
Y. Okada
Deprtment of Basic Veterinary Medicine, School of Veterinary Medicine, Nippon Veterinary and Life Science University, 1-7-1 Kyonancho, Musashino, 180-8602 Tokyo, Japan
K. Kawasumi
Deprtment of Basic Veterinary Medicine, School of Veterinary Medicine, Nippon Veterinary and Life Science University, 1-7-1 Kyonancho, Musashino, 180-8602 Tokyo, Japan
M. Koide
Japan Horseback Riding Club, 685-1 Kitanagai Miyoshicho, Iruma-gun, 354-0044 Saitama, Japan
Y. Hirakawa
Deprtment of Basic Veterinary Medicine, School of Veterinary Medicine, Nippon Veterinary and Life Science University, 1-7-1 Kyonancho, Musashino, 180-8602 Tokyo, Japan
N. Mori
Deprtment of Basic Veterinary Medicine, School of Veterinary Medicine, Nippon Veterinary and Life Science University, 1-7-1 Kyonancho, Musashino, 180-8602 Tokyo, Japan
I. Yamamoto
Deprtment of Basic Veterinary Medicine, School of Veterinary Medicine, Nippon Veterinary and Life Science University, 1-7-1 Kyonancho, Musashino, 180-8602 Tokyo, Japan
T. Arai
Deprtment of Basic Veterinary Medicine, School of Veterinary Medicine, Nippon Veterinary and Life Science University, 1-7-1 Kyonancho, Musashino, 180-8602 Tokyo, Japan
Asian Journal of Animal and Veterinary Advances
Year: 2016 | Volume: 11 | Issue: 4 | Page No.: 253-257







ABSTRACT
Background: Aging is generally associated with alterations in physical activity, weight status and energy metabolism, which predisposes aged individuals to metabolic syndrome. In this manuscript, age effects on energy metabolic indicators of similar physical activity and weight status but of varying ages were investigated. Materials and Methods: Energy metabolic indicators, such as plasma adiponectin, leukocytic AMP-activated protein kinase, plasma malate dehydrogenase and lactate dehydrogenase along with common plasma metabolites, were measured in healthy young (AV = 7.1 years) and aged (AV = 14.1 years) riding horses of similar physical activity, diet and weight status. Malate dehydrogenase and lactate dehydrogenase ratio was also calculated as the indicator of energy metabolism. Results: Plasma adiponectin concentration and leukocytic AMP-activated protein kinase activity in aged horses were significantly lower than those in young horses (p<0.05, Mann-Whitney U test). Although not significant, energy metabolism indicators, malate dehydrogenase, lactate dehydrogenase and their ratio were lower in aged group when compared to those of young group. Conclusion: The present results indicate the decline in energy metabolism with aging in healthy horses even without any visible changes in adiposity. Such changes reflect dysfunction of energy metabolism and predispose the aged individuals to the development of metabolic syndrome.
PDF Abstract XML References Citation
How to cite this article
Y. Okada, K. Kawasumi, M. Koide, Y. Hirakawa, N. Mori, I. Yamamoto and T. Arai, 2016. Changes in Energy Metabolic Indicators with Aging in Thoroughbred Riding Horses. Asian Journal of Animal and Veterinary Advances, 11: 253-257.
DOI: 10.3923/ajava.2016.253.257
URL: https://scialert.net/abstract/?doi=ajava.2016.253.257
DOI: 10.3923/ajava.2016.253.257
URL: https://scialert.net/abstract/?doi=ajava.2016.253.257
INTRODUCTION
Energy metabolism and body fat distribution (proportion) generally alter with aging in animals. Glucose and lipids are the main sources of energy in mammals and their homeostasis is influenced by altered body weight and adipose tissue status. Previously, study showed down regulation of lipid and glucose metabolism in overweight animals with aging1. Since, the identification of the hormone, leptin2, adipose tissue has become recognized as not only a mere inert fuel storage but also an important secretory and endocrine organ that actively releases substances, adipokines involved in a wide array of physiological processes including glucose and lipid homeostasis, blood pressure, body weight regulations and immune functions3,4. As suggested by many studies, the dysregulated synthesis of harmful adipokines, such as tumor necrosis factor-alpha (TNFα)5, interleukin-6 (IL-6) and of beneficial adipokines, such as adiponectin (ADN)6 and leptin7, are involved in the development of metabolic syndrome, which is frequently seen in obese and/or aged individuals.
