
Hamza I. Eid
Deptartment of Bacteriology, Mycology and Immunology, Faculty of Veterinary Medicine, Suez Canal University, 41522 Ismailia, Egypt
Abdelazeem M. Algammal
Deptartment of Bacteriology, Mycology and Immunology, Faculty of Veterinary Medicine, Suez Canal University, 41522 Ismailia, Egypt
LiveDNA: 20.14499
Soad A. Nasef
Reference Laboratory for Veterinary Quality Control on Poultry Production (Dokki), Animal Health Research Institute, 12618 Giza, Egypt
Wael K. Elfeil
Department of Avian and Rabbit Medicine, Faculty of Veterinary Medicine, Suez Canal University, 41522 Ismailia, Egypt
LiveDNA: 20.5311
Ghada H. Mansour
Reference Laboratory for Veterinary Quality Control on Poultry Production (Dokki), Animal Health Research Institute, 12618 Giza, Egypt
Asian Journal of Animal and Veterinary Advances
Year: 2016 | Volume: 11 | Issue: 6 | Page No.: 350-356







ABSTRACT
Background: Avian pathogenic E. coli cause serious disease in chickens, which mainly chracterised by airsaculitis, perihepatitis and pericarditis resulting in large economic losses in poultry industry worldwide. Methodology: In order to investigate the genetic variation among different E. coli strains isolated from chickens, one hundred diseased broiler chickens at Sharkia province, Egypt were examined. Liver and heart blood samples were collected from each bird and subjected to bacteriological examination, where the prevalence of E. coli was 60% from the total collected samples. Escherichia coli strains were serogrouped. The PCR was used for detection of Shiga-like toxins genes (stx1 and stx2), attaching and effacing (eaeA) gene and enterohaemolysin gene (hly) in the typable isolated E. coli strains. Results: The results showed that the isolated strains belonged to 11 serogroups including O1, O2, O26, O44, O55, O78, O111, O119, O125, O127 and O128. Untypable strains were also recovered. The detected virulence genes were stx1 in all E. coli strains (100%), stx2 in 17 strains (47.2%), eaeA in 12 strains (33.3%) and hly only in three strains (8.3%). Conclusion: In conclusion, the combination of genotypic and phenotypic analysis of E. coli is more valuable as an epidemiological tool for identification of isolates. This study established the presence of stx1 and stx2 containing E. coli in chickens.
PDF Abstract XML References Citation
How to cite this article
Hamza I. Eid, Abdelazeem M. Algammal, Soad A. Nasef, Wael K. Elfeil and Ghada H. Mansour, 2016. Genetic Variation among Avian Pathogenic E. coli Strains Isolated from Broiler Chickens. Asian Journal of Animal and Veterinary Advances, 11: 350-356.
DOI: 10.3923/ajava.2016.350.356
URL: https://scialert.net/abstract/?doi=ajava.2016.350.356
DOI: 10.3923/ajava.2016.350.356
URL: https://scialert.net/abstract/?doi=ajava.2016.350.356
INTRODUCTION
Escherichia coli is one of the major infectious agents that can cause tremendous problems in chickens. Pathogenic E. coli strains can be classified into intestinal and extra-intestinal E. coli according to their virulence factors and clinical symptoms1. Colibacillosis is one of the most serious diseases affecting poultry resulting in severe economic losses due to mortalities, weight loss, carcass condemnations and costs of treatment and preventive measures. In addition, Avian Pathogenic E. coli (APEC), the causitive agent of colibacillosis, is proved to be a zoonotic pathogen2.
Presently, diarrheagenic E. coli strains are classified into 6 main pathogenic types according to their virulence factors, including enteropathogenic E. coli (EPEC), enterotoxogenic E. coli (ETEC) that produce heat labile and heat stable enterotoxins, enteroinvasive E. coli (EIEC) that simulates Shigella strains in their ability to invade the intestinal epithelial cells, enterohaemorrhagic E. coli (EHEC) that produce Shiga-like toxins, enteroaggregative strain (EAggEC) and Diffusively adherent E. coli (DAEC)3. Shiga toxin-producing E. coli (STEC) is the most important group of foodborne pathogens4. The EHEC strains are one of the subsets of Shiga toxin (Stx)-producing E. coli (STEC) strains5,6.
