
Jianchang Wang
Inspection and Quarantine Technical Center, Hebei Entry-Exit Inspection and Quarantine Bureau, 318 Heping West Rd, Xinhua District,
Shi Jiazhuang, 050051 Hebei, China
Jinfeng Wang
Inspection and Quarantine Technical Center, Hebei Entry-Exit Inspection and Quarantine Bureau, 318 Heping West Rd, Xinhua District,
Shi Jiazhuang, 050051 Hebei, China
Libing Liu
Inspection and Quarantine Technical Center, Hebei Entry-Exit Inspection and Quarantine Bureau, 318 Heping West Rd, Xinhua District,
Shi Jiazhuang, 050051 Hebei, China
Ruiying Chai
College of Veterinary Medicine, Agricultural University of Hebei, 38 Lingyusi Street, Baoding, 071001 Hebei, China
Wanzhe Yuan
College of Veterinary Medicine, Agricultural University of Hebei, 38 Lingyusi Street, Baoding, 071001 Hebei, China
Asian Journal of Animal and Veterinary Advances
Year: 2016 | Volume: 11 | Issue: 7 | Page No.: 405-410







ABSTRACT
Background: Schmallenberg virus (SBV), a novel orthobunyavirus was first identified in Germany in 2011. It primarily affects ruminants, e.g., cattle, sheep and goat, etc., causing schmallenberg disease. So far multiple diagnostic methods such as antibody testing and real time reverse transcription polymerase chain reaction (RT-PCR) have been developed for SBV detection. While antibody testing can be used for blood samples, it is not suitable for the test of other samples such as semen, cerebrum and spinal cord. Real time RT-PCR involves complex techniques and requires an expensive thermocycler. Loop-mediated isothermal amplification (LAMP), a novel isothermal gene amplification technique has been explored for the detection of diverse pathogens. The LAMP has several advantages over PCR, including the use of a water bath and being less sensitive to inhibitors that often appear in test samples. In this study, RT-LAMP was explored for SBV detection. Materials and Methods: Viral genes including SBV genomic RNA were used as template. A set of primers targeting the S gene were designed. The RT-LAMP reactions were performed at 61°C in a water bath. Sensitivity of the RT-LAMP assay was determined and compared with that of real time RT-PCR by using different copy numbers of SBV genomic RNA as template. Specificity was evaluated by using different vial genes as template. The RT-LAMP assay was further validated with SBV-spiked animal blood samples. Results: The detection limit of RT-LAMP was 10 copies of SBV genomic RNA, comparable to real time RT-PCR. Specificity analysis revealed that RT-LAMP detected SBV but did not amplify other viruses commonly found in ruminants. Like some real time RT-PCR assays, RT-LAMP cross-detected shamonda virus and Australian douglas virus, members of the Bunyaviridae family, which required further DNA sequencing for differentiation. Validation results showed that RT-LAMP had a complete agreement with real time RT-PCR in detecting SBV in spiked blood samples. Conclusion: A one-step RT-LAMP method was successfully developed. The RT-LAMP assay is comparable to real time RT-PCR in term of assay sensitivity and specificity but it is simpler and more cost-effective. The RT-LAMP method might be potentially applied to replace real time RT-PCR for SBV detection in resource-limited settings.
PDF Abstract XML References Citation
How to cite this article
Jianchang Wang, Jinfeng Wang, Libing Liu, Ruiying Chai and Wanzhe Yuan, 2016. Rapid and Sensitive Detection of Schmallenberg Virus by a One-step Reverse Transcription Loop-mediated Isothermal Amplification Method. Asian Journal of Animal and Veterinary Advances, 11: 405-410.
DOI: 10.3923/ajava.2016.405.410
URL: https://scialert.net/abstract/?doi=ajava.2016.405.410
DOI: 10.3923/ajava.2016.405.410
URL: https://scialert.net/abstract/?doi=ajava.2016.405.410
INTRODUCTION
Since schmallenberg virus (SBV) was first detected in Germany in 20111, it has emerged in many other countries2. The SBV is an enveloped, negative-sense, segmented, single-stranded RNA virus with its genome consisting of three segments, large (L), medium (M) and small (S)3. It belongs to the Bunyaviridae family and primarily affects ruminants, e.g., cattle, sheep and goat, etc., causing Schmallenberg disease that is characterized by fever, drop in milk production and diarrhea4-6. Other symptoms include abortions, stillbirths and severe fetal malformations7,8. Diagnostic methods such as antibody testing9,10 and real time reverse transcription polymerase chain reaction (RT-PCR)11-13 have been developed for SBV detection. Antibody testing can be used for blood samples but it is not suitable for the test of other samples such as semen, cerebrum and spinal cord. Real time RT-PCR involves complex techniques and requires an expensive thermocycler. Loop-mediated isothermal amplification (LAMP), a novel isothermal gene amplification technique has been explored for the detection of diverse pathogens14,15. It uses a strand-displacing DNA polymerase along with two internal primers (FIP and BIP) and two outer primers (F3 and B3)16. The LAMP has several advantages over PCR, including the use of a water bath and being less sensitive to inhibitors that often appear in test samples. Therefore, RT-LAMP was explored for SBV detection in the present study.
