
Faez Firdaus Jesse Abdullah
Department of Clinical Studies, Faculty of Veterinary Medicine, Universiti Putra Malaysia, 43400 UPM Serdang, Selangor, Malaysia
LiveDNA: 60.13767
Abdinasir Yusuf Osman
Department of Clinical Studies, Faculty of Veterinary Medicine, Universiti Putra Malaysia, 43400 UPM Serdang, Selangor, Malaysia
Lawan Adamu
Department of Clinical Studies, Faculty of Veterinary Medicine, Universiti Putra Malaysia, 43400 UPM Serdang, Selangor, Malaysia
Zunita Zakaria
Department of Pathology and Microbiology, Faculty of Veterinary Medicine, Universiti Putra Malaysia, 43400 UPM Serdang, Selangor, Malaysia
Rasedee Abdullah
Department of Clinical Studies, Faculty of Veterinary Medicine, Universiti Putra Malaysia, 43400 UPM Serdang, Selangor, Malaysia
Mohd Zamri-Saad
Research Centre for Ruminant Disease, Universiti Putra Malaysia, 43400 UPM Serdang, Selangor, Malaysia
Abdul Aziz Saharee
Department of Clinical Studies, Faculty of Veterinary Medicine, Universiti Putra Malaysia, 43400 UPM Serdang, Selangor, Malaysia
Asian Journal of Animal and Veterinary Advances
Year: 2013 | Volume: 8 | Issue: 6 | Page No.: 806-813







ABSTRACT
Pasteurella multocida is an opportunistic pathogen with veterinary and human significance. The species diverse and complexity between the types exist with respect to antigenic variation, host predilection and pathogenesis. The aim of the present study was to determine hematological and serum concentration of biochemical values in calves having infected with Pasteurella multocida type B and its immunogens; Lipopolysaccharide (LPS) and Outer Membrane Protein (OMP). A total of eight clinically healthy calves were divided into four groups of 2 calves in each group. Group 1 were inoculated with sterile Phosphate Buffered Saline (PBS). Group 2 were inoculated with wild-type P. multocida type B: 2 while Group 3 and 4 were inoculated with LPS and OMP extracted from Pasteurella multocida type B, respectively. Blood samples were collected for haematological and biochemical analysis. Results indicated significant differences in the mean values of Prothrombin Time (PT), Partial Thromboplastin Time (PTT) and thrombocyte levels in infected groups 2-4 compared to the control group. There was significant decrease in the mean concentration of Alanine Transaminase (ALT) for group 2 (20.607±1.639 U L-1), group 3 (22.067±1.617 U L-1) and group 4 (40.327±8.351 U L-1). Conversely, there was no significant difference in Alkaline Phosphatase (ALP) for group 2 (149.857±17.327 U L-1) but a significant decrease for groups 3 (114.147±20.901 U L-1) and group 4 (128.971±17.068 U L-1). There was no significant differences in the serum levels of Gamma-glutamyltranspeptidase (GGT) and almost all tested electrolytes in infected groups. In conclusion, our results provide evidence in support of using some haematological and biochemical markers for early detection of individuals at risk for wild type of Pasteurlla multocida.
PDF Abstract XML References Citation
Received: May 06, 2013;
Accepted: August 17, 2013;
Published: September 25, 2013
How to cite this article
Faez Firdaus Jesse Abdullah, Abdinasir Yusuf Osman, Lawan Adamu, Zunita Zakaria, Rasedee Abdullah, Mohd Zamri-Saad and Abdul Aziz Saharee, 2013. Haematological and Biochemical Alterations in Calves Following Infection
with Pasteurella multocida Type B: 2, Bacterial Lipopolysaccharide and
Outer Membrane Protein Immunogens (OMP). Asian Journal of Animal and Veterinary Advances, 8: 806-813.