Aging is associated with decreased physical activity, one of the main causes of obesity. Riding horses are generally maintained at higher physical activity compared to other domestic animals. In this study, common plasma metabolites and energy metabolism indicators in riding horses of similar physical activity and weight status but of varying ages, were measured to elucidate the relationship among age, adipose statusand energy metabolism in horses.
MATERIALS AND METHODS
Animals: Subjects utilized in this study were twenty clinically normal thoroughbred riding horses (female n = 3, male n = 17), maintained at Japan Horseback Riding Club (Saitama, Japan). All males were gelded. They were divided into two groups based on age: young group (3-8 years old, average 7.1±0.5) and aged group (11-18 years old, average 14.1±0.7). The weight status of each subject was determined by the modified8 Carrol and Huntington method9 of six-scale Equine Body Condition Score (EBCS) and classified as follows: (1) Very thin, (2) Thin, (3) Fair, (4) Good, (5) Fat and (6) Very fat. Each horse was maintained on a diet consisting of: 5.2- 6.4 kg of hay cube, 3.0-4.0 kg of Italian ryegrass, 0-1.3 kg of wheat bran, 0-1.8 kg of barley, fed twice daily at 6:00 and 16:00. Exercise regimen of each horse consisted of: walking at 100-110 m min–1 for 10-30 min, trotting at 200-220 m min–1 for 10-30 min and cantering at 300-350 m min–1 for 15 min, 1-3 times daily for 6 days per week. The range in the exercise amount was due to the varying skills of the riding students. Ethical approval for this study was obtained from Nippon Veterinary and Life Science University Animal Research Committee.
Blood sampling and analysis: Blood sample from each horse was collected from the jugular vein into heparinized tubes. Plasma was recovered by centrifugation at 1200 g, for 5 min, 4°C and stored at -80°C until use. Buffy coat was stored in STE buffer at-80°C until use. Cytosolic fractions of leukocytes were prepared and isolated via ultrasonification, then centrifuged at 12000 g for 30 min at 4°C, a method previously described10.
Glucose (GLU), Total Cholesterol (TC), triglyceride (TG) concentrations and alanine aminotransferase (ALT), aspartate aminotransferase (AST), alkaline phosphatase (ALP) activities were measured using an auto-analyzer (JCA-BM2250, JEOL Ltd., Tokyo, Japan) with the manufacture’s reagents at Monolis Inc. (Tokyo, Japan). Activities of plasma lactate dehydrogenase (LDH) with pyruvate as substate11 and malate dehydrogenase (MDH)12 were calculated by conversion of NADH to NAD+, interpreted by the changes in spectrophotometric absorption at 340 nm. Plasma MDH/LDH ratio was calculated by the division of MDH activity by LDH activity. Plasma non-esterified fatty acid (NEFA) was measured using a commercial kit, NEFA-C test (Wako Pure Chemical Industries, Inc., Tokyo, Japan). Plasma Insulin (INS) and adiponectin (ADN) concentrations were measured with commercial ELISA kits, Lbis Rat T insulin kit (Shibayagi Co., Gunma, Japan), mouse/rat adiponectin ELISA kit (Osuka Pharmaceutical Co., Ltd., Tokyo, Japan), respectively. Leukocytic AMP-activated protein kinase (AMPK) activities were measured using a commercial ELISA kit, CycLex AMPK Kinase Assay kit (CycLex Co., Ltd. na, Nagano, Japan). The AMPK activity was measured at 30°C for 30 min and is expressed as nanogram of phosphorylated substrate per minute per mg of protein (specific activity). Protein concentration in cytosol fraction was measured with the Bradford method13 using bovine serum albumin as the standard. The enzyme unit (IU) in IU per liter of plasma (volume activity) represents 1 μmol of substrate degraded per min.
Statistical analysis: All values are expressed as Mean±SE. Statistical significance was analyzed by Mann-Whitney U test. The significance level was set at p<0.05.
RESULTS
The body weight of study subjects ranged from 450-500 kg. The weight status of all the horses in this study was determined as EBCS = 4, based on the modified Carroll and Huntington EBCS scoring system8,9.