The most dangerous E. coli serotype associated with EHEC is O157:H7, which is chracterised by the ability to cause severe disease in humans, such as Hemorrhagic Colitis (HC), Hemolytic Uremic Syndrome (HUS) and Thrombotic Thrombocytopenic Purpura (TTP)7. The major virulence factors involved in STEC infection are the powerful Shiga toxins, which are classified into 2 main groups: Stx1 and Stx2. Other important virulence factors of STEC are intimin (encoded by eaeA gene), a plasmid-encoded enterohaemolysin and, in strains lacking eae, an autoagglutinating adhesin8.
The aim of this study was to investigate the genetic variation among avian pathogenic E. coli strains isolated from chickens. This aim was achieved by isolation and identification of E. coli, serogrouping of isolated E. coli strains and detection of virulence genes in the isolated E. coli strains by using PCR.
MATERIALS AND METHODS
Sampling: A total of 100 samples (liver and heart blood from each bird) from 100 diseased broiler chickens were collected under complete aseptic condition separately in sterile plastic bags and transferred to the laboratory for bacteriological examination.
Samples were collected from cases exhibiting perihepatitis, pericarditis and airsacculitis. Liver and heart blood samples were collected based on clinical signs and pathogenomic lesions observed during post mortem examination of broiler chickens at Reference Laboratory for Veterinary Quality Control on Poultry Production in Sharkia province.
Isolation and identification of E. coli: For enrichment, one gram of each collected sample was aseptically added to 9 mL of buffered peptone water and incubated at 37°C for 24 h. A Loopful from the incubated broth was streaked onto MacConkey s agar plates and incubated at 37°C for 24 h. Lactose fermenting colonies were picked up and streaked onto EMB agar and incubated at 37°C for 24 h. Metallic green sheen colored colonies on EMB were sub cultured on Nutrient agar slant and incubated at 37°C for 24 h for storage at 4°C in the refrigerator for further studies and characterization. Suspected E. coli isolates were identified morphologically by Gram stain, motility test and biochemically according to Quinn et al.9.
Serotyping of E. coli isolates: The somatic (O) antigen was determined by slide agglutination test according to Edwards and Ewing10, while Flagellar (H) antigen serotyping was carried out using tube method according to Davies and Wray11. Serotyping of the isolated E. coli strains was carried out at The Serology Unit in Animal Health Research Institute in Dokki, Cairo, Egypt.
In vitro pathogenicity testing
Congo red test: Various serotypes were tested for pathogenicity based on Congo red dye binding test as described by Berkhoff and Vinal12. Trypticase soya agar was supplemented with 0.003% Congo red dye (Sigma) and 0.15% bile salts. Each isolate was streaked on a separate plate and incubated at 37°C for 24 h. After 24 h incubation, plates were left at room temperature for 48 h to facilitate annotation of results. Invasive E. coli were identified by their ability to take up Congo red dye. Appearance of red colonies was recorded as a positive reaction. Negative colonies did not bind the dye and remained white or grey.
Hemolysis production test: Escherichia coli strains were propagated on tryptose blood agar (Difco) plates supplemented with 5% defibrinated washed sheep blood and incubated at 37°C for 24 h. Then plates were examined for "greening" or clearing of the agar around areas of bacterial growth as an indication of alpha or beta hemolytic activity Livezey and Zusi13.
PCR detection of stx 1, stx2, eaeA and hly genes in the isolated E. coli strains: Thirty sex typable E. coli strains were subjected to PCR for detection of stx1, stx2, eaeA and hly genes.
Extraction of DNA: Escherichia coli strains were cultured in brain heart infusion broth (LAB M-LAB 49); at 37°C for 18-24 h. DNA was extracted using QIAamp DNA Mini Kit (Catalogue No. 51304).
Polymerase chain reaction: Primers used in PCR were shown in Table 1.
DNA samples were tested in 50 μL. Reaction volume in a 0.2 mL PCR tube, containing PCR buffer (50 mM KCl, 10 mM tris-HCl, 1 mM MgCl2) each dNTPS (Deoxy nucleotide triphosphate) 200 μM each (dATP, dGTP, dCTP and dTTP), Two primer pairs each at 50 picomol/reaction and 0.5 of taq DNA polymerase. Thermal cycling in a programmable heating block (Coy vorporation, Grasslake, Michan, USA) was done14.