DOI: 10.3923/ajava.2013.806.813
URL: https://scialert.net/abstract/?doi=ajava.2013.806.813
DOI: 10.3923/ajava.2013.806.813
URL: https://scialert.net/abstract/?doi=ajava.2013.806.813
INTRODUCTION
Haemorrhagic Septicaemia (HS) caused by mainly Pasteurella multocida manifest often as respiratory infection with characteristic of per-acute signs in various species of animals. It is one of the most common diseases of cattle, goats and sheep in tropical countries and has a major of economic importance and welfare implications worldwide. Outbreaks of this disease usually relate to high mortality and great economic alterations to the ruminant industry (Gilmour et al., 1991; Brogden et al., 1998; Odugbo et al., 2003). This is evidenced by the global attention and the political debate of this disease particularly in temperate areas including Malaysia. The organism is important of primary or secondary animal pathogen for over a century and is becoming important as human pathogen leading to a disease entity termed as pasteurellosis. The organisms usually act as secondary invaders in animals with concurrent diseases or suffering from debilitating stressful conditions (Benirschke et al., 1978). The serotype B has been recovered from HS-affected animals in Southern Europe, Central and South America, the Middle East and Asia, including Malaysia. The infection causes substantial morbidity and mortality in cattle, buffaloes, sheep and goats (Bain et al., 1982). In Malaysia, the stressful condition is during the raining season where most outbreaks occurred (Saharee et al., 1993).
Although, in Malaysia Haemorrhagic Septicaemia (HS) does appear to receive the attention it deserves, the disease, however, is still threatening the large ruminant sector resulting economic losses in the production of milk, meat and the by-products such as hide for the leather industry. According to the National Immunization Plan (NIP), killed whole cell vaccines have been used in Malaysia to control the Haemorrhagic septicaemia. These vaccines have not been nationally licensed due to the defects that imply shorter duration of immunity and swelling at the site of inoculation. However, previous studies have hypothetically proposed that the capsular antigens; Lipopolysaccharide (LPS) or LPS-protein complex and the Outer Membrane Proteins (OMPs), including the iron-regulated OMPs might be useful as an effective immunogens for sero-groups B and E (Carter and de Alwis, 1989). However, the mechanisms of these immunogens remain largely obscure and to improve strategies for the overall understanding of these components, it is desirable to establish a base line data in relation to the blood picture of these immunogens in the actual host. Thus, the objective of this study was to determine the changes in the blood parameters of cattle following infection by live P. multocida B: 2 and inoculation with the bacterial OMP and LPS.
MATERIALS AND METHODS
Animals: Eight clinically healthy calves of approximately 12 months of age and weighing 150±50 kg were selected from a farm. Nasal swabs were collected immediately after selection and every 3 days for at least 2 weeks to ensure that the calves were free of P. multocida prior to the start of the experiment. The animals were then transported by road to the Animal Experimental Unit (AEU). Upon arrival at the AEU, anthelmintic (Ivermectin) was administered subcutaneously at the rate of 1 mL 50 kg-1 b.wt. to control internal parasitism which has been shown to influence disease development. The animals were then divided equally into 4 groups, kept in separate rooms and fed with cut grass and supplemented feed at the rate of 1 kg day-1.
Inoculum: A total of three types inocula were used throughout the experiments: (1) Wild-type P. multocida B: 2 isolated from a previous outbreak of HS in the state of Kelantan, Malaysia; (2) The Lipopolysaccharide (LPS) extracted from P. multocida B: 2 and (3) Outer Membrane Proteins (OMP) extracted from P. multocida type B (Table 1). The inocula (LPS and OMP) were extracted according to the method described by Abdullah et al. (2013). Isolates of P. multocida type B were collected from freeze-dried stock cultures and grown on blood agar for 24 h. McFarland method was used to determine the concentration of the inocula.
| Table 1: | Experimental design |
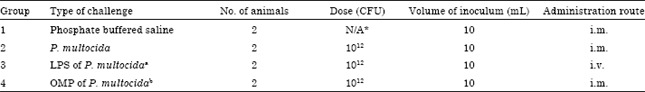 | |
| *N/A: Not applicable, aLipopolysaccharide and bOuter membrane protein | |
Experimental procedure: At the start of the experiment, the heifers were divided into 4 groups of 2 cows in each group. Cows of group 1 served as control group and were inoculated intramuscularly with 10 mL of sterile Phosphate Buffered Saline (PBS). Cows of group 2 were inoculated intramuscularly with 10 mL of 1012 Colony Forming Unit (CFU) of wild-type P. multocida. Cows of group 3 received intravenously the inoculum containing 10 mL of LPS broth extract. Cows of group 4 received intramuscularly the inoculum containing 10 mL of OMP broth extract (Table 1). Following intramuscular/intravenous challenge, all animals were observed for 48 h for signs of respiratory infection and mortality pattern before the surviving animals were euthanized after 48 h post-inoculation. Serum samples were collected from all challenged and non-challenged animals via jugular venipuncture at 3 h intervals. The entire respiratory tracts were monitored and the extent of the damage of the visceral organs with emphasis on lung lesions was determined according to the method described by Gilmour et al. (1990).
Blood analysis
Haematology: Hematocrit, total solids and white blood cell counts were conducted using the Cell-Dyn 3700 Automatic Analyzer (Vet Package, Abbot Diagnostic, 2007) conducted at the Hematology and Biochemistry Laboratory, Faculty of Veterinary Medicine, Universiti Putra Malaysia. Pack Cell Volume (PCV) was determined by micro-haematocrit technique. Blood smears were stained with Wright Stain (Sigma®) for differential counting of leucocytes which is the percentage of lymphocytes, monocytes, basophils, eosinophils and neutrophils. Activated Partial Thromboplastin Time (APTT) and Prothrombin Time (PT) were analyzed using ST Art 4® (Diagnostic Stago, France).
Serum biochemistry: Sera were processed by automated chemistry analyzer (HITACHI 902 Automatic Analyzer®, Japan) for biochemistry parameters. The analyzed parameters include sodium (Na), potassium (K), chloride (Cl), calcium (Ca), inorganic phosphate (P), Blood Urea Nitrogen (BUN), creatinine (Crea), glucose, Total Bilirubin (T. Bill), Total Protein (TP), Alanine Aminotransferase (ALT), Alkaline Phosphatase (ALP), Aspartate Aminotransferase (AST), Creatinine Kinase (CK), Albumin (Alb), Gamma-glutmyltransferase (GGT) and globulin and albumin ratio (A:G).
Statistical methods: The mean values obtained from haematological and biochemical assays of samples from infected animals with Pasteurella multocida type B and its immunogens; LPS and OMP were compared with data from the control group using the Tukey-test. Differences were considered to be statistically significant with values of p<0.05.
RESULTS
Blood and biochemistry: Haematologically, the characteristics of tested parameters in the different groups involved in the study are summarized in Table 2. There were significant differences in the mean values of Prothrombin Time (PT), Partial Thromboplastin Time (PTT) and thrombocyte levels in the challenged groups 2-4 compared to the control group (Table 1). Mean PT was significantly higher in group 2-4 with mean levels 25.300±4.623; 23.292±1.780; 22.250±1.512, respectively than the negative control group. For Partial Thromboplastin Time (PTT), only group 3 (30.327±1.543) showed a significant 1 time increased compared to the negative control (Table 2). Similarly, mean levels of thrombocyte were significantly decreased in group 2-4 (239.571±75.681; 296.029±79.48; 335.49±125.20, respectively) than control group with mean values of 404.50±96.763 (Table 2).
Among the challenged groups, the animals received the inoculum containing live Pasteurella multocida showed significant differences in PCV and leukocyte counts where this group showed no significant differences in RBC and Hemoglobin counts compared to control group. Both groups 3 and 4 inoculated with immunogens (LPS and OMP) did not show significant difference in leucocyte counts and white blood cells. There were relatively little effect of immunogens LPS and OMP broth extract compared to live P. multocida B: 2 on all investigated hematological parameters (Table 2).
There was no significant differences in the serum levels of Gamma-glytamyltranspeptidase (GGT) and almost all tested electrolytes; Na, Cl and K (Table 3). In contrast, all challenged groups showed significant decrease in the levels of calcium ion for group 2-4 with mean values of 1.851±0.319, 2.185±0.176 and 2.260±0.141 mmol L-1, respectively. Similarly, there was significant decrease in the mean concentration of Alanine Transaminase (ALT) for group 2 (20.607±1.639 U L-1), group 3 (22.067±1.617 U L-1) and group 4 (40.327±8.351 U L-1). Moreover, there was no significant difference in Alkaline Phosphatase (ALP) for group 2 (149.857±17.327 U L-1) but a significant decrease for groups 3 (114.147±20.901 U L-1) and group 4 (128.971±17.068 U L-1).
| Table 2: | Blood haematology changes in cattle inoculated with various immunogens of P. multocida B: 2 |
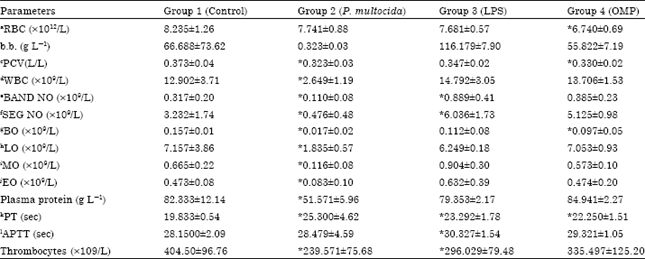 | |
| aRed blood cells, bHaemoglobin, cPacked cell volume, dWhite blood cells, eBanded neutrophils, fSegmented neutrophils, gBasophils, hLymphocytes, IMonocytes, jEosinophils, kProthrombin timem, lActivated partial thromboplastin time. *Significant value p<0.05. Comparison between immunogens groups and negative control group | |
| Table 3: | Blood serum biochemistry in calves inoculated with different immunogens of P. multocida B: 2 |
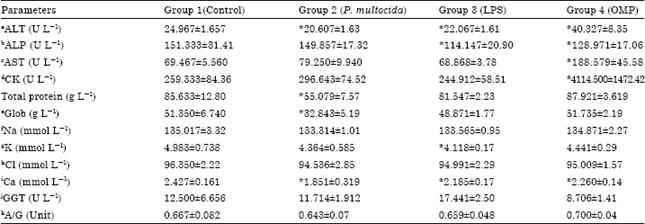 | |
| aAlanine transaminase, bAlkaline phosphatase, cAspartate transaminase, dCreatine kinase, eGlobulin, fSodium, gPotassium, hChloride, iCalcium, jGamma-glutamyl transpeptidase, kAlbumin/globulin ratio. *Significant value p<0.05. Comparison between immunogens groups and negative control group | |
In Aspartate Transaminase (AST), only group 4 (188.579±45.581 U L-1) showed significant increase in the mean concentration where groups 2-3 showed no differences with mean values of (79.250±9.940 U L-1) and (68.868±3.784 U L-1), respectively. In total protein levels, group 3 (81.547±2.235 g L-1) and 4 (87.921±3.619 g L-1) showed no significant difference but group 2 (55.079±7.578 g L-1) showed a significant increase in the level of total protein compared to the negative control (Table 3). Similarly, groups 3 (48.871±1.770 g L-1) and 4 (51.735±2.192 g L-1) showed no significant difference in the globulin level but group 2 (32.843±5.194 g L-1) showed a significant decrease in the level of globulin. There was no significant difference in Creatine Kinase (CK) level in groups 2 (296.643±74.524 U L-1) and 3 (244.912±58.513 U L-1) but a significant 16 fold increase in group 4 (4114.500±1472.42 U L-1) (Table 3).
DISCUSSION
Severe inflammatory diseases, regardless of the inciting agent, can lead to neutropenia if neutrophil margination and emigration into inflamed tissues exceeds the release of neutrophils from marrow. With small storage pool of segmented neutrophils in bovine marrow, it is common for cattle with acute bacterial infections to have neutropenia. However, within days, increased neutrophil production and release may result in neutrophilia (Steven, 2000). Todar (2008) stated that injection of living or killed Gram-negative cells or purified LPS into experimental animals causes a wide spectrum of non-specific pathophysiological reactions such as fever, changes in white blood cell counts, Disseminated Intravascular Coagulation (DIC), shock and death. Jacobsen et al. (2004) stated that after 30 min of LPS injection to the cows, it was observed that the host showed leukocytosis, thrombocytopenia, hypocalcemia, hypoferremia and hypozincemia followed by a short hyperzincemia. Smedegard et al. (1989) stated that effect of endotoxin from Gram-negative bacteria in rats lead to systemic hypotension, increased hematocrit and decreased number of circulating leukocytes (polymorphonuclear), monocytes and platelets. Brigham and Meyrick (1986) also mentioned that effects of LPS in experimental animals are systemic hypotension and pulmonary hypertension and cause a decreased in circulating leukocytes and platelets. Steven (2000) stated that eosinophilia is an occasional finding in bacterial infection and may due to inflammatory reactions. Calves inoculated with whole cells showed significant overall decrease (p<0.05) in white blood cell counts. This result was consistent with Steven (2000) and Todar (2008), who indicated that acute bacterial infection changes the white blood cell counts, mainly through neutropenia. However, calves inoculated with LPS showed no significant change in lymphocytes count. This result was not in agreement with Brigham and Meyrick (1986), Smedegard et al. (1989) and Jacobsen et al. (2004), who found that administration of LPS lead to decreased leukocytes. This may due to the fact that LPS was administered only once in this study instead of repeated administration (Rokke et al., 1988; Warner et al., 1988). For lymphocytes count, the positive control group of calves showed significant decreased (p<0.05) in lymphocyte number which may cause lysis of the bacteria that lead to formation of high concentration of endotoxin that can cause lysis of lymphocytes. In this study, the monocytes and eosinophil counts decreased significantly (p<0.05) in the positive control group calves as documented by Smedegard et al. (1989) who stated that endotoxin can caused reduction in monocyte number. The thrombocytes counts were reduced significantly (p<0.05) in the positive control and LPS group calves which can be associated with endotoxin that cause decreased in circulating platelets or thrombocytes (Brigham and Meyrick, 1986; Smedegard et al., 1989; Jacobsen et al., 2004). In this study, the prothrombin time increased significantly (p<0.05) in the positive control and OMP group calves indicating that in these two groups, the extrinsic pathway of coagulation had been activated. For Activated Partial Thromboplastin Time (APTT) only the LPS immunogen group calves showed significant increase (p<0.05) in time indicating that in the LPS immunogen group calves, the intrinsic pathway of coagulation had been activated. Moustafa et al. (1991) stated that during trypanosomiasis and pasteurellosis or stress the total serum protein and albumin levels will decrease and globulin level will increase. Kamal (2008) and Seleim et al. (2003) observed increased levels in creatinine and elevation of serum ALT and AST concentrations during pasteurellosis. Tietz (1994), Koneka et al. (1997) and Seleim et al. (2003) observed decrease in sodium and chloride levels and increase in potassium in septicaemic disease such as pasteurellosis. In this study, the positive control group calves showed significant decrease (p<0.05) in total serum protein and globulin levels. The reduction in the levels of total serum protein is an agreement with Moustafa et al. (1991) but the decrease in globulin level was not in agreement with Moustafa et al. (1991). All the immunogen group calves showed significant decrease (p<0.05) in calcium levels and this reduction in calcium levels may due to hypoalbuminemia. In this study, there are no significant changes in sodium, potassium and chloride levels in all the three immunogen groups and these results were not in agreement with Tietz (1994), Koneka et al. (1997) and Seleim et al. (2003). Morris et al. (2004) stated that Gamma Glutamyltransferase (GGT) is a specific liver enzyme in the serum of cattle with liver damage. Morris et al. (2004) mentioned that there is a significant relationship between severity of liver injury and serum GGT levels in ruminants. Ozmen et al. (2008) also stated that high serum GGT levels in sheep can be associated with severely affected livers. From this study, all immunogen treated group calves did not show any significant changes in GGT level. The positive control and LPS group calves showed significant decrease (p<0.05) in ALT levels whereas the OMP group calves showed significant increase (p<0.05) in the concentration ALT. From the results of this study, it can be concluded that all the immunogen group calves did not develop severe liver damage, as severe liver damage will cause the GGT to rise due to the fact that, GGT is a specific liver enzyme in ruminants (Morris et al., 2004; Ozmen et al., 2008). Bishop et al. (2004) mentioned that elevation of Creatinine Kinase (CK) can be related to muscle injury. From the results of this study, the OMP group calves showed significant increase (p<0.05) in CK level which can be associated with muscle damage caused by OMP immunogen. The Aspartate Aminotransferase(AST) level increased significantly (p<0.05) in the OMP group calves indicating muscle damage due to OMP whereas, AST level will increase in both muscle and liver damage (Bishop et al., 2004). Less iron is available for production of erythroid precursors which can lead to anemic condition due to the use of iron by bacteria during bacterial infection and shifting of iron to storage (Silver et al., 1997). Steven (2000) also stated that most anemias associated with bacterial and viral infections are caused by the inflammatory reaction. All the immunogen group calves showed decrease in the number of Red Blood Cells (RBC) but the OMP immunogen group calves showed significant decrease (p<0.05) in RBC counts where the decrease in RBC may due to inflammatory reactions caused by this immunogen as mentioned by Silver et al. (1997) and Steven (2000).
CONCLUSION
Measurement of haematological and biochemical parameters are important and reliable way to assess the health status of animals. This will help improve livestock production in Malaysia and elsewhere, as changes in these parameters are indicators of various disease conditions. The outcome of the present study shows that the undertaken approach is very useful and supports the prospect of routine check as a diagnostic tool in pasteurellosis based on its abnormal effects in blood levels.
ACKNOWLEDGMENT
We thank Mr. Mohd Jefri Bin Norsidin and Mr. Yap Keng Chee of the Department of Veterinary Clinical Studies, Universiti Putra Malaysia, for their technical assistance and Mr. Mohd Fahmi Bin Mashuri for taking adequate care of the experimental animals.
REFERENCES
- Brigham, K.L. and B. Meyrick, 1986. Endotoxin and lung injury. Am. Rev. Respir. Dis., 133: 913-927.
PubMedDirect Link - Brogden, K.A., H.D. Lehmkuhi and R.C. Cutlip, 1998. Pasteurella haemolytica complicated respiratory infections in sheep and goats. Vet. Res., 29: 233-254.
PubMed - Gilmour, M.I., C.M. Wathes and F.G.R. Taylor, 1990. The airborne survival of Pasteurella haemolytica and its deposition in and clearance from the mouse lung. Vet. Microbiol., 21: 363-375.
CrossRefDirect Link - Jacobsen, S., P.H. Andersen, T. Toelboell and P.M.H. Heegaard, 2004. Dose dependency and individual variability of the Lipopolysaccharide-induced bovine acute phase protein response. J. Dairy Sci., 87: 3330-3339.
Direct Link - Kamal, A.M., 2008. Some biochemical, haematological and clinical studies of selected ruminal and blood constituents in camels affected by various diseases. Res. J. Vet. Sci., 1: 16-27.
CrossRefDirect Link - Morris, C.A., N.R. Towers, W.D. Hohenboken, N. Maqbool, B.L. Smith and S.H. Phua, 2004. Inheritance of resistance to facial eczema: A review of research findings from sheep and cattle in New Zealand. New Zealand Vet. J., 52: 205-215.
CrossRefDirect Link - Odugbo, M.O., L.E. Odama, J.U. Umoh and A.A. Makinde, 2003. Serotypes of Pasteurella hemolytica from pneumonic lungs of sheep in Northern Nigeria. Small Ruminant Res., 48: 239-243.
Direct Link - Ozmen, O., S. Sahindurian, M. Haligur and M.K. Albay, 2008. Clinicopathological studies on facial eczema outbreak in sheep in Southwest Turkey. Trop. Anim. Health Prod., 40: 545-551.
CrossRefDirect Link - Rokke, O., A. Revhaug, B. Osterud and K.E. Giercksky, 1988. Increased plasma levels of endotoxin and corresponding changes in circulatory performance in a porcine sepsis model: The effect of antibiotic administration. Prog. Clin. Biol. Res., 272: 247-262.
PubMedDirect Link - Silver, B.J., B.D. Hamilton and Z. Toossi, 1997. Suppression of TNF-alpha gene expression by Hemin: Implications for the role of iron homeostasis in host inflammatory responses. J. Leukocyte Biol., 62: 547-552.
Direct Link - Smedegard, G., L. Lui and T.E. Hugli, 1989. Endotoxin-Induced shock in the rat: A role for C5a. Am. J. Pathol., 135: 489-497.
Direct Link - Warner, A.E., M.M. DeCamp Jr., R.M. Molina and J.D. Brain, 1988. Pulmonary removal of circulating endotoxin results in acute lung injury in sheep. Lab. Invest., 59: 219-230.
PubMedDirect Link