MATERIALS AND METHODS
The conserved region of the SBV S gene was identified through comparison of genomic sequences of SBV [GenBank: HE649914.1], akabane virus (AKAV) [GenBank: NC_009896.1], shamonda virus (SHAV) [GenBank: NC_018464.1], aino virus (AINOV) [GenBank: NC_018460.1], douglas virus (DOUV) [GenBank: AF362393.1] and simbu virus (SIMV) [GenBank: AF362397.1]. Primer explored V4 software (http://primerexplorer.jp/e/) was used to design the RT-LAMP primers targeting the SBV S segment (Table 1). All primers were synthesized by Sangon Biotech (Shanghai, China).
The LAMP reaction was optimized with a total volume of 25 μL consisting of 2.5 μL of 10×buffer, 1.0 mM of MgCl2, 0.4 mM of dNTPs, 0.32 U μL–1 of bst DNA polymerase, 0.8 μM each of inner primers, 0.2 μM each of outer primers, 0.2 μL–1 of AMV-reverse transcriptase and 0.8 μL–1 RNase inhibitor. Nuclease-free water was used to adjust the volume. All components were obtained from new England Biolabs (Beijing, China) unless otherwise indicated. The reaction was performed in a water bath at 61°C for 40 min followed by inactivation at 80°C for 2 min. Afterwards, amplified products were electrophoresed on 2.0% (w/v) agarose gel, stained with Goldview (Dongsheng Biotech Co., Ltd., Guangzhou, China) and visualized under UV light.
Sensitivity test was done using SBV genomic RNA as template (SBV RNA was kindly provided by Dr. Bernd Hoffmann from the Friedrich-Loeffler-Institute, Germany). The RNA was diluted in a 10-fold serial dilution manner to achieve concentrations ranging from 1.0×107 to 1.0×100 copies μL–1. Different copy numbers of RNA were then amplified by RT-LAMP reactions. For comparison, different copy numbers of RNA were also amplified using the virotype SBV RT-PCR kit (Qiagen, Shanghai, China) following the instructions provided in the Kit.
To analyze the specificity of the RT-LAMP assay, 10 ng of the genomic nucleotides of SBV, AKAV, foot-and-mouth disease virus (FMDV), Bovine Viral Diarrhea Virus (BVDV), peste des petits ruminants virus (PPRV) and bovine herpesvirus-1 (BoHV-1) was amplified with LAMP (genomic nucleotides of AKAV, FMDV, BVDV, PPRV and BoHV-1 were prepared in this laboratory). Ten nanogram of plasmids containing the S gene of SHAV, DOUV, AINOV and SIMV was also amplified with LAMP (all plasmids were kindly provided by Dr. Yongning Zhang, the Institute of Animal Quarantine, Chinese Academy of Inspection and Quarantine).
The RT-LAMP assay was validated with animal blood samples. Collection and use of animal blood were approved by the Ethics Committee of Agricultural University of Hebei, China. Blood samples from 50 sheep and 50 cattle were collected. Animals were restrained with minimal distress and 5 mL of blood was collected into an ETDA-vacutainer (BD, Franklin Lakes, USA) from the tail vein of cattle or the jugular vein of sheep according to standard venipuncture procedures.
| Table 1: | Sequences of SBV RT-LAMP primers |
 | |
| aNumbers represent the nucleotide position within the S gene of SBV (GenBank accession number: HE649914.1) | |
Spiked blood samples were prepared as follows, schmallenberg virus-like particles containing the S segment (obtained from the Chinese Academy of Inspection and Quarantine, Beijing, China) was diluted in a 10-fold serial dilution manner and then each dilution of viral particles was added to 1 mL cattle blood free of SBV to achieve concentrations ranging from 1×104 to 1×100 PFU mL–1 blood. Total RNA was extracted from 1 mL blood, directly-collected or artificially spiked, using an RNeasy Mini kit (Qiagen, Shanghai, China) according to the manufacturer’s instructions. The RNA was eluted in 20 μL nuclease-free H2O and 5 μL RNA was tested for the presence of SBV by the RT-LAMP assay as well as the virotype SBV RT-PCR kit.
