
Qian Zhang
College of Veterinary Medicine, Northwest A&F University, Yangling 712100, Shaanxi, China
Kang-Kang Guo
College of Veterinary Medicine, Northwest A&F University, Yangling 712100, Shaanxi, China
Yan-Ming Zhang
College of Veterinary Medicine, Northwest A&F University, Yangling 712100, Shaanxi, China
Chen Dai
College of Veterinary Medicine, Northwest A&F University, Yangling 712100, Shaanxi, China
Yan-Fen Jiang
College of Veterinary Medicine, Northwest A&F University, Yangling 712100, Shaanxi, China
Yang-Xin Peng
College of Veterinary Medicine, Northwest A&F University, Yangling 712100, Shaanxi, China
Asian Journal of Animal and Veterinary Advances
Year: 2013 | Volume: 8 | Issue: 5 | Page No.: 732-739







ABSTRACT
MicroRNAs (miRNAs) play important regulatory roles in mammalian cells and viral replication. Let-7 family of microRNA are important regulators in cell differentiation, human cancer and virus infection. The aim of the present study was to investigated miRNA which involved in Classical Swine Fever Virus (CSFV) infected Swine Umbilical Vein Endothelial Cells (SUVECs). Software predicting analysis was performed to screen out the potential miRNA. Four members of let-7 family, let-7a, let-7c, let-7f and let-7i were screened out as candidates. Luciferase activity assay showed that let-7a, let-7c and let-7i could bind to the 3' untranslated regions (3' UTR) of CSFV. Taqman qRT-PCR analyse showed that the inhibition or overexpression of let-7a and let-7c influenced the replication of CSFV. Mimics of let-7a and let-7c could significantly down-regulate CSFV replication. Further study showed that the expression of let-7a and let-7c were down-regulated in the CSFV infected cells and heterogeneous nuclear ribonucleoprotein A1( hnRNP A1), a negative regulator of let-7a, was up-regulated. The results indicated let-7a and let-7c could inhibit viral replication and the CSFV induced negative feedback regulation to interfere with let-7a expression. This study firstly suggested that miRNAs let-7a and let-7c play important roles in the replication of CSFV. These findings provide novel information on the interaction of miRNAs and the replication of CSFV.
PDF Abstract XML References Citation
Received: September 05, 2012;
Accepted: March 25, 2013;
Published: August 05, 2013
How to cite this article
Qian Zhang, Kang-Kang Guo, Yan-Ming Zhang, Chen Dai, Yan-Fen Jiang and Yang-Xin Peng, 2013. Members of the Let-7 Family Inhibit the Replication of Classical Swine Fever Virus in Swine Umbilical Vein Endothelial Cells. Asian Journal of Animal and Veterinary Advances, 8: 732-739.
DOI: 10.3923/ajava.2013.732.739
URL: https://scialert.net/abstract/?doi=ajava.2013.732.739
DOI: 10.3923/ajava.2013.732.739
URL: https://scialert.net/abstract/?doi=ajava.2013.732.739
INTRODUCTION
MicroRNAs (miRNAs) are endogenous RNAs of 21-22 nucleotides and are found in plants, invertebrates and vertebrates (Bartel, 2004). The biological functions of miRNAs include development, lifespan regulation, cell proliferation and differentiation and apoptosis; they also have roles in signaling pathways and metabolism (Cheng et al., 2005; Roush and Slack, 2008; Schmittgen, 2008). Animal miRNAs usually interact with the 3' noncoding regions (3' untranslated regions [3' UTRs]) of target mRNAs and cause downregulation of mRNA expression (Bartel, 2009; Chekulaeva and Filipowicz, 2009). Recently, Bioinformatics analyses coupled with experimental validations have revealed that miRNAs are not only important biomarkers, but they also confer virus resistance (Grassmann and Jeang, 2008; Jopling et al., 2005; Israsena et al., 2009). The H1N1 influenza A viruses (IAV), a member of Orthomyxoviridae, has two open reading frames (ORFs). One of them encoded PB1, which is involved in both transcription and replication of RNA genome. MiR-323, miR-491 and miR-654 inhibited replication of the IAV through binding to the PB1gene (Song et al., 2010). The other highly abundant miRNA, miR-29a which specifically targeted the HIV-1 3’UTR region. Inhibiting miR-29a enhanced HIV viral production and infectivity, whereas expressing a miR-29a mimic suppressed viral replication. Most studies have focused on the role of miRNAs in humans and few have demonstrated the role of miRNAs in animal diseases.