| Table 1: | Comarison of the biochemical and energy metabolism parameters between the two age groups of riding horse |
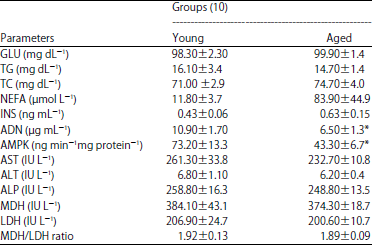 | |
Values are presented Mean±SE, *Significantly different (p<0.05) when compared against young group, (p<0.05, Mann-Whitney U test) GLU: Glucose, TG: Triglyceride, TC: Total cholestrol, NEFA: Non-esterified fatty acid, INS: Insulin, ADN: Adiponectin, AMPK: AMP-activated protein kinase, AST: Aspartate aminotransferase, ALT: Alanine aminotransterase, ALP: Alkalinc phosphatase, MDH: Malate dehydrogenase and LDH: Lactate dehydrogenase | |
As shown in Table 1, plasma ADN concentration and leukocyte cytosolic AMPK activity level in aged horses were significantly lower compared to those in young horses (6.5±1.3 vs. 10.9±1.7 μg mL–1; 43.4±6.7 vs. 73.3±13.3 ng min–1 mg protein–1, respectively). Although not significant, plasma NEFA and INS concentrations in aged group were higher than those in young group (83.9±44.9 vs. 11.8±3.7 μmol mL¯1 and 0.63±0.15 vs. 0.43±0.06 ng mL–1, respectively). Although not significant, the activity levels of MDH, LDH and M/L ratio (1.89±0.09 vs. 1.92±0.13) of aged group were lower compared to those in young group. No difference was noted in the commonly measured plasma biochemical parameters GLU, TG, TC, AST, ALT, ALP between the two groups.
DISCUSSION
Aging and obesity share similar clinical consequences such as diabetes, hypertension, neoplasia, orthopedic and cardiac diseases14,15. Many evidences attribute abnormal adipose tissue (excessive or dysfunctional) and dysregulated synthesis of adipokines to the development of age-related diseases and reduced life span16,17. It is further supported by the fact that limiting energy intake (dietary restriction) leads to extended lifespan8.
In this study, the subjects in two groups all had EBCS of 4.0, which is considered good according to the 6.0-scale evaluation system, thus were not considered overweight. Furthermore, most plasma biochemical parameters, commonly used to assess overall health status of individuals, showed no significant difference between the two groups. However, significantly lower ADN concentration and AMPK activity were shown in aged group, as well as decreasing tendencies of MDH, LDH activities and M/L ratio, all of which are considered to reflect changes in glucose lipid metabolism, energy stateand the metabolic health of individuals. The present study results are in concordance with the previous studies. Changes in energy metabolism are indeed part of the normal aging process in many species18.
The ADN is a cytokine secreted by adipocytes. It is involved in glucose and lipid metabolism and exerts direct effects on vasculatures, thus positively influences the metabolic health of individuals19. It modulates the expression of fatty acid transport proteins in liver and skeletal muscle and stimulates fatty acid oxidation and glucose utilization via activation of AMPK20. It also exerts anti-inflammatory actions via inhibition of macrophage activity and C-reactive protein, TNFα production and action. It is considered beneficial adipokine and its decline in plasma is associated with disease processes such as obesity, insulin resistant Type 2 diabetes, coronary artery disease in humans5,20, which commonly develop in aged individuals. Plasma ADN concentration was also shown to decline with aging in clinically healthy dogs21. Unlike other adiopokines, secretion and circulating levels of ADN are inversely proportional to body fat mass22. Many evidences further suggest the significance of adipose status, such as adipose depot distribution (subcutaneous vs. visceral) and adipocyte size (small vs. large) in ADN synthesis and secretion6,23.
AMP-activated protein kinase is a downstream component of protein kinase cascade activated by increasing AMP coupled with ATP depletion. It is a sensor of cellular energy status and mediates effects of adipokines in food intake, body weightand glucose and lipid metabolism24, reducing the risk of obesity and insulin resistance24,25. Additionally, AMPK activation suppresses oxidative stress and inflammatory processes, which are the characteristics commonly seen in older individuals. Previous studies have shown a crucial role of AMPK in regulating aging speed and lifespan in several species19,26,27,28 and its decline in activity with aging.
The MDH, a rate-limiting enzyme of the malate-aspartate shuttle, plays a crucial role in ATP production as it is involved in transportation of cytosolic NADH, generated by glycolysis, into mitochondria29. Its activity level and M/L ratio determined by the division of MDH by lactate dehydrogenase (LDH), a cytosolic marker involved in reversible conversion of pyruvate to lactate, are considered to reflect energy metabolism changes in various individuals and health conditions, such as race horses under intense training30, canine lymphoma31 and diabetes mellitus32 and acute weight gain in dogs33. The present results may also suggest its potential diagnostic significance in assessing the effects of aging in horses.
The results in this study showed shifts in energy metabolism indicators with aging in horses. These changes reflect dysfunction of energy metabolism and predispose the aged individuals to the development of metabolic syndrome. The energy metabolism indicators such as ADN, AMPK and M/L ratio may be useful for the early detection of latent metabolic disorders in aging animals.
