
H. Lu
College of veterinary medicine, Northwest A and F University, Yangling-712100, Shaanxi, Peoples� Republic of China
S.S. Wang
College of veterinary medicine, Northwest A and F University, Yangling-712100, Shaanxi, Peoples� Republic of China
B.Y. Zhao
College of veterinary medicine, Northwest A and F University, Yangling-712100, Shaanxi, Peoples� Republic of China
Asian Journal of Animal and Veterinary Advances
Year: 2012 | Volume: 7 | Issue: 9 | Page No.: 822-831







ABSTRACT
Locoweeds cause significant livestock poisoning and economic loss all over the world. The principal compound responsible for its toxicity is the indolizidine alkaloid swainsonine, a new potential anticancer and antiviral drug. O. glabra is a locoweed found mainly distributed throughout the inner Mongolia region of China and presents a serious hazard to the local livestock industry. The purpose of this study was to analyze the main toxic products of O. glabra first by TLC and GC-MS methods and then to evaluate its alkaloids’ toxicity in vivo. Twelve Sprague-Dawley rats were randomly divided into control group and experimental group in which the rats received 0 and 800 mg kg-1 (swainsonine content 88.64 mg) body weight day-1 of crude alkaloid extract by intragastric administration for 65 days. Results showed that a white needle crystal was isolated and purified through systematic extraction of the alkaloid by a silica gel column and gradient sublimation from O. glabra; it was identified to be swainsonine (SW) according to physicochemical properties and spectral data analyses. Pathological lesions were revealed in the brain, heart, liver and kidney of alkaloid-treated rats. We also observed diffuse vacuolation of neurons, epithelial cells of renal tubules by electron microscope. These histopathological changes demonstrated that the main toxic ingredient of O. glabra is swainsonine.
PDF Abstract XML References Citation
Received: February 09, 2012;
Accepted: March 29, 2012;
Published: May 10, 2012
How to cite this article
H. Lu, S.S. Wang and B.Y. Zhao, 2012. Isolation and Identification of Swainsonine from Oxytropis glabra and its Pathological Lesions to SD Rats. Asian Journal of Animal and Veterinary Advances, 7: 822-831.
DOI: 10.3923/ajava.2012.822.831
URL: https://scialert.net/abstract/?doi=ajava.2012.822.831
DOI: 10.3923/ajava.2012.822.831
URL: https://scialert.net/abstract/?doi=ajava.2012.822.831
INTRODUCTION
Locoweeds are a general term for those plants of the genera Astragalus and Oxytropis that contain the indolizidine alkaloid swainsonine (SW) (Dorling et al., 1980; Molyneux and James, 1982). They are perennial herbs that grow in clumps with pinnately compound leaves, lacking tendrils, they have pea-like flowers (blue, white or purple) and often inflated pods, both short and long. Furthermore, they are widely distributed around the world in the United States (Taylor and Strickland, 2002), Australia (Martyn et al., 2003), Mexico (Smith et al., 1992), Brazil (Medeiros et al., 2003) and China (Cao et al., 1992). In China, locoweeds are distributed in the western part and cover land area up to 110,000 km2 (Zhao et al., 2003).
Oxytropis glabra, also known by the common names of zuimacao (in inner Mongolia) or mabanchang (in Ningxia province), is one of the locoweeds from the genus Oxytropis. It is a perennial ryegrass which usually grows in the drought desert areas. It is mainly distributed in a large area of Inner Mongolia, Shaanxi, Shanxi, Ningxia and most grassland in Xinjiang province.
In the drought years when most other pastures grow sparsely, O. glabra grows well. This induces hungry domestic animals to eat it and when their ingestion amount reaches a certain level, the animals show clinical symptoms characterized by head level tremors and dyskinesia. Mortality due to O. glabra ingestion can bring enormous economic loss in local animal husbandry. The percentage of livestock that died from poisoning caused by O. glabra can reach 5 to 10% each year in Aksu of Xinjiang province and up to 50% in serious cases (Sun and Gao, 1993). However, O. glabra can also be used as an anesthetic, sedative and analgetic medicine for alleviating the symptoms of diseases such as arthritis, odontalgia, neurasthenia, etc., (Jiangsu New Medical College, 1977).