Classical Swine Fever (CSF) is a contagious and harmful disease. High-virulence strains of Classical Swine Fever Virus (CSFV) cause acute disease with high mortality rates and the disease is characterized by hemorrhagic fever, thrombocytopenia and disseminated intravascular coagulation (Summerfield et al., 1998).
CSFVs replicate in endothelial cells (ECs) in vitro and increase the expression of proinflammatory and procoagulant factors (Campos et al., 2004; Tang et al., 2010). Many functions of miRNAs in ECs have been confirmed that involved in disease (Bonauer et al., 2010; Doebele et al., 2009; Fish et al., 2008; Weber et al., 2010); however, whether cellular miRNAs play a role in CSFV infection is unknown. A microRNA catalog of Swine Umbilical Vein ECs (SUVECs), an established cell line, have been identified by deep sequencing. One or more cellular miRNA were hypothesized could be involved in the replication of CSFV. In this study, let-7a and let-7c was predicted and screened out , which inhibited the replication of CSFV in SUVECs. At the same time the study demonstrated the expression of let-7a was negative regulated by HnRNP A1 in CSFV infected SUVECs. The study provide a sight into viral and host cell interaction.
MATERIALS AND METHODS
MiRNA target prediction: MiRNA target prediction was performed to determine the miRNAs involved in CSFV infection in SUVECs. The algorithm RNA22 (http://cbcsrv.watson.ibm.com/rna22.html) was used to predict the miRNA-binding site (Miranda et al., 2006).
Cell culture and reagents: SUVECs were cultured in M199 (Gibco) supplemented with 20% Fetal Bovine Serum (FBS) (Hyclone) and 100 mg mL-1 penicillin/streptomycin at 37°C and 5% CO2. Let-7 mimics and inhibitors were synthesized and purified by GenePharma Co. (Shanghai, China).
MicroRNA real-time reverse transcription polymerase chain reaction: Total RNA was isolated using Trizol reagent (Invitrogen), reverse-transcribed using PrimeScript RT reagent kit (TaKaRa) and used for two-step quantitative reverse transcription polymerase chain reaction (RT-PCR). The resulting cDNA was amplified using Sybr Premix Ex Taq kit(TaKaRa) and the ssc-let-7a Bulge-Loop miRNA qPCR primer sets (RiboBio) to detect miRNA expression. The synthesized let-7 was used for generating standard curves. To elucidate the effects of CSFV infection on let-7 expression, cells were infected with CSFV Shimen strain and maintained in Dulbecco’s modified Eagle’s medium (DMEM) (Gibco) supplemented with 2% FBS for 12 and 24 h; then, total RNA was isolated and used for detecting let-7 expression. The reactions were monitored by iQ5 multicolor real-time PCR detection system. The PCR conditions were as follows: preheating at 95°C for 20 sec and 40 cycles at 95°C for 10 s, 60°C for 20 sec and 70°C for 30 sec. U6 was quantified as an endogenous control.
Dual-luciferase assay: The 3' UTR of CSFV was predicted to contain the putative let-7 target site. The 3' UTR containing the let-7 target site or a sequence of the 3' UTR mutated in the putative seed region for let-7 binding was inserted into the NotI and XhoI sites of the psiCHECK-2 vector (Promega). The constructs, named psiCHECK-2-CSFV and psiCHECK-2-CSFV-mut, respectively, were transfected into SUVECs. The cells were seeded in 96-well plates at 70% confluence before transfection. The SUVECs were co-transfected with let-7 mimics or inhibitors by using Lipofectamine 2000 (Invitrogen). Firefly and Renilla luciferase activities were measured 48 h after transfection by using the Dual-Glo Luciferase Assay System (Promega), according to the manufacturer’s instructions. The psiCHECK-2 control vector and let-7 inhibitor or mimic controls were used as negative controls. Each experiment was repeated 3 times.