PCR recycling conditions:
| • | The stx1 gene according to Sahilah et al.15: Initial denaturation at 94°C for 7 min, 35 cycles (denaturation at 94°C for 30 sec, annealing at 51°C for 30 sec, extension at 72°C for 30 sec), final extension 72°C for 7 min |
| Table 1: | Oligonucleotide primers sequences |
 | |
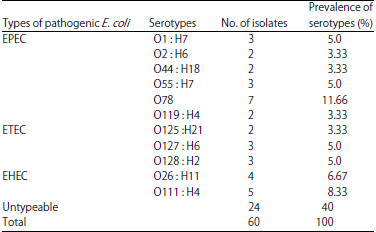
| Table 2: | Prevalence of detected serotypes based on total No. of E. coli strains |
 | |
| • | The stx2 gene according to Dipineto et al.16: Initial denaturation at 94°C for 10 min, 35 cycles (denaturation at 94°C for 1 min, annealing at 58°C for 1 min, extension at 72°C for 1 min), final extension 72°C for 10 min |
| • | The hly gene according to Piva et al.8: Initial denaturation at 94°C for 15 min, 35 cycles (denaturation at 94°C for 1 min, annealing at 60°C for 1 min, extension at 72°C for 15 min), final extension 72°C for 12 min |
| • | The eaeA gene according to Bisi-Johnson et al.17: Initial denaturation at 94°C for 7 min, 40 cycles (denaturation at 94°C for 30 sec, annealing at 51°C for 30 sec, extension at 72°C for 30 sec), final extension 72°C for 7 min |
c-Screening of PCR products: Ten microliter of amplified PCR product was analyzed by electrophoresis on a 2% agarose gel stained with 0.5 μg of ethedium bromide/mL. Electrophoresis was carried out in 1X TAE buffer at 80 volt for 1 h14,18. Gels were visualized under UV transilluminator (UVP, UK) and photographed.
RESULTS
Prevalence and serogruoping of E. coli: In the present study, the prevalence of E. coli was (60%), the isolated E. coli strains were typed serologically into 11 different ‘O’ groups; O78 (11.66%), O111: H4 (8.33%), O26: H11 (6.67%), O1: H7, O55: H7, O127: H6 and O128: H2 (5% for each), O2: H6, O44: H18, O119: H4 and O125: H21 (3.33% for each) and 24 E. coli strains were untypable (40%) as shown in Table 2.
In vitro pathogenicity testing
Congo red test: In vitro pathogenicity testing (Congo red binding assay) indicates that (91.7%) (n = 55 strains) of the isolated E. coli strains were positive and only (8.3%) (n = 5) were negative.
Hemolysis on blood agar: Regarding to hemolysis production on blood agar; all the isolated strains were non hemolytic.
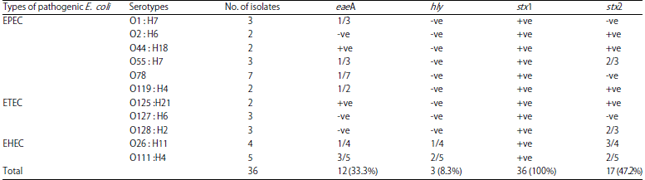
PCR detection of stx 1, stx2, eaeA and hly genes: As ahown in Table 3 and Fig. 1-4, 36 E. coli strains were subjected to PCR for detection of stx1, stx2, hly and eaeA genes. The PCR results revealed that:
| stx1 gene: | All E. coli strains (100%) were positive to stx1 gene |
| stx2 gene: | The 17 strains (47.2%) were positiv for stx2 gene including; eight EPEC strains [2 (O2: H6), 2 (O44: H18), 2 (O55: H7) and 2(O119: H4)]; four ETEC strains [ 2 (O125: H21) and 2 (O128: H2)]; five EHEC strains [3 (O26: H11) and 2 (O111: H4)] |
| eaeA gene: | The 12 strains (33.3%) were positive for eaeA gene including; six EPEC strains [ 1 (O1: H7), 2 (O44: H18), 1 (O55: H7), 1(O78) and 1(O119: H4)]; two ETEC [2 O125: H21)]; 4 EHEC strains [1 (O26: H11) and 3 (O111: H4)] |
