
Sidharth Prasad Mishra
Department of Animal Breeding and Genetics, College of Veterinary Science and Animal Husbandry, Orissa University of Agriculture and Technology, Bhubaneswar, Odisha, India
Gangadhar Nayak
Department of Animal Breeding and Genetics, College of Veterinary Science and Animal Husbandry, Orissa University of Agriculture and Technology, Bhubaneswar, Odisha, India
Chinmoy Mishra
Department of Animal Breeding and Genetics, College of Veterinary Science and Animal Husbandry, Orissa University of Agriculture and Technology, Bhubaneswar, Odisha, India
Asian Journal of Animal and Veterinary Advances
Year: 2016 | Volume: 11 | Issue: 7 | Page No.: 423-428







ABSTRACT
Objective: A study was undertaken on the captive tiger population of Nandankanan Zoological Park, Odisha to identify the deleterious effect of inbreeding on different litter size, mortality, survivability and death due to different disease condition and other fitness traits. Methodology: The path of pedigree and inbreeding coefficient for 342 tigers were calculated from the available information of tiger national studbook. The correlation values between inbreeding coefficient and different fitness traits were estimated by using SAS software. Results: The litter size of tiger, longevity and inbreeding coefficient were found to be significantly (p<0.05) and positively correlated. However, age at death, survivability of cubs upto weaning and total numbers of cubs born throughout lifetime were significantly (p<0.05) and negatively correlated with inbreeding coefficient. The death due to a disease conditions like stress, accidental injury, nephritis and senility were significantly (p<0.05) associated with inbreeding of a tiger. The inbreeding had also an associationship with white colour of tigers. Conclusion: So, temptation to produce white tiger should be minimise in order to reduce inbreeding depression and proper care and management must have practiced for a longevity life of a inbred animal in captive condition as they will face extinction in wild environment.
PDF Abstract XML References Citation
How to cite this article
Sidharth Prasad Mishra, Gangadhar Nayak and Chinmoy Mishra, 2016. Effect of Inbreeding on Different Fitness Traits of Captive Tiger Population. Asian Journal of Animal and Veterinary Advances, 11: 423-428.
DOI: 10.3923/ajava.2016.423.428
URL: https://scialert.net/abstract/?doi=ajava.2016.423.428
DOI: 10.3923/ajava.2016.423.428
URL: https://scialert.net/abstract/?doi=ajava.2016.423.428
INTRODUCTION
Tiger is one of the species on the verge of extinction and its existence is threatened in its land of survival. In a certain period of time a species can get extinct if sufficient genetic diversity is not found in its population. In order to increase its number, breeding of tigers inside Zoo is inevitable. Habitat loss and invasive species are primary cause of population extinction. The genetic variability is an important criterion to sustain the danger of extinction1. But for survival of tigers, small populations inside Zoo are bred among themselves resulting inbreeding. There is growing incidence that the small population is more prone to inbreeding depression2,3. The population breeds with its own relatives. The genetic diversity or variability is reduced with increase in inbreeding. Again loss of genetic variation due to inbreeding increases homozygosity at chromosome level. Most of the deleterious traits are expressed in homozygous form in a population. Thus, inbreeding results in inbreeding depression which leads to decline in the phenotypic value of a trait4.
The fitness traits are affected most adversely due to inbreeding. Some of the important fitness traits of the tiger are number of young surviving upto the age of weaning, age at first parturition, age at mating, litter size, gestation period, sex ratio, inter parturition period etc. However, some metric traits (e.g., birth weight, disease conditions, life span etc.) are indirectly associated with fitness and are therefore, affected by increased level of inbreeding. Close inbreeding leads to reduction in fitness3,5,6. Moreover, genetic drift is another consequence of close relative mating5. An interesting finding with brother-sister, father-daughter and mother-son mating was birth of white tigers. But deformities and deficiencies begin to surface very soon in white tiger population e.g., cub mortality is high in tiger population7,8. Therefore, the present study was conducted in white tiger population (live and dead) with the objectives of (1) To estimate the inbreeding coefficient of each tiger, (2) To find the correlation values between inbreeding coefficient and fitness traits, (3) To find the dependency of inbreeding with diseases, colour and death of tigers and (4) To guide the authorities about the future breeding strategy of tiger to avoid inbreeding.