The effects of aging on shifts in fat cell size and distribution (adipose status) and energy metabolism, were not directly assessed in this study but are strongly suggested since weight and EBCS were not different between the two age groups. Further studies assessing the differences not only in mere adiposity but also in adipose status (fat cell size and distribution) against age and energy metabolism indicators are warranted in order to elucidate the physical and biochemical effects of aging in horses.
CONCLUSION
The energy metabolism indicators such as adiponectin, AMP-activated protein kinase, malate dehydrogenase and lactate dehydrogenase ratio may be useful for the early detection of latent metabolic disorders in aging animals. Further studies assessing the differences not only in mere adiposity but also in adipose status (fat cell size and distribution) against age and energy metabolism indicators are warranted in order to elucidate the physical and biochemical effects of aging in horses.
ACKNOWLEDGMENTS
The authors would like to acknowledge Dr. T. Nuruki of YCL Horse Clinic, for sample collection and Dr. A. Usui of Nippon Veterinary and Life Science University Equestrian Club Head Coach, for assessing physical status of the study subjects.
REFERENCES
- Kawasumi, K., N. Kashiwado, Y. Okada, M. Sawamura and Y. Sasaki et al., 2014. Age effects on plasma cholesterol and triglyceride profiles and metabolite concentrations in dogs. BMC Vet. Res., Vol. 10.
CrossRef - Zhang, Y., R. Proenca, M. Maffei, M. Barone, L. Leopold and J.M. Friedman, 1994. Positional cloning of the mouse obese gene and its human homologue. Nature, 372: 425-432.
PubMedDirect Link - Trayhurn, P. and I.S. Wood, 2005. Signalling role of adipose tissue: Adipokines and inflammation in obesity. Biochem. Soc. Trans., 33: 1078-1081.
CrossRefDirect Link - Trayhurn, P., C. Bing and I.S. Wood, 2006. Adipose tissue and adipokines-energy regulation from the human perspective. J. Nutr., 136: 1935S-1939S.
Direct Link - Ouchi, N., S. Kihara, Y. Arita, K. Maeda and H. Kuriyama et al., 1999. Novel modulator for endothelial adhesion molecules: Adipocyte-derived plasma protein adiponectin. Circulation, 100: 2473-2476.
CrossRefPubMedDirect Link - Kantartzis, K., K. Rittig, B. Balletshofer, J. Machann and F. Schick et al., 2006. The relationships of plasma adiponectin with a favorable lipid profile, decreased inflammation and less ectopic fat accumulation depend on adiposity. Clin. Chem., 52: 1934-1942.
CrossRefDirect Link - Kershaw, E.E. and J.S. Flier, 2004. Adipose tissue as an endocrine organ. J. Clin. Endocrinol. Metab., 89: 2548-2556.
Direct Link - Roth, G.S., M.A. Lane, D.K. Ingram, J.A. Mattison and D. Elahi et al., 2002. Biomarkers of caloric restriction may predict longevity in humans. Science, 297: 811-811.
CrossRefDirect Link - Carroll, C.L. and P.J. Huntington, 1988. Body condition scoring and weight estimation of horses. Equine Vet. J., 20: 41-45.
CrossRefDirect Link - Washizu, T., E. Kuramoto, M. Abe, T. Sako and T. Arai, 1998. A comparison of the activities of certain enzymes related to energy metabolism in leukocytes in dogs and cats. Vet. Res. Commun., 22: 187-192.
CrossRefDirect Link - Kaloustian, H.D., F.E. Stolzenbach, J. Everse and N.O. Kaplan, 1969. Lactate dehydrogenase of lobster (Homarus americanus) tail muscle I. Physical and chemical properties. J. Biol. Chem., 244: 2891-2901.
Direct Link - Bradford, M.M., 1976. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem., 72: 248-254.
CrossRefPubMedDirect Link - German, A.J., V.H. Ryan, A.C. German, I.S. Wood and P. Trayhurn, 2010. Obesity, its associated disorders and the role of inflammatory adipokines in companion animals. Vet. J., 185: 4-9.
CrossRefPubMedDirect Link - Barzilai, N., L. She, B.Q. Liu, P. Vuguin, P. Cohen, J. Wang and L. Rossetti, 1999. Surgical removal of visceral fat reverses hepatic insulin resistance. Diabetes, 48: 94-98.
CrossRefDirect Link - Muzumdar, R., D.B. Allison, D.M. Huffman, X. Ma and G. Atzmon et al., 2008. Visceral adipose tissue modulates mammalian longevity. Aging Cell, 7: 438-440.