CSFV real-time RT-PCR: In order to confirm that let-7 can regulate the replication of CSFV, SUVECs were transfected with let-7 inhibitors or mimics by using Lipofectamine 2000, according to the manufacturer’s instructions. Twelve hours after transfection, the cells were infected with CSFV Shimen strain and maintained in DMEM supplemented with 2% FBS for 48 h. The cells were then washed in 0.01 M phosphate-buffered saline (PBS) 3 times and total RNA was isolated using Trizol reagent. One-step quantitative RT-PCR was performed using one-step PrimeScript RT-PCR kit (TaKaRa). The absolute level of CSFV was determined using TaqMan assay under the following conditions: 42°C for 10 min; 95°C for 30 sec; and 40 cycles at 95°C for 15 sec, 57°C for 20 sec and 72°C for 15 sec. The primers used were as follows: sense, 5'-GTTCTGCGAGGTGACCAAAAG-3' and antisense, 5'-GATGCACACATAAGTATGGTATAAGC-3'. The TaqMan probe used was 5'-(FAM) TCCGTCGCTACCTGTCACCCTACCT (Eclipse)-3'. The CSFV RNA standards were used for preparing standard curves.
Real-time RT-PCR assay for heterogeneous nuclear ribonucleoprotein A1 mRNA: To determine a potential factor downregulating let-7a, SUVECs were infected with CSFV Shimen strain and overlaid with 2 mL culture medium containing 2% FBS. The cells were harvested at 12 and 24 h and stored at -70°C. For heterogeneous nuclear ribonucleoprotein A1 (hnRNP A1) mRNA detection, total RNA was extracted using Trizol reagent, resuspended in RNase-free water and reverse-transcribed using PrimeScript RT reagent kit (TaKaRa) and random primers at 42°C for 30 min and 85°C for 10 sec. The resulting cDNA was amplified using Sybr Premix Ex Taq kit(TaKaRa). The primers used were as follows: sense, 5'-GATGACCATGACTCCGTAGACAAG-3' and antisense, 5'- GGATAGGGCTTTCCTCACTTCAC-3'. The PCR conditions were as follows: preheating at 95°C for 20 sec, followed by 40 cycles at 95°C for 10 sec, 60°C for 20 sec and 70°C for 30 sec. β-actin mRNA was quantified as an endogenous control.
Statistical analysis: All quantitative data were analyzed using two-tail homoscedastic Student’s t-test in 2011. In all figures, values are expressed as Mean±standard deviation (SD) and p<0.05 was considered statistically significant. The data generated in vitro were representative of at least 3 independent experiments. SPSS11.5 software was used for statistical analysis.
RESULTS
MiRNA target prediction showed that members of the let-7 family can bind, according to Watson and Crick base-pairing, to the 3' UTR of CSFV RNA through 8 nucleotides in the seed sequence of the let-7 family (Table 1).
| Table 1: | The let-7 target site along the 3' untranslated regions (3' UTRs) of classical swine fever virus (CSFV) is predicted by RNA22 software. The underlined sequences are the seed-matched sites |
 | |
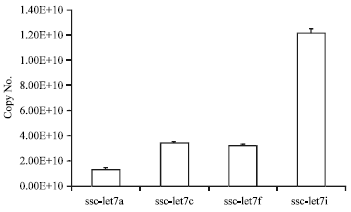 | |
| Fig. 1: | Results of quantitative polymerase chain reaction (PCR) of 4 members of the let-7 family in swine umbilical vein endothelial cells |
The expression of let-7, including let-7a, let-7c, let-7f and let-7i, in SUVECs were detected by using two-step quantitative RT-PCR (Fig. 1). Let -7i is the highest expression level in SUVECs, let-7a is the lowest in the minimum amount.
To examine whether let-7 can bind to the putative target site in the CSFV 3' -UTR, dual-luciferase reporter plasmids psiCHECK-2-CSFV and psiCHECK-2-CSFV-mut was constructed, which contained the putative let-7 binding site of CSFV 3' UTR and a mutated sequence of the 3' UTR, respectively, downstream of the Renilla luciferase gene (Fig. 2d). These vectors were co-transfected with let-7 inhibitors or mimics into SUVECs. The luciferase activities of cells with let-7 mimics were markedly reduced, except let-7f mimics group (Fig. 2a) and these activities increased in cells with let-7 inhibitors, except in the case of let-7f (Fig. 2b). The luciferase activity of psiCHECK-2-CSFV-mut group was not evident (Fig. 2c). These data suggested that CSFV was one of the potential targets of let-7 in SUVECs.