MATERIALS AND METHODS
Detail pedigree information of 342 tigers was collected from stud book of Nadankanan Zoologolical Park, Bhubaneswar, Odisha. The information on sire, dam, date of birth, date of death, sex and colour were collected for the period between 1964-2011. The path of pedigree for each tiger was determined and inbreeding coefficient (F) of each tiger was calculated by Falconer and Mackay9. The following traits were measured on each tiger: Birth weight, age at first parturition (females only), age at first mating, parity, total number of cubs born in life time, number of cubs live upto weaning (weaning age of cubs is 1 year), age at death, litter size, number of white cubs born, number of normal colour cubs born, gestation period, sex ratio of cubs, average inter parturition period, causes of death (death due to rejection by mother, cardiac failure, shock, accidental injury, still birth, chronic nephritis, septicemic disease condition, inanition pneumonia, hepatitis, respiratory failure, trypanosomiasis, debility, senility, endometritis, peritonitis, haemorragic gastroenteritis, anemia, tumor, paralysis, dyspnoea, jaundice and parasitic disease) were collected from post mortem registers.
Correlation between inbreeding coefficient and birth weight, age at first parturition, age at first mating, parity, number of cubs born in life time, number of cubs live upto weaning, age at death, litter size, number of white cubs born in life time, gestation period, sex ratio of cubs, average inter-parturition period were calculated by Becker10. The test of significance at 5% level was estimated for the traits that were affected with inbreeding by using SAS Software.
The animals were divided into seven groups according to range of F values (0-0.05, 0.05-0.10, 0.10-0.15, 0.15-0.20, 0.20-0.25, 0.25-0.30 and 0.30-0.35). Chi-square test of heterogeneity by Snedecor and Cochran11 between inbreeding coefficient groups and 24 different said diseases was conducted to find the effect of inbreeding on such disease conditions by using SAS Software. Further, the same test was applied to know the dependency of death on colour, sex and inbreeding coefficients as well as to find out the association of white colour and inbreeding coefficients.
RESULTS AND DISCUSSION
Inbreeding is defined as the mating of a pair of animals who are related to one or more common ancestors. The animal more closely related to each other shows greater degree of inbreeding and maximum loss of hybrid vigor. Scientific study has revealed that there is a direct link between inbreeding depression and loss of genetic variation and population viability12-14 illustrated that the probability of inbreeding in New Zealand species is expected to be much higher than that of other continental species but they are less affected to inbreeding depression as compared to elsewhere specie15. A population with a history of inbreeding is likely to exhibit less inbreeding depression as purged most of the deleterious allele as compared to an outbred population with a more recent and brief history of inbreeding supported to the findings of Templeton and Read16. Frankham et al.17 argued that inbred population can reach the same population size as non-inbred population or may be more susceptible to new disease condition or parasites and be slower to recover from any human setback or disaster.
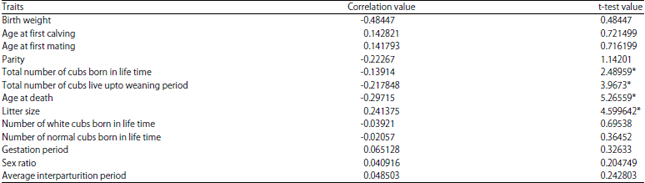
Correlation values of different fitness traits with inbreeding coefficients were calculated (Table 1).
Effect of inbreeding on litter size and litter survivability: Litter size and age at death were significantly (p<0.05) correlated with inbreeding. This is in agreement with previous finding of Wright4 who explained that inbreeding affected various components of fitness in animal. Dwyer18 reported that dams with high inbreeding coefficient developed a good maternal behaviour, which resulted in increased in survival rate of the litter but reproductive success is reduced. But, litter size was positively correlated whereas; age at death was negatively correlated with inbreeding coefficient. It satisfy the study of Dwyer18 who speculated that maternal behaviour of the dam increased because of the enhanced progesterone levels which favours the survival of the offsprings. But Boakes et al.19 illustrated that maternal inbreeding of 119 Zoo populations had a negative effect on fitness. Litter size increases with inbreeding coefficient value, but age at death decreases with the inbreeding coefficient value. This might be due to the fact that litter size is governed by additive effect of genes, but age at death is influenced by both heredity and environment. Quilicot20 illustrated that increase in dams total inbreeding coefficient is significant with mortality at days 7 (p<0.05), 30 (p<0.05) and 90 (0.10) indicating that it would decreases the mortality risk of the litter.
Effect of inbreeding on weaning age, calving interval, age at first calving and number of cubs born in the lifetime: The correlation of survivability of cubs upto weaning and total number of cubs born in the lifetime with inbreeding coefficient was negative but significant (p<0.05). Quilicot20 demonstrated that total inbreeding of an individual had a negative significant effect on mortality at weaning is an indication that inbreeding of the individual is the one influencing its survival. The similar trend was also reported earlier by Ralls et al.21 and Cassell et al.22. Inbred animals cannot survive in a harsh environment, but the survival is not affected if, the environment is conducive21. McParland et al.23 reported that increase in new inbreeding were much more associated than the ancestral coefficient with increase in calving interval (p<0.05) and age at 1st calving (p<0.001). But Ballou24 and Hedrick and Kalinowski2 showed that ancestral inbreeding had an unfavorable effect on calving interval, age at 1st calving and survival.