CrossRefDirect Link - Apfeld, J., G. O'Connor, T. McDonagh, P.S. DiStefano and R. Curtis, 2004. The AMP-activated protein kinase AAK-2 links energy levels and insulin-like signals to lifespan in C. elegans. Genes Dev., 18: 3004-3009.
CrossRefPubMedDirect Link - Laflamme, D.P., 2012. Companion animals symposium: Obesity in dogs and cats: What is wrong with being fat? J. Anim. Sci., 90: 1653-1662.
CrossRefPubMedDirect Link - Chandran, M., T. Ciaraldi, S.A. Phillips and R.R. Henry, 2003. Adiponectin: More than just another fat cell hormone? Diabetes Care, 26: 2442-2450.
CrossRefPubMedDirect Link - Mori, N., K. Kawasumi and T. Arai, 2012. Comparison of the plasma insulin and adiponectin concentrations as metabolic markers in clinically healthy dogs with ageing. J. Anim. Vet. Adv., 11: 971-974.
CrossRefDirect Link - Arita, Y., S. Kihara, N. Ouchi, M. Takahashi and K. Maeda et al., 1999. Paradoxical decrease of an adipose-specific protein, adiponectin, in obesity. Biochem. Biophys. Res. Commun., 257: 79-83.
CrossRefPubMedDirect Link - Meyer, L.K., T.P. Ciaraldi, R.R. Henry, A.C. Wittgrove and S.A. Phillips, 2013. Adipose tissue depot and cell size dependency of adiponectin synthesis and secretion in human obesity. Adipocyte, 2: 217-226.
CrossRefDirect Link - Kahn, B.B., T. Alquier, D. Carling and D.G. Hardie, 2005. AMP-activated protein kinase: Ancient energy gauge provides clues to modern understanding of metabolism. Cell Metab., 1: 15-25.
CrossRefDirect Link - Balsan, G.A., J.L.C. Vieira, A.M. de Oliveira and V.L. Portal, 2015. Relationship between adiponectin, obesity and insulin resistance. Revista Associacao Medica Brasileira, 61: 72-80.
CrossRefDirect Link - Viollet, B., F. Andreelli, S.B. Jorgensen, C. Perrin and D. Flamez et al., 2003. Physiological role of AMP-activated protein kinase (AMPK): Insights from knockout mouse models. Biochem. Soc. Trans., 31: 216-219.
PubMedDirect Link - Funakoshi, M., M. Tsuda, K. Muramatsu, H. Hatsuda, S. Morishita and T. Aigaki, 2011. A gain-of-function screen identifies wdb and lkb1 as lifespan-extending genes in Drosophila. Biochem. Biophys. Res. Commun., 405: 667-672.
CrossRefDirect Link - Hwang, A.B., E.A. Ryu, M. Artan, H.W. Chang and M.H. Kabir et al., 2014. Feedback regulation via AMPK and HIF-1 mediates ROS-dependent longevity in Caenorhabditis elegans. Proc. Natl. Acad. Sci., 111: E4458-E4467.
CrossRefDirect Link - Setoyama, C., T. Joh, T. Tsuzuki and K. Shimada, 1988. Structural organization of the mouse cytosolic malate dehydrogenase gene: Comparison with that of the mouse mitochondrial malate dehydrogenase gene. J. Mol. Biol., 202: 355-364.
CrossRefDirect Link - Li, G., P. Lee, N. Mori, I. Yamamoto and T. Arai, 2012. Long term intensive exercise training leads to a higher plasma malate/lactate dehydrogenase (M/L) ratio and increased level of lipid mobilization in horses. Vet. Res. Commun., 36: 149-155.
CrossRefDirect Link - Washizu, T., D. Azakami, M. Bonkobara, M. Washizu and T. Arai, 2005. Changes in activities of enzymes related to energy metabolism in canine lymphoma cells. J. Vet. Med. Sci., 67: 615-616.
CrossRefDirect Link - Magori, E., M. Nakamura, A. Inoue, A. Tanaka and N. Sasaki et al., 2005. Malate dehydrogenase activities are lower in some types of peripheral leucocytes of dogs and cats with type 1 diabetes mellitus. Res. Vet. Sci., 78: 39-44.
CrossRefDirect Link - Okada, Y., K. Kawasumi, N. Mori, I. Yamamoto and T. Arai, 2015. Changes in malate dehydrogenase, lactate dehydrogenase and M/L ratio as energy metabolism markers of acute weight gain Asian J. Anim. Vet. Adv., 10: 132-140.
CrossRefDirect Link