To determine whether let-7 can regulate the replication of CSFV, SUVECs were transfected with let-7 mimics or inhibitors. About 12 h after transfection, SUVECs were infected with CSFV Shimen strain and 48 h after infection, the copy number of CSFV was determined. The result showed a decrease in the copy number of CSFV in the let-7 mimic-transfected group compared to the negative control group (Fig. 3a), except in case of let-7i, but this effect was not evident in the let-7 inhibitor-transfected group (Fig. 3b). Further, presumed that the expressions of let-7a and let-7c may be regulated by CSFV during the incubation period. These expressions were decreased when the SUVECs were infected after 12 and 24 h, which suggested that CSFV can downregulate the expressions of let-7a and let-7c (Fig. 4a, 4b). HnRNP A1 is a negative regulator of mature let-7a. The expression of hnRNP A1 mRNA were examined in infected SUVECs at 12, 24 and 48 h. As expected, the results showed that hnRNP A1 mRNA was upregulated in CSFV-infected SUVECs (Fig. 4c).
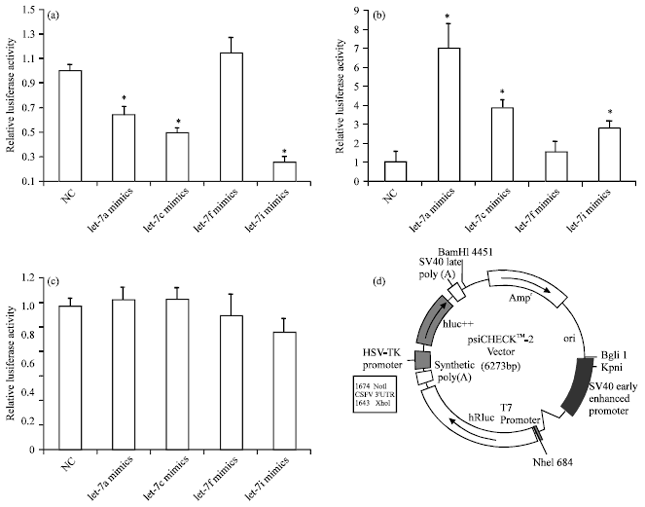 | |
| Fig. 2(a-d): | Effect of let-7 on the expression of classical swine fever virus (CSFV), determined by luciferase reporter assay, (a, b) The relative luciferase activity of psiCHECK-2-CSFV after co-transfection with let-7 mimics or inhibitors in swine umbilical vein endothelial cells (SUVECs), (c) The relative luciferase activity of psiCHECK-2-CSFV-mut after co-transfection with let-7 inhibitor in SUVECs, (d) Circular map of the psiCHECK-2 vector. The 3' untranslated regions (3' UTRs) of CSFV were inserted into the XhoI and NotI sites. The data were normalized by the ratio of firefly and Renilla luciferase activities (n = 3; *p<0.05) |
DISCUSSION
MiRNAs are a large family of post-transcriptional regulators of gene expression. More recently, such regulation has an important role in function of miRNA(Krol et al., 2010). MiRNAs have been implicated in the pathology of various diseases, including virus. Virus-encoded miRNAs of EB virus was first found by Pfeffer et al. (2004). So far, Virus-encoded miRNAs were clone from the Herpesviridae, Adenoviridae and Polyomaviridae (Takane and Kanai, 2011). And found that they were able to regulate the related gene of the virus and host cell. Recently gradually reported that cellular miRNAs infection were very important to the replication of the virus in cells. Some bioinformatics and experimentally demonstrated miRNA can be as target to the virus, including HIV, HCV. A cluster of human miRNAs including miR-28, miR-125b, miR-150, miR-223 and miR-382 was confirm to targeted for the 3’UTR of HIV-1 mRNAs (Wang et al., 2011). MiR-122 is specifically expressed and is highly abundant in the human liver.