| hly gene: | Only three strains (8.3%) were positive for hly gene including; three EHEC [1 (O26: H11) and 2 (O111: H4)] |
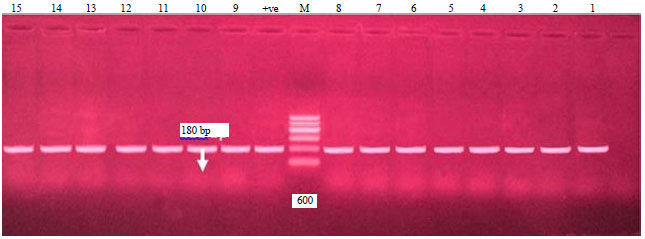 | |
| Fig. 1: | Electrophoretic pattern of the PCR products of stx1 gene, Agarose gel electrophoresis of the PCR products of stx1 gene (180 bp). Lane M: 100 bp DNA Ladder (100-600 bp), lanes (1-15) positive E. coli strains, positive control (lane +ve) and negative control (lane -ve) |
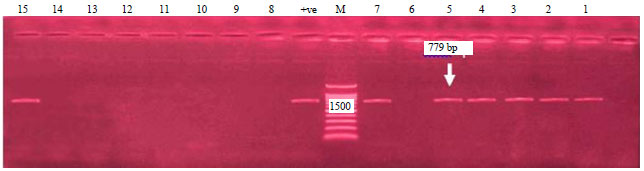 | |
| Fig. 2: | Electrophoretic pattern of the PCR products of stx2 gene, Agarose gel electrophoresis of the PCR products of stx2 gene (779 bp). Lane M: DNA marker (100-1500 bp), lanes (1, 2, 3, 4, 5, 7, 15) +ve E. coli strains, lanes (6, 8, 9, 10, 11, 12, 13, 14) –ve E. coli strains, positive control (lane +ve) and lane: negative control |
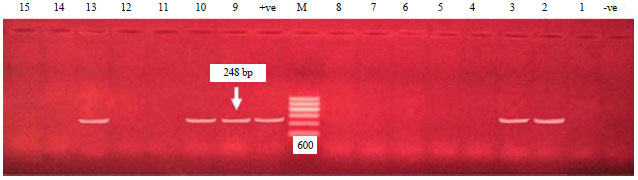 | |
| Fig. 3: | Electrophoretic pattern of the PCR products of eaeA gene, Agarose gel electrophoresis of the PCR products of eaeA gene (248 bp). Lane M: 100 bp DNA Ladder (100-600 bp), lanes (2, 3, 9, 10, 13): +ve E. coli strains, lanes (1, 4, 5, 6, 7, 8, 11, 12, 14, 15): -ve E. coli strains, positive control (lane +ve) and negative control (lane -ve) |
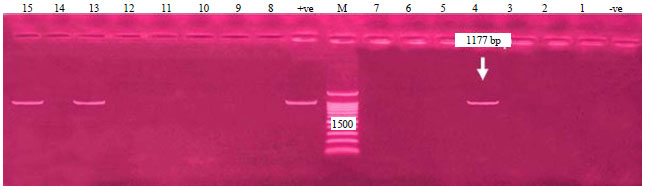 | |
| Fig. 4: | Electrophoretic pattern of the PCR products of hly gene, Agarose gel electrophoresis of the PCR products of hly gene (1177 bp). Lane M: DNA marker (100-1500 bp), lanes (4, 13, 15): +ve E. coli strains, lanes (1, 2, 3, 5, 6, 7, 8, 9, 10, 11, 12, 14): -ve E. coli strains, positive control (lane +ve) and negative control (lane -ve) |
| Table 3: | Prevalence of stx1, stx2, eaeA and hly genes among the isolated E. coli serotypes |
 | |
DISCUSSION
In the present study,the prevalence of E. coli was (60%) as shown in Table 2. Higher rates of E. coli also were recorded by Peer et al.19 and Ammar et al.20. While, lower rates were recorded by Literak et al.21 and Radwan et al.22. Young aged broiler chickens up to 3 weeks have high affinity to the disease, but older chickens are more resistant. Different predisposing factors may increase the affinity of chickens to colibacillosis, such as viruses affecting the respiratory tract of chickens and bad hygienic mesearues23.
In this study, the recovered E. coli strains were typed serologically into 11 different ‘O’ groups; O78 (11.66%), O111: H4 (8.33%), O26: H11 (6.67%), O1: H7, O55: H7, 127: H6 and O128: H2 (5% for each), O2: H6, O44: H18, O119: H4 and O125: H21 (3.33% for each) and 24 E. coli strains were Untypable (40%). High percentage of Untypable strains in APEC was previously recorded by numerous studies in different countries24,25. This supports the suggestion that serotyping is not recommended as a specific diagnostic tool for the identification of avian pathogenic E. coli 26 and highlights the need for molecular characterization of E. coli virulence factors as several authors fail to differentiate between APEC and avian fecal E. coli by serogrouping only26,27. Untypable strains by O serogrouping are mainly arise due to autoagglutination and an incomplete antisera panel28.