Foreign scholars have come a long way to discover the toxic ingredients of locoweed and significant effort has been devoted to their study for nearly a century. They found that the toxic ingredients of locoweed could be divided into three groups: aliphatic nitro compounds, selenium or selenium compounds and locoweed toxin. However, native scholars have different ideas about the toxic ingredients of locoweed. It has been confirmed that the locoweed toxin alkaloids are responsible for locoweed intoxication, instead of aliphatic nitro compounds or selenium and selenium compounds. The alkaloids, which have already been isolated and identified from locoweed, can be divided into 3 groups according to their structures: Indolizidine alkaloids (swainsonine and nitrogen oxide swainsonine), quinolizidine alkaloids (anagyrine, thermopsine, lupinine and n-methylcytisine) and piperidine alkaloids (2,2,6,6-tetramethyl-4-piperidone). The indolizidine alkaloid SW, the main toxic ingredient of locoweed, was also isolated from O. ochrocephala, O. kansuensis, A. variabilis and A. strictus; each study reported that SW is the main toxic compound from each plant (Cao et al., 1989; Boliang et al., 1995; Huang et al., 1992; Zhao et al., 1993; Tong et al., 2001). SW is a potential inhibitor of α-mannosidase which can inhibit the activity of lysosomal α-mannosidase and Golgi α-mannosidase, resulting in the accumulation of oligosaccharides and cellular vacuolation (Tulsiani et al., 1984). Previous studies have reported that the main toxic compounds from O. glabra are alkaloid but the composition of the alkaloids has not yet been analyzed (Yang et al., 1989; Yu et al., 1991). Thus the principal compound responsible for the toxicity of O. glabra remains unknown. The objective of this study was to determine the main toxic ingredient of the O. glabra and its toxicity to Sprague-Dawley (SD) rats in order to efficiently utilize SW for medicinal sources as well as to protect domestic animals from poisoning.
MATERIALS AND METHODS
Plant materials: The aerial part of O. glabra was collected from a grassland in Alasan Left Banner, where the locoism often occurred, from Inner Mongolia Autonomous Region in July 2009, then taxonomically identified by Researcher Zhang Yue-Jin, College of Life Science, Northwest A and F University, China. It was subsequently dried in the shade, finely ground and comminuted afterwards.
Chemical reagents and apparatus: Chloroform (AR), Silica Gel GF254 was used for TLC (batch number 990706) with grade size over 200 meshes (Qingdao Haihua Dehumidizer Factory, China). Silica Gel (batch number 010830) was used for column chromatography with grade size ranging from 100 to 200 meshes (Qingdao Haihua Dehumidizer Factory, China). SW standard sample was obtained from SIGMA Corporation, USA. Apparatus used included: Rotary Evaporator RE-52AA (Shanghai Yarong Biochemical Equipment Factory, China), X-4 micro melting apparatus (uncorrected) (Peking No.3 Optical Instrument Factory, China), NICOLET AVATAR 360 FT-IR spectrometer (made in the USA) and Velocitron VG-ZAB-HS (HP Corporation, USA).
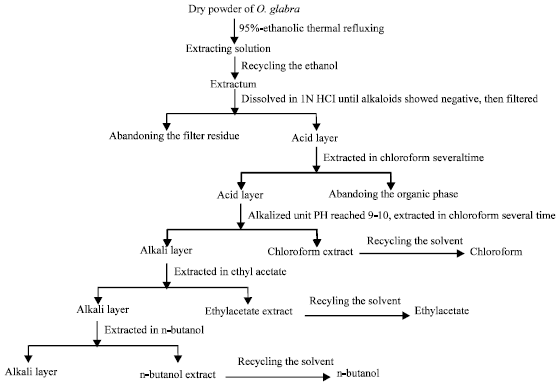 | |
| Fig. 1: | Flow diagram of the process for alkaloid isolation from O. Glabra. Dry powder of O. Glabra was thermal refluxed in aqueous EtOH and crude extraction of alkaloid was obtained under reduced pressure, then the aqueous fraction was successively extracted by chloroform, ethyl acetate and n-butanol |
Animals: The animals used in this study were approved by the Animal Care Committee of Xi’an Jiaotong University. Sprague-Dawley rats were purchased from Experimental Animal Center of College of Medicine, Xi’an Jiaotong University (Xi’an, China).