RESULTS
Figure 1a shows that, the detection limit of the RT-LAMP assay was 10 copies per reaction. The detection limit of the Qiagen commercial RT-PCR Kit was also 10 copies per reaction (Fig. 1b). Similar results were observed in 3 independent experiments.
Figure 2a shows that, only SBV RNA was amplified while others were not. Using 10 ng plasmids containing the S gene of SHAV, DOUV, AINOV and SIMV as templates, it was shown that LAMP cross-detected SHAV and DOUV (Fig. 2b).
All blood samples collected from animals were SBV negative as determined by both RT-LAMP and the Qiagen SBV RT-PCR kit. The detection limit for the spiked blood was 100 PFU mL–1 blood as determined by both the RT-LAMP assay and the Qiagen RT-PCR kit (Table 2).
DISCUSSION
Multiple assays have been developed for SBV detection. Antibody testing approaches for SBV detection have been described9,10. These methods can detect SBV antibodies in blood but they are not suitable for the test of other samples such as semen, cerebrum and spinal cord. Therefore, molecular methods such as real time RT-PCR have been rapidly established for the detection of SBV genome11-13. Semi-quantitative RT-PCR was used to diagnose SBV infection in biting midges vectors, which produced results in a rapid and straightforward way but the sensitivity was limited17. Fischer et al.12 developed several SBV real time RT-PCR assays using different primers targeting the M (SBV-M1 assay), the L (SBV-L1 and SBV-L1.4 assays) and the S (SBV-S3 assay) segment. After testing with the SBV genomic RNA and comparing the results, it was found that the SBV-S3 assay was the most suitable system with the highest sensitivity (detection limit: 1-10 copies per reaction) and reliability for the detection of SBV genome.
| Table 2: | Validation results |
 | |
| 6/6: 6 out of 6 samples were tested positive, 0/6: 0 out of 6 samples were tested positive | |
Based on these findings, primers targeting the S gene were designed for this study and the results showed that both RT-LAMP and the Qiagen real time RT-PCR kit had the detection limit of 10 copies per reaction. More recently, the development of a one-step multiplex real time RT-PCR assay was reported and showed a detection limit of 2.4 copies of SBV genome18. Of note, the multiplex real time RT-PCR method was not tested with DOUV and SHAV18. Recombinase polymerase amplification has also been investigated for SBV detection. However, a big problem with this method is its low sensitivity (1000 times less sensitive than real time RT-PCR19.
The specificity analysis demonstrated that RT-LAMP amplified SBV but did not amplify other viruses commonly found in ruminants, i.e., AKAV, FMDV, BVDV, PPRV and BoHV-1. However, cross-detection of DOUV and SHAV was observed. There had been no schmallenberg cases reported in China while this study was conducted, therefore spiked blood samples were tested and the results from the RT-LAMP assay were in complete agreement with those obtained using the Qiagen SBV RT-PCR kit.
While this method was under development, a RT-LAMP assay for SBV detection was published19. Of note, primers targeting L gene were used in that study and the results showed that the RT-LAMP assay did not detect any of the related simbu serogroup viruses. However, the sensitivity was significantly reduced (100 times less sensitive than real time RT-PCR). Therefore, the researchers suggested that their RT-LAMP assay was attractive for confirmatory diagnosis and rapid differentiation of SBV but not suitable for large screening investigations due to low sensitivity. In term of sensitivity and specificity, our method appears to be complementary to that described by Aebischer et al.19.
One challenge for the development of LAMP-based assays is the selection of primers. As 4 primers are used in LAMP reactions with each inner primer containing both sense and antisense sequences, non-specific primer-primer interaction can occur and limit the availability of primers for target amplification20.
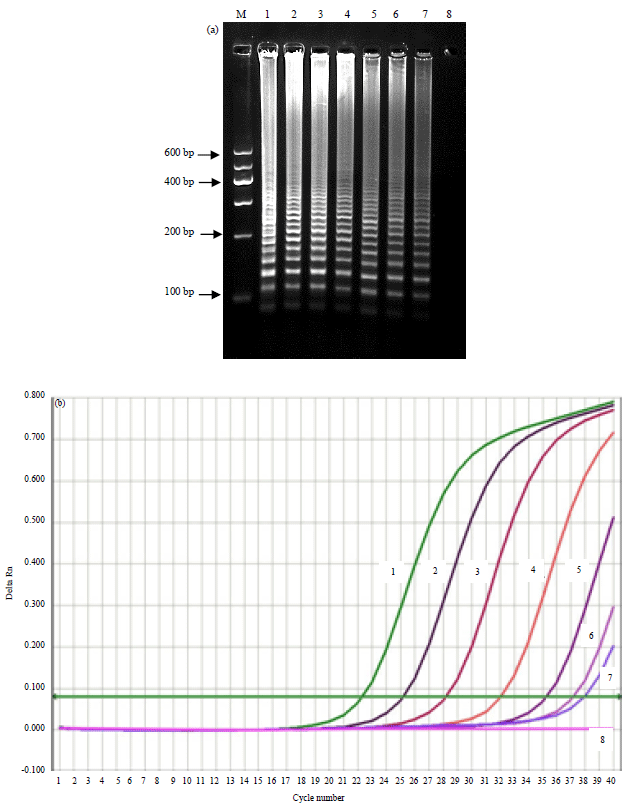 | |
| Fig. 1(a-b): | Sensitivity analysis. Different copy numbers of SBV RNA were used as the template for RT-LAMP and real time RT-PCR, (a) Amplified products from RT-LAMP reactions as analyzed by agarose gel electrophoresis and (b) A representative amplification curve graph of real time RT-PCR. Template copy numbers were as follows: Lane 1 or curve 1: 1.0×107 copies, Lane 2 or curve 2: 1.0×106 copies, Lane 3 or curve 3: 1.0×105 copies, Lane 4 or curve 4: 1.0×104 copies, Lane 5 or curve 5: 1.0×103 copies, Lane 6 or curve 6: 1.0×102 copies, Lane 7 or curve 7: 1.0×101 copies, Lane 8 or curve 8: 1.0×100 copies and M: DNA marker. All experiments were repeated three times and similar results were obtained |
 | |
| Fig. 2(a-b): | Specificity analysis, (a) No cross detection of viruses commonly found in the ruminant by RT-LAMP was observed, M: DNA marker, Lane 1: AKAV, Lane 2: FMDV, Lane 3: BVDV, Lane 4: PPRV, Lane 5: BoHV-1 and Lane 6: SBV and (b) Cross detection of SHAV and DOUV was observed with RT-LAMP, Lane 1: Non-template control, Lane 2: SBV, Lane 3: SHAV, Lane 4: DOUV, Lane 5: AKAV, Lane 6: AINOV and Lane 7: SIMV. All experiments were repeated three times and similar results were obtained |
In this study, 5 primer sets targeting the S segment was designed only 1 set (Table 1) successfully amplified the SBV RNA. Another shortcoming of this RT-LAMP method is that, like some real time RT-PCR assays12, it cross-detected SHAV and DOUV, members of the Bunyaviridae family, which requires further DNA sequencing for differentiation.
All gene-amplification based methods developed so far for SBV detection have pros and cons, e.g., the LAMP assay recently described elsewhere had high specificity but low sensitivity19 the method reported here showed high sensitivity but cross-detected DOUV and SHAV. In practice, combined use of these assays might provide better testing results.
CONCLUSION
A one-step RT-LAMP method was successfully developed. The RT-LAMP assay is comparable to real time RT-PCR in term of assay sensitivity and specificity but it is simpler and more cost-effective. The RT-LAMP method might be potentially applied to replace real time RT-PCR for SBV detection in resource-limited settings.
ACKNOWLEDGMENTS
The authors sincerely thank Dr. Bernd Hoffmann for providing the SBV RNA sample and Dr. Yongning Zhang for providing all plasmids used in this study. This study was supported by the Science and Technology Project (Grant No. 2013IK054), the General Administration of Quality Supervision, Inspection and Quarantine, China.
REFERENCES
- Hoffmann, B., M. Scheuch, D. Hoper, R. Jungblut and M. Holsteg et al., 2012. Novel orthobunyavirus in cattle, Europe, 2011. Emerg. Infect. Dis., 18: 469-472.
CrossRefPubMedDirect Link - Zientara, S. and C. Ponsart, 2014. Viral emergence and consequences for reproductive performance in ruminants: Two recent examples (bluetongue and Schmallenberg viruses). Reprod. Fertil. Dev., 27: 63-71.
CrossRefDirect Link - Coupeau, D., F. Claine, L. Wiggers, B. Martin, N. Kirschvink and B. Muylkens, 2013. Characterization of messenger RNA termini in Schmallenberg virus and related Simbuviruses. J. Gen. Virol., 94: 2399-2405.
CrossRefDirect Link - Garigliany, M.M., C. Bayrou, D. Kleijnen, D. Cassart, S. Jolly, A. Linden and D. Desmecht, 2012. Schmallenberg virus: A new Shamonda/Sathuperi-like virus on the rise in Europe. Antiviral Res., 95: 82-87.
CrossRefDirect Link - Beer, M., F.J. Conraths and W.H.M. van der Poel, 2013. Schmallenberg virus-a novel orthobunyavirus emerging in Europe. Epidemiol. Infect., 141: 1-8.
CrossRefDirect Link - Conraths, F.J., M. Peters and M. Beer, 2013. Schmallenberg virus, a novel orthobunyavirus infection in ruminants in Europe: Potential global impact and preventive measures. N. Z. Vet. J., 61: 63-67.
CrossRefDirect Link - Herder, V., P. Wohlsein, M. Peters, F. Hansmann and W. Baumgartner, 2012. Salient lesions in domestic ruminants infected with the emerging so-called Schmallenberg virus in Germany. Vet. Pathol., 49: 588-591.
CrossRefDirect Link - Wernike, K., M. Holsteg, H. Schirrmeier, B. Hoffmann and M. Beer, 2014. Natural infection of pregnant cows with Schmallenberg virus-a follow-up study. PloS ONE, Vol. 9.
CrossRefDirect Link - Van der Heijden, H.M.J.F., R.J. Bouwstra, M.H. Mars, W.H.M. van der Poel, G.J. Wellenberg and C. van Maanen, 2013. Development and validation of an indirect enzyme-linked immunosorbent assay for the detection of antibodies against Schmallenberg virus in blood samples from ruminants. Res. Vet. Sci., 95: 731-735.
CrossRefDirect Link - Mansfield, K.L., S.A. la Rocca, M. Khatri, N. Johnson, F. Steinbach and A.R. Fooks, 2013. Detection of Schmallenberg virus serum neutralising antibodies. J. Virol. Methods, 188: 139-144.
CrossRefDirect Link - Bilk, S., C. Schulze, M. Fischer, M. Beer, A. Hlinak and B. Hoffmann, 2012. Organ distribution of Schmallenberg virus RNA in malformed newborns. Vet. Microbiol., 159: 236-238.
CrossRefDirect Link - Fischer, M., H. Schirrmeier, K. Wernike, A. Wegelt, M. Beer and B. Hoffmann, 2013. Development of a pan-Simbu real-time reverse transcriptase PCR for the detection of Simbu serogroup viruses and comparison with SBV diagnostic PCR systems. Virol. J., Vol. 10.
CrossRefDirect Link - Schulz, C., W.H.M. van der Poel, C. Ponsart, A.B. Cay and F. Steinbach et al., 2015. European interlaboratory comparison of Schmallenberg Virus (SBV) real-time RT-PCR detection in experimental and field samples: The method of extraction is critical for SBV RNA detection in semen. J. Vet. Diagn. Invest., 27: 422-430.
CrossRefDirect Link - Fu, S., G. Qu, S. Guo, L. Ma and N. Zhang et al., 2011. Applications of loop-mediated isothermal DNA amplification. Applied Biochem. Biotechnol., 163: 845-850.
CrossRefPubMedDirect Link - Mori, Y., H. Kanda and T. Notomi, 2013. Loop-mediated isothermal amplification (LAMP): Recent progress in research and development. J. Infect. Chemother., 19: 404-411.
CrossRefPubMedDirect Link - Notomi, T., H. Okayama, H. Masubuchi, T. Yonekawa, K. Watanabe, N. Amino and T. Hase, 2000. Loop-mediated isothermal amplification of DNA. Nucl. Acids Res., 28: e63-e63.
CrossRefDirect Link - Veronesi E., M. Henstock, S. Gubbins, C. Batten and R. Manley et al., 2013. Implicating Culicoides biting midges as vectors of Schmallenberg virus using semi-quantitative RT-PCR. PLoS ONE, Vol. 8.
CrossRefDirect Link - Lee, J.H., H.J. Seo, J.Y. Park, S.H. Kim and Y.S. Cho et al., 2015. Detection and differentiation of Schmallenberg, Akabane and Aino viruses by one-step multiplex reverse-transcriptase quantitative PCR assay. BMC Vet. Res., Vol. 11.
CrossRefDirect Link - Aebischer, A., K. Wernike, B. Hoffmann and M. Beer, 2014. Rapid genome detection of Schmallenberg virus and bovine viral diarrhea virus by use of isothermal amplification methods and high-speed real-time reverse transcriptase PCR. J. Clin. Microbiol., 52: 1883-1892.
CrossRefDirect Link - Gandelman, O., R. Jackson, G. Kiddle and L. Tisi, 2011. Loop-mediated amplification accelerated by stem primers. Int. J. Mol. Sci., 12: 9108-9124.
CrossRefDirect Link