The results of congo red test revealed that (91.7%) (n = 55 strains) of the isolated E. coli strains were positive and only (8.3%) (n = 5) were negative. These results agreed with those obtained by Sharda et al.29 who also reported a clear relationship between the expression of congo red and the pathogenicity in avian E. coli and stated that it was due to presence of β–D-glucan in bacterial cell wall. Yoder30 has reported that Congo red binding did not correlate well with pathogenicity.
The negativity of all the isolates to hemolysis on 5% sheep blood agar is in accordance with Erganiş et al.31 who attributed heavy mortality in chicks due to non-hemolytic strains indicating that avian pathogenic E. coli to be independent of hemolytic activity, Sharada et al.32 who reported that avian E. coli to be pathogenic needeed not to be hemolytic and Rodriguez et al.33 who reported that none of their isolates from colisepticemic cases was positive for hemolysis on 5% sheep blood agar.
In the present study as shown in Table 3 and Fig. 1-4; 36 serotyped E. coli strains were analyzed by PCR for detection of stx1, stx2, hly and eaeA genes. Regarding to the occurrence of stx1 gene, all E. coli strains were positive for stx1 gene (100%) with specific size of 180 bp, as shown in Fig. 1. These results were similar to those obtained by Momtaz and Jamshidi34 who showed that 100% of EHEC serogroups were positive for stx1.
Regarding to the presence of stx2 gene, 17 E. coli strains (8 EPEC strains, 4 ETEC strains and 5 EHEC strains) were positive for stx2 gene by percentage of (47.2%) and with specific size of 779 bp as shown in Fig. 2. These result were nearly similar to the findings which were recorded by El-Jakee et al.35 who detected stx2 in 5 E. coli strains (41.67%). The detection of STEC in chicken disagree with the results obtained by Kobayashi et al.36, Schroeder et al.37 and Wani et al.38.
Concerning PCR detection of Intimin gene (eaeA), as shown in Fig. 3, 12 E. coli strains (33.3%) (6 EPEC strains, 2 ETEC and 4 EHEC strains) were positive for intimin gene with specific size of 248 bp as shown in Fig. 3. These findings were nearly agreed with those obtained by El-Jakee et al.35 who detected eaeA in 5 E. coli strains (41.67%).
Regarding PCR amplification of hly gene, three non hemolytic EHEC strains (8.3%) were positive for hly gene with specific size of 1177 bp as shown in Fig. 4. These results agreed with the findings of Farah et al.39 who detected hlyA gene in seven non hemolytic strains. The absence of a hemolytic phenotype in the presence of hly gene has been identified and mainly attributed to defects in the hly genes or defects in the transcriptional activator rfaH40.
CONCLUSION
The combination of genotypic and phenotypic analysis of E. coli is more valuable as an epidemiological tool for identification of isolates. The PCR is a rapid and a specific diagnostic tool used for genetic characterization of Avian pathogenic E. coli. Escherichia coli strains originating from birds are carrying a low percentage of eaeA and hly genes. This study established the presence of stx1 and stx2 containing E. coli in chickens. Chickens might serve as vectors for transmission of STEC to environment and human.
REFERENCES
- Samah Eid, A.S. and A.M. Erfan, 2013. Characterization of E. coli associated with high mortality in poultry flocks. Assiut Vet. Med. J., 59: 51-61.
Direct Link - Tivendale, K.A., C.M. Logue, S. Kariyawasam, D. Jordan and A. Hussein et al., 2010. Avian-pathogenic Escherichia coli strains are similar to neonatal meningitis E. coli strains and are able to cause meningitis in the rat model of human disease. Infect. Immunity, 78: 3412-3419.
CrossRefDirect Link - Xia, X., J. Meng, P.F. McDermott, S. Ayers and K. Blickenstaff et al., 2010. Presence and characterization of Shiga toxin-producing Escherichia coli and other potentially diarrheagenic E. coli strains in retail meats. Applied Environ. Microbiol., 76: 1709-1717.
CrossRefDirect Link - Abong'o, B.O. and M.N. Momba, 2009. Prevalence and characterization of Escherichia coli O157:H7 isolates from meat and meat products sold in Amathole District, Eastern cape province of South Africa. Food Microbiol., 26: 173-176.