Isolation of alkaloids in O. glabra: The plant sample (15 kg) was thermal refluxed in 95% aqueous EtOH 4 times, 4 h each cycle, followed by the recovery of EtOH under reduced pressure to obtain crude extraction for alkaloid extraction and isolation. The extraction was dissolved in 1 N HCl and the filtrate was then extracted by chloroform and alkalized. The aqueous fraction was successively extracted by chloroform, ethyl acetate and n-butanol (Fig. 1).
Analysis of alkaloids in O. glabra
Thin-layer chromatography (TLC): TLC detection was performed on silica gel G precoated plates with the solvent chloroform:methanol:ammonia:water (70:26:2:2, V/V), chloroform:methanol:ammonia:water (70:26:10:10, V/V) as well as methanol:ethylacetate: ammonia (4:1:1, V/V) were the developing solvents and modified potassium heptaiodobismuthate reagent or H2O2/10% acetic anhydride in EtOH/Ehrlich’s reagent was used as the chromogenic agent.
The extracts were dissolved in methanol, spotted onto the silica gel G precoated plates GF254. Plates were developed with an ascendant run after saturation with the mobile phase in a glass chamber for 5-10 min. The plates were dried as soon as the mobile phase reached 10 mm from the front edge of the plates. The plates were stained successively with a spray of H2O2 (heated for 10 min in an oven at 115°C), a spray of 10% acetic anhydride in dehydrated alcohol (heated at the same temperature until the smell of acetic anhydride disappeared) and finally a spray of Ehrlich’s reagent (heated for 15 min at 120°C). The color of the spots in each plate was recorded and the Rf was worked out.
Detection of SW: GC detection method was used and 0.04028, 0.02478 and 0.04858 g chloroform, ethyl acetate and n-butanol extracts, respectively were weighed accurately, dissolved in 2 mL pyridine and then 0.1 mL were measured and added to equivalent BSTFA reagent. The mixture was incubated in 50°C water bath for 30 min and 1-2 pL reactant was used for GC. The SW standard was treated in the same way. The conditions of GC were as follows: DEGS chromatographic column was 2 m long and the column temperature was 200, 80°C min-1, N2 with a speed of 25 mL min-1 was used as the carrier gas. The FID detector’s temperature was 250°C and the injector port was 250°C.
Isolation of SW: The extraction of n-butanol was eluted by eluant (chloroform:methanol:ammonia:water = 70:26:2:2, V/V) in the G gel chromatography. Eluted fractions were obtained every 5 mL and detected by TLC. Similar fractions were put together and the results were compared with the standard SW sample. These SW containing fractions were purified by sublimation under reduced pressure after treatment with alkaline chloroform. The compound was then detected by TLC using at least three developing solvent. It could be identified as a single compound if the result showed a single point, for which Rf and color was then compared with standard SW sample.
Structure identification: The X-4 micro melting apparatus (uncorrected), NICOLET AVATAR 360 FT-IR spectrometer, Velocitron VG-ZAB-HS and nuclear magnetic resonance apparatus were used to identify the structure of the isolated compound. To confirm the structure, the data was analyzed by spectral analysis and compared with data reported in related literature (Cao et al., 1989; Huang et al., 1992; Tong et al., 2001).
Analysis of GC-MS: Collected samples were dissolved by ethyl acetate. The gas chromatograph conditions: Hp-1 capillary column (30 mx0.33 mmx0.25 pm) was used with a temperature of 100-270°C, which raised at a speed of 8°C min-1. N2 was used as carrier gas with a speed of 1.2 mL min-1. Split injection and split ratio was 20:1. Injector temperature was 300°C and the detector was FID. The conditions of MS: EI ionization mode, 70 eV ionization voltage, ion power temperature: 230°C. The scanning range of ion mass: 15-500 amu.
