseudomonas aeruginosa. Antimicrob. Agents Chemother., 49: 479-487.
CrossRefDirect Link - Sabuda, D.M., K. Laupland, J. Pitout, B. Dalton, H. Rabin, T. Louie and J. Conly, 2008. Utilization of colistin for treatment of multidrug-resistant Pseudomonas aeruginosa. Can. J. Infect. Dis. Med. Microbiol., 19: 413-418.
CrossRefDirect Link - Meletis, G. and M. Bagkeri, 2013. Pseudomonas aeruginosa: Multi-drug-resistance development and treatment options. In: Infection Control, Basak, S. (Ed.), InTech Open Publisher, Rijeka, Croatia, ISBN: 978-953-51-1145-0, pp: 33-56.
CrossRefDirect Link