CrossRefDirect Link - Karch, H., P.I. Tarr and M. Bielaszewska, 2005. Enterohaemorrhagic Escherichia coli in human medicine. Int. J. Med. Microbiol., 295: 405-418.
CrossRefDirect Link - Caprioli, A., S. Morabito, H. Brugere and E. Oswald, 2005. Enterohaemorrhagic Escherichia coli: Emerging issues on virulence and modes of transmission. Vet. Res., 36: 289-311.
Direct Link - Piva, I.C., A.L. Pereira, L.R. Ferraz, R.S.N. Silva and A.C. Vieira et al., 2003. Virulence markers of enteroaggregative Escherichia coli isolated from children and adults with diarrhea in Brasilia, Brazil. J. Clin. Microbiol., 41: 1827-1832.
CrossRefDirect Link - Davies, R.H. and C. Wray, 1997. Immunomagnetic separation for enhanced flagellar antigen phase inversion in salmonella. Lett. Applied Microbiol., 24: 217-220.
CrossRefDirect Link - Berkhoff, H.A. and A.C. Vinal, 1986. Congo red medium to distinguish between invasive and non-invasive Escherichia coli pathogenic for poultry. Avian Dis., 30: 117-121.
PubMedDirect Link - Livezey, B.C. and R.L. Zusi, 2007. Higher-order phylogeny of modern birds (Theropoda, Aves: Neornithes) based on comparative anatomy. II. Analysis and discussion. Zool. J. Linnean Soc., 149: 1-95.
CrossRefDirect Link - Sambrook, J. and D.W. Russell, 2001. Molecular Cloning: A Laboratory Manual. 3rd Edn., Cold Spring Harbor Laboratory Press, New York, USA., ISBN-13: 9780879695774, Pages: 2344.
Direct Link - Sahilah, A.M., L.Y.Y. Audrey, S.L. Ong, W.N. Wan Sakeenah and S. Safiyyah et al., 2010. DNA profiling among egg and beef meat isolates of Escherichia coli by Enterobacterial Repetitive Intergenic Consensus-PCR (ERIC-PCR) and Random Amplified Polymorphic DNA-PCR (RAPD-PCR). Int. Food Res. J., 17: 853-866.
Direct Link - Dipineto, L., A. Santaniello, M. Fontanella, K. Lagos, A. Fioretti and L.F. Menna, 2006. Presence of Shiga toxin-producing Escherichia coli O157:H7 in living layer hens. Lett. Applied Microbiol., 43: 293-295.
CrossRefPubMedDirect Link - Bisi-Johnson, M.A., C.L. Obi, S.D. Vasaikar, K.A. Baba and T. Hattori, 2011. Molecular basis of virulence in clinical isolates of Escherichia coli and Salmonella species from a tertiary hospital in the Eastern Cape, South Africa. Gut Pathog, Vol. 3.
CrossRefDirect Link - Abouelmaatti, R.R., A.M. Algammal, X. Li, J. Ma, E.A. Abdelnaby and W.M.K. Elfeil, 2013. Experimental immunology cloning and analysis of Nile tilapia Toll-like receptors type-3 mRNA. Cent. Eur. J. Immunol., 38: 277-282.
CrossRefDirect Link - Peer, F.U., M.M. Ansari, I.A. Gani and M.M. Willayat, 2013. Serotyping and antibiotic sensitivity patterns of Escherchia coli isolates obtained from broiler chicks in Kashmir Valley, India. Adv. Anim. Vet. Sci., 1: 75-76.
Direct Link - Literak, I., T. Reitschmied, D. Bujnakova, M. Dolejska and A. Cizeket al., 2013. Broilers as a source of quinolone-resistant and extraintestinal pathogenic Escherichia coli in the Czech Republic. Microb. Drug Resistance, 19: 57-63.
CrossRefDirect Link - Radwan, I.A.E.H., H.S.H. Salam, S.A.E.A. Abd-Alwanis and M.A.Y. Al-Sayed, 2014. Frequency of some virulence associated genes among multidrug-resistant Escherichia coli isolated from septicemic broiler chicken. Int. J. Adv. Res., 2: 867-874.
Direct Link - Ashraf, A.A.E.T., A.M. Ammar, A.R. Ali, F.I.E. Hofy and M.E.E.S Ahmed, 2013. Detection of common (inv A) gene in salmonellae isolated from poultry using polymerase chain rection technique. Benha Vet. Med. J., 25: 70-77.
Direct Link - Hussein, A.H.M., I.A.I. Ghanem, A.A.M. Eid, M.A. Ali and J.S. Sherwood et al., 2013. Molecular and phenotypic characterization of Escherichia coli isolated from broiler chicken flocks in Egypt. Avian Dis., 57: 602-611.
CrossRefDirect Link - Ewers, C., E.M. Antao, I. Diehl, H.C. Philipp and L.H. Wieler, 2009. Intestine and environment of the chicken as reservoirs for extraintestinal pathogenic Escherichia coli strains with zoonotic potential. Appl. Environ. Microbiol., 75: 184-192.
CrossRefPubMedDirect Link - Oh, J.Y., M.S. Kang, H. Yoon, H.W. Choi and B.K. An et al., 2012. The embryo lethality of Escherichia coli isolates and its relationship to the presence of virulence-associated genes. Poult. Sci., 91: 370-375.
CrossRefDirect Link - Ozaki, H. and T. Murase, 2009. Multiple routes of entry for Escherichia coli causing colibacillosis in commercial layer chickens. J. Vet. Med. Sci., 71: 1685-1689.
CrossRefDirect Link - Sharda, R., S.W. Ruban and M. Thiyageeswaran, 2010. Isolation, characterization and antibiotic resistance pattern of Escherichia coli isolated from poultry. Am.-Eurasian J. Scient. Res., 5: 18-22.
Direct Link - Yoder, Jr. H.W., 1989. Congo red binding by Escherichia coli isolates from chickens. Avian Dis., 33: 502-505.
CrossRefDirect Link - Erganis, O., O. Kaya, M. Corlu and E. Istanbulluoglu, 1989. Hemagglutination, hydrophobicity, enterotoxigenicity and drug-resistance characteristics of avian Escherichia coli. Avian Dis., 33: 631-635.
CrossRefDirect Link - Rodriguez, M.F., G.D. Wiens, M.K. Purcell and Y. Palti, 2005. Characterization of Toll-like receptor 3 gene in rainbow trout (Oncorhynchus mykiss). Immunogenetics, 57: 510-519.
CrossRefDirect Link - Momtaz, H. and A. Jamshidi, 2013. Shiga toxin-producing Escherichia coli isolated from chicken meat in Iran: Serogroups, virulence factors and antimicrobial resistance properties. Poult. Sci., 92: 1305-1313.
CrossRefPubMedDirect Link - El-Jakee, J.K., R.M. Mahmoud, A.A. Samy, M.A. El-Shabrawy, M.M. Effat and W.A. Gad El-Said, 2012. Molecular characterization of E. coli isolated from chicken, cattle and buffaloes. Int. J. Microbiol. Res., 3: 64-74.
Direct Link - Kobayashi, H., T. Pohjanvirta and S. Pelkonen, 2002. Prevalence and characteristics of intimin- and Shiga toxin-producing Escherichia coli from gulls, pigeons and broilers in Finland. J. Vet. Med. Sci., 64: 1071-1073.
CrossRefPubMedDirect Link - Schroeder, C.M., D.G. White, B. Ge, Y. Zhang and P.F. McDermott et al., 2003. Isolation of antimicrobial-resistant Escherichia coli from retail meats purchased in Greater Washington, DC, USA. Int. J. Food Microbiol., 85: 197-202.
CrossRefDirect Link - Wani, S.A., I. Samanta, M.A. Bhat and Y. Nishikawa, 2004. Investigation of shiga toxin-producing Escherichia coli in avian species in India. Lett. Applied Microbiol., 39: 389-394.
CrossRefDirect Link - Farah, S.M.S.S., E.M. De Souza, F.O. Pedrosa, K. Irino and L.R. Da Silva et al., 2007. Phenotypic and genotypic traits of Shiga toxin-producing Escherichia coli strains isolated from beef cattle from Parana State, Southern Brazil. Lett. Applied Microbiol., 44: 607-612.
CrossRefDirect Link - Bailey, M.J.A., V. Koronakis, T. Schmoll and C. Hughes, 1992. Escherichia coli HIyT protein, a transcriptional activator of haemolysin synthesis and secretion, is encoded by the rfaH (sfrB) locus required for expression of sex factor and lipopolysaccharide genes. Mol. Microbiol., 6: 1003-1012.
CrossRefDirect Link