Alkaloids poisoning: These experiments were performed in male and female Sprague-Dawley rats (200-220 g of body weight). After a week of adaptation period in a room with controlled temperature (21±1°C) and lighting (12 h light/12 h dark), twelve SD rats each were assigned to control and experimental groups (800 mg kg-1 Alkaloids, containing 88.64 mg kg-1 SW). Alkaloid solutions prepared in distilled water were given as drinking water ad libitum to SD rats. After drinking 65 days, all SD rats were anesthetized with ether vapor and sacrificed by decapitation; brain, heart, liver and kidney were quickly collected and refrigerated in 4°C.
Transmission electron microscopy: Samples from brain, heart, liver and kidney were prefixed in 2.5% glutaraldehyde solution and diced into 1 mm3, followed by three 15 min rinses with 0.1 M phosphate buffer (pH 7.4). Post-fixation samples were incubated in cold 1% aqueous osmium tetroxide for 1 h. After rinsing with phosphate buffer again, the specimens were dehydrated in a graded ethanol series of 50-100% and then embedded in Epon 812. Ultrathin sections were sliced with glass knives on a LKB-V ultramicrotome (Nova, Sweden), stained with uranyl acetate and lead citrate and finally examined under a TecNai 12 transmission electron microscope (Philips Company, Holland).
RESULTS
Isolation of alkaloids from O. glabra: Gross alkaloid extract was isolated from O. Glabra as by alcohols solvent protocol as detailed in methods. The contents of this gross alkaloid extract were further extracted by different organic solvents indicating polarity, with results listed in Table 1. It indicates that paste-forming rate of O. glabra is 21.7%. The percentage extraction of alkaloids by organic solvents was as follows: 0.04% by chloroform, 0.14% by ethyl acetate and 1.10% by n-butanol, occupying 3.39, 10.75 and 85.86% of the whole alkaloids, respectively. This indicates that the predominant component of the alkaloids found in O. glabra is of maximum polarity.
TLC detection: The alkaloids extracted by different organic solvents appear in a line after being developed by the developing solvent and colorated by modified potassium heptaiodobismuthate and there are no single points. At least 9 alkaloids in different extracts appeared clearly and discretely after coloration by H2O2, 10% acetic anhydride dehydrated alcohol and Ehrlich’s reagent. Among these are more kinds of alkaloids in the chloroform and ethyl acetate extracts. We can see that there are points similar to SW’s in all the extracts of O. glabra after comparing them with the SW standard sample. The Rf of different extractums are as follows: SW standard sample, 0.47 (prunosus); chloroform, 0.48 (violet); ethyl acetate extractum, 0.49 (prunosus); n-butanol extractum, 0.46 (prunosus).
GC-MS detection: Considering the results of TLC detection, we hypothesized that there is likely some SW present in O. glabra, so we detected the content of SW in different extractums from O. glabra using GC-MS. It can be seen that most of the SW was in the n-butanol extractum for O. glabra (Table 2).