Effect of inbreeding on longevity of captive tiger population: The overall longevity of tiger population was also found to be significantly reduced by increase in inbreeding coefficient. Cassinello25 studied that three species of gazelle suffered from reduced longevity as affected by inbreeding depression. But Dorcas gazelle's was only affected by inbreeding depression while the other two species suffered a lower juvenile survival rate. Supported by Van Coillie et al.26, study on captive breeding bonobo (Panthera paniscus) population that infant mortality was higher in inbred offspring than non-inbred ones. But in contrast, Li27 assumed that not only inbreeding, but external factors like management and husbandry practice can have significant effect on survivability of the captive population.
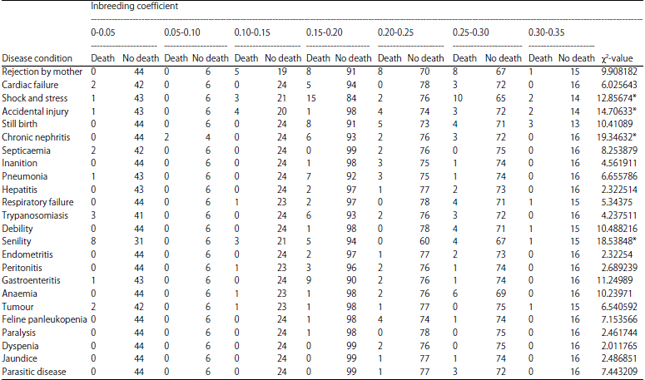
Effect of inbreeding on death due to different disease condition: The tigers under different inbreeding range face the problem of death due to 24 different diseases were identified (Table 2). There were 24 disease conditions.
| Table 1: | Correlation values of different traits with inbreeding coefficient in tiger population |
 | |
| *p<0.05 | |
| Table 2: | Number of animal affected with disease conditions |
 | |
| *p<0.05 | |
| Table 3: | χ2-test of heterogenity for colour, sex and inbreeding coefficient with respect to survibility |
 | |
| *p<0.05 | |
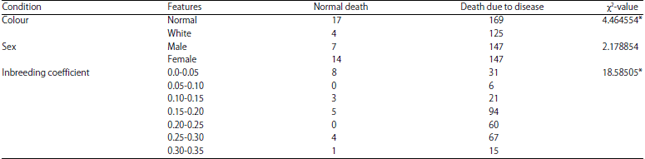
The diseases that were most affected due to inbreeding leading to death of an animal were: Shock stress, exertions, accidental injury, nephritis and senility. Cassinello25 suggested that death in inbred gazelles was due to reduce in resistance to pathogenic diseases. Inbred animals cannot cope up with the changing environment like fluctuating temperature, limited feed, unpredictable rainfall etc., by Falk and Holsinger28. The above four diseases were the product of improper environment. Thus death due to disease condition is significantly associated with inbreeding of tiger (Table 3). Supporting to this findings Miller and Lambert29 postulated that Black Robin population with no evidence of viral, protozoan or bacterial infections could still be susceptible to novel infection. Frankham et al.17 argued by explaining that one surviving population of Black Robin is already in a vulnerable condition, as evidence of its viability is like existence of one 80 years old smoker as evidence that smoking is harmless.
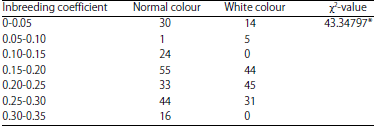
The white colour tiger, a product of homozygosity due to inbreeding was susceptible to different maladies as reported earlier Warrick7. The present finding (Table 3) supports his opinion. Likewise, inbreeding had an associationship with white colour of tigers (Table 4) confirms the findings of Carney30.
| Table 4: | χ2-test of heterogenity for colour with respect to inbreeding coefficient |
 | |
| *p<0.05 | |
CONCLUSION
Inbreeding has adverse effect on surrvibility of tigers that may lead to its extinction. In order to save tigers, planned breeding should be made, avoiding brother-sister, father-daughter and mother-son mating. The studbook should be properly maintained in a Zoo. The white coloration in tigers invites inbreeding as well as different maladies. So, temptation to produce more white tigers must be avoided. But inbred animal with better care and management now survive in captivity that would have died in wild condition. Although, it is applicable to the inbred animals upto 1 year of age thereafter inbreeding depression starts to play its role in the longevity of the young ones.