 | |
| Fig. 3(a-b): | MiRNAs inhibit the replication of classical swine fever virus (CSFV). (a) Real-time reverse transcription polymerase chain reaction was used to detect the effects of let-7 mimics on CSFV replication in swine umbilical vein endothelial cells (SUVECs). (b) The effects of let-7 inhibitors on the replication of CSFV in SUVECs (n = 3; *p<0.05) |
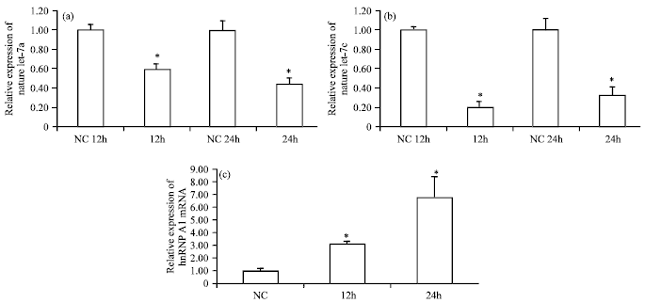 | |
| Fig. 4(a-c): | The relative expressions of let-7a and let-7c in classical swine fever virus (SCFV)-infected swine umbilical vein endothelial cells (SUVECs), (a) Real-time polymerase chain reaction (PCR) was used to detect the expression of let-7a in SUVECs 12 and 24 h after infection, (b) The expression of let-7c in SUVECs 12 and 24 h after infection, (c) Real-time PCR was used to detect the expression of hnRNP A1 mRNA in SUVECs 12 and 24 h after infection (n = 3; *p<0.05) |
Sequestration of miR-122 in liver cells results in a marked loss of autonomously replicating hepatitis C viral RNAs (Jopling et al., 2006).
However, there is no direct evidence to show the role of MiRNAs in the infection of CSFV. Therefore, in this study cellular miRNAs interacted with CSFV were explored. Two cellular miRNA were screened out which could inhibit the replication of the CSFV through the degradation CSFV RNA in SUVECs. The result suggested that cellular miRNA are factors in the host’s resistance to CSFV infection, which play a role in CSFV prevention. It is well known that CSFV has a major impact on the socioeconomic damage. CSFV is a member of the Flaviviridae family. The virion contains 12.3kb single-stranded RNA genome of positive polarity. CSFV has only one open reading frames (ORFs) which is involved in both transcription and replication of RNA genome. Bioinformatics software predicted that let-7 family target for 3’UTR of CSFV gene. Let-7 family was highly conserved acrossed animal species and involved in physiological functions and pathologic processes (Roush and Slack, 2008). Let-7 family mainly targeted for 3’UTR of gene in acknowledged research. The study showed that two members of let-7 family, let-7a and let-7c mimics could inhibit the replication of CSFV in vitro. The study was performed in a single cell line in vitro. The result indicated that may be more specificity miRNAs involved in CSFV infection in vivo, especially in Viral target organs.
While growing evidence has showed that virus regulated the expression cellular miRNA to protect them against the cell’s RNAi restriction. Yu found that hepatitis B virus x protein down-regulation let-7a supports cell proliferation in HBx cells (Wang et al., 2010). In this study, the expression levels of the let-7a and let-7c were downregulation during CSFV infection in SUVECs. And HnRNP A1 a negative regulator of let -7a, binds the conserved terminal loop of pri-let-7a and inhibits its processing by the Drosha complex (Michlewski and Caceres, 2010). HnRNP A1 was hypothesized that could be upregulated in infected SUVECs. The result conformed the hypothesized that the expression of hnRNP A1 was upregulate in 24 h. The downregulation of miRNA in infected SUVECs reflected the mechanism of the virus defense against the antiviral factors during the early staged of infection.
In conclusion, our results indicated that let-7a and let-7c targeted the CSFV gene can inhibit relication of virus in SUVECs. And confirmed CSFV induced hnRNP A1 negative feedback regulation to let-7a expression. The host cells blocked viral invasion by miRNA, while the virus can resist this interference effect to create favorable condition for its own replication. The findings indicated that there was dual directional regulation mechanism between cell miRNAs and virus in the cells.
ACKNOWLEDGMENTS
We wish to thank Zongsong Wu for designing the experiments and Pengbo Ning and Dr. Qinghai Tang for reviewing this article. This work was supported by grants from the National Natural Science Foundation of China (No. 30771607).
REFERENCES
- Bartel, D.P., 2004. MicroRNAs: Genomics, biogenesis, mechanism and function. Cell, 116: 281-297.
PubMed - Bartel, D.P., 2009. MicroRNAs: Target recognition and regulatory functions. Cell., 136: 215-233.
PubMedDirect Link - Bonauer, A., R.A. Boon and S. Dimmeler, 2010. Vascular micrornas. Curr. Drug Targets, 11: 943-949.