| Table 1: | Extraction of alkaloids in O. glabra |
 | |
| Alkaloids in O. glabra were extracted by three organic solvent, the result indicates that extraction rate of alkaloids of O. glabra is maximum in N-butanol fractions | |
| Table 2: | SW content in different extractums from O. glabra |
 | |
| The content of SW was detected to different extractums of O. glabra by GC-MS, most of the SW was in the n-butanol extractum for O. glabra | |
Isolation and identification of SW: A white, acicular crystal with a melting point of 144-145°C had been isolated after sublimation. It was weighted 0.0625 g and the extraction rate was 0.002%. Rf was 0.59, similar to SW standard sample according to TLC detection, in which the developing solvents were chloroform:methanol:ammonia:water (70:26:2:2) and the chromogenic agents were 30% H2O2/10% H2O2/10% acetic anhydride dehydrated alcohol/Ehrlich’s reagent. IR max (KBr, cm-1): 3423 (O-H), 2943, 2891 (C-H), 2828-2724 (Bohlman zone), 1441-1347 (C-H, O-H), 1324-1222 (C-N), 1072 (C-O) absorption peak. EI-MSm/e: 173 (M+), 155 (M-H2O), 138 (M-H2O-H), 116, 115, 96, 84, 72, 43 (fragmentation peak of the mother nucleus); 1HNMR (400 M, D2O, ppm)δ: 4.340 (m, J = 2.0 and 8.0 HZ, H-2), 4.245 (dd, J = 4.0 and 5.6 Hz, H-1), 3.792 (td, J = 4.4 and 10.4 Hz, H-8), 2.868 (dd, J = 2.0 and 8.0 Hz, H-3eq), 2.543 (dd, J = 8.0 and 11.2 Hz, H-3ax), 2.031 (m, H-7eq), 1.945 (m, H-8a), 1.918 (m, J = 3.6 and 12.8 Hz, H-5ax), 1.708 (br, d, J = 13.6 Hz, H-6eq), 1.504 (qt, J = 4.0 and 13.2 Hz, H-6ax), 1.228 (qd, J = 4.0 and 12.8 Hz, H-7ax). 13CNMR (400M, D2O, ppm) δ: 75.1 (C-2), 72.0(C-1), 71.4 (C-8), 68.7 (C-8a), 62.9 (C-3), 54.0 (C-5), 34.8 (C-7), 25.5 (C-6). The data above showed that the white needle crystal was SW.
Ultrastructure changes: In the cerebral cortex, neurons showed vacuolation in the mitochondria and nucleus (Fig. 2a); there was increased space around glial cellular nuclei, which was also called edema in glial cells.
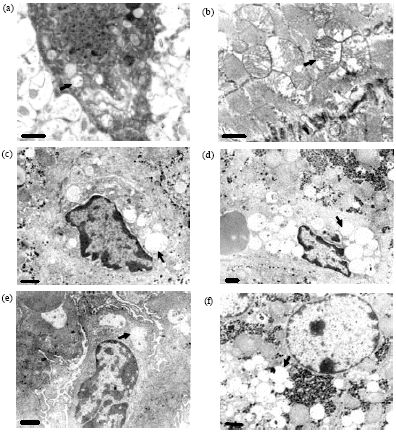 | |
| Fig. 2(a-f): | Ultrastructure views of the different tissues of the SD rats. (a) Vacuolization in the mitochondria of neurons, x14500, (b) Vacuolization in the mitochondria of the myocardial cells, x14500, (c) Vacuolization in Kuppfer cells, x8700, (d) Vacuolization in endothelial cells of hepatic sinusoids, x5800, (e) Vacuolization in glomerular epithelial cells, x7250 and (f) Vacuolization in cytoplasm of liver cells, x7250 (arrows) |
The space around the capillaries increased (again called edema) and vacuolation was observed in endothelial cells. In the heart, the mitochondria of the myocardial cells showed vacuolation (Fig. 2b). In the liver, vacuolation was observed in the cytoplasm of hepatocytes (Fig. 2c), Kupffer cells (Fig. 2d) and the endothelial cells in hepatic sinusoids (Fig. 2e). In the kidneys, vacuolation was observed in the glomerulus epithelial cells (Fig. 2f) and in the mitochondria of the epithelial cells in the renal tubule, in which the number of lysosomes was increased.
DISCUSSION
Ingestion of locoweed by livestock, such as the poisonous Astragalus and Oxytropis genera, is mainly characterized by symptoms of the Central Nervous System (CNS) such as depression, lethargy, asthenia and tremor. It is the most dangerous toxic plant, even more dangerous than Loimia and snowstorm, threatening the development of animal husbandry in grassland all over the world. Taking the Alasan Left League as an example, there were 5 million ha. of available grassland, 2.33 million ha. of which were covered with poisonous weeds: O. glabra, Astragalus variabilis, Achnatherum inebrians and Cynanchum komarovii (Dong et al., 2003). Their growth density is 7.5, 14, 4.7, 3.6 grasses m-2 in a severe case, respectively