ACKNOWLEDGMENT
The authors are thankful to authority of Nandankanan Zoo for providing necessary support to conduct the research.
REFERENCES
- Hedrick, P.W., 1994. Purging inbreeding depression and the probability of extinction: Full-sib mating. Heredity, 73: 363-372.
CrossRefDirect Link - Hedrick, P.W. and S.T. Kalinowski, 2000. Inbreeding depression in conservation biology. Annu. Rev. Ecol. Syst., 31: 139-162.
CrossRefDirect Link - Keller, L.F. and D.M. Waller, 2002. Inbreeding effects in wild populations. Trends Ecol. Evol., 17: 230-241.
CrossRefDirect Link - Sarre, S.D. and A. Georges, 2009. Genetics in conservation and wildlife management: A revolution since Caughley. Wildlife Res., 36: 70-80.
CrossRefDirect Link - Rabon, Jr. D.R. and W. Waddell, 2010. Effects of inbreeding on reproductive success, performance, litter size, and survival in captive red wolves (Canis rufus). Zoo Biol., 29: 36-49.
CrossRefPubMedDirect Link - Warrick, D.M., 2010. Inbreeding depression in captive white tigers: Methods for purifying tiger lineages. Zoo's Print, 25: 7-15.
Direct Link - Xu, X., G.X. Dong, X.S. Hu, L. Miao and X.L. Zhang et al., 2013. The genetic basis of white tigers. Curr. Biol., 23: 1031-1035.
CrossRefDirect Link - Snedecor, G.W. and W.G. Cochran, 1967. Statistical Methods. 6th Edn., Oxford and IBH Publishing Co. Pvt. Ltd., New Delhi, India, Pages: 593.
Direct Link - Westemeier, R.L., J.D. Brawn, S.A. Simpson, T.L. Esker and R.W. Jansen et al., 1998. Tracking the long-term decline and recovery of an isolated population. Science, 282: 1695-1698.
CrossRefDirect Link - Madsen, T., R. Shine, M. Olsson and H. Wittzell, 1999. Conservation biology: Restoration of an inbred adder population. Nature, 402: 34-35.
CrossRefDirect Link - Craig, J., S. Anderson, M. Clout, B. Creese and N. Mitchell et al., 2000. Conservation issues in New Zealand. Annu. Rev. Ecol. Syst., 31: 61-78.
CrossRefDirect Link - Templeton, A.R. and B. Read, 1984. Factors eliminating inbreeding depression in a captive herd of speke's gazelle (Gazella spekei). Zoo Biol., 3: 177-199.
CrossRefDirect Link - Dwyer, C.M., 2008. Genetic and physiological determinants of maternal behavior and lamb survival: Implications for low-input sheep management. J. Anim. Sci., 86: E246-E258.
PubMed - Boakes, E.H., J. Wang and W. Amos, 2007. An investigation of inbreeding depression and purging in captive pedigreed populations. Heredity, 98: 172-182.
CrossRefDirect Link - Ralls, K.J., J.D. Ballou and A.D. Templeton, 1988. Estimates of lethal equivalents and the cost of inbreeding in mammals. Conserv. Biol., 2: 185-193.
CrossRefDirect Link - Cassell, B.G., V. Adamec and R.E. Pearson, 2003. Maternal and fetal inbreeding depression for 70-day nonreturn and calving rate in Holsteins and Jerseys. J. Dairy Sci., 86: 2977-2983.
CrossRefDirect Link - McParland, S., F. Kearney and D.P. Berry, 2009. Purging of inbreeding depression within the Irish Holstein-Friesian population. Genet. Sel. Evol., Vol. 41.
CrossRefDirect Link - Ballou, J.D., 1997. Ancestral inbreeding only minimally affects inbreeding depression in mammalian population. J. Hered., 88: 169-178.
Direct Link - Cassinello, J., 2005. Inbreeding depression on reproductive performance and survival in captive gazelles of great conservation value. Biol. Conserv., 122: 453-464.
CrossRefDirect Link - Van Coillie, S., P. Galbusera, A.D. Roeder, W. Schempp and J.M.G. Stevens et al., 2008. Molecular paternity determination in captive bonobos and the impact of inbreeding on infant mortality. Anim. Conserv., 11: 306-314.
CrossRefDirect Link - Miller, H.C. and D.M. Lambert, 2004. Genetic drift outweighs balancing selection in shaping post-bottleneck major histocompatibility complex variation in New Zealand robins (Petroicidae). Mol. Ecol., 13: 3709-3721.
CrossRefDirect Link