Direct Link - Campos, E., C. Revilla, S. Chamorro, B. Alvarez, A. Ezquerra, J. Dominguez and F. Alonso, 2004. In vitro effect of classical swine fever virus on a porcine aortic endothelial cell line. Vet. Res., 35: 625-633.
CrossRefDirect Link - Chekulaeva, M. and W. Filipowicz, 2009. Mechanisms of mirna-mediated post-transcriptional regulation in animal cells. Curr. Opin. Cell Biol., 21: 452-460.
CrossRef - Cheng, A.M., M.W. Byrom, J. Shelton and L.P. Ford, 2005. Antisense inhibition of human mirnas and indications for an involvement of mirna in cell growth and apoptosis. Nucleic Acids Res., 33: 1290-1297.
Direct Link - Fish, J.E., M.M. Santoro, S.U. Morton, S. Yu and R.F. Yeh et al., 2008. miR-126 regulates angiogenic signaling and vascular integrity. Dev. Cell, 15: 272-284.
PubMed - Grassmann, R. and K.T. Jeang, 2008. The roles of micrornas in mammalian virus infection. Biochim. Biophys. Acta., 1779: 706-711.
Direct Link - Jopling, C.L., K.L. Norman and P. Sarnow, 2006. Positive and negative modulation of viral and cellular mrnas by liver-specific microrna mir-122. Cold Spring. Harb. Symp. Quant. Biol., 71: 369-376.
CrossRef - Israsena, N., P. Supavonwong, N. Ratanasetyuth, P. Khawplod and T. Hemachudha, 2009. Inhibition of rabies virus replication by multiple artificial micrornas. Antiviral. Res., 84: 76-83.
CrossRef - Krol, J., I. Loedige and W. Filipowicz, 2010. The widespread regulation of microRNA biogenesis, function and decay. Nat. Rev. Genet., 11: 597-610.
CrossRef - Song, L., H. Liu, S. Gao, W. Jiang and W. Huang, 2010. Cellular micrornas inhibit replication of the h1n1 influenza a virus in infected cells. J. Virol., 84: 8849-8860.
Direct Link - Michlewski, G. and J.F. Caceres, 2010. Antagonistic role of hnRNP A1 and KSRP in the regulation of let-7a biogenesis. Nat. Struct. Mol. Biol., 17: 1011-1018.
CrossRefDirect Link - Miranda, K.C., T. Huynh, Y. Tay, Y.S. Ang and W.L.Tam et al., 2006. A pattern-based method for the identification of microrna binding sites and their corresponding heteroduplexes. Cell, 126: 1203-1217.
CrossRefDirect Link - Pfeffer, S., M. Zavolan, F.A. Grasser, M. Chien and J.J. Russo et al., 2004. Identification of virus-encoded MicroRNAs. Science, 304: 734-736.
CrossRef - Roush, S. and F.J. Slack, 2008. The let-7 family of microRNAs. Trends Cell Biol., 18: 505-516.
CrossRef - Schmittgen, T.D., 2008. Regulation of microrna processing in development, differentiation and cancer. J. Cell. Mol. Med., 12: 1811-1819.
CrossRef - Summerfield, A., S.M. Knotig and K.C. McCullough, 1998. Lymphocyte apoptosis during classical swine fever: Implication of activation-induced cell death. J. Virol., 72: 1853-1861.
Direct Link - Takane K. and A. Kanai, 2011. Vertebrate virus-encoded MicroRNAs and their sequence conservation. Jpn. J. Infect. Dis., 64: 357-366.
PubMedDirect Link - Tang, Q.H., Y.M. Zhang, Y.Z. Xu, L. He, C. Dai and P. Sun, 2010. Up-regulation of integrin beta 3 expression in porcine vascular endothelial cells cultured in vitro by classical swine fever virus. Vet. Immunol. Immunop., 133: 237-242.
CrossRefDirect Link - Wang, X., L. Ye, Y. Zhou, M.Q. Liu, D.J. Zhou and W.Z. Ho, 2011. Inhibition of anti-HIV MicroRNA expression: A mechanism for opioid-mediated enhancement of HIV infection of monocytes. Am. J. Pathol., 178: 41-47.
PubMedDirect Link - Weber, M., M.B. Baker, J.P. Moore and C.D. Searles, 2010. MiR-21 is induced in endothelial cells by shear stress and modulates apoptosis and eNOS activity. Biochem. Biophys. Res. Commun., 393: 643-648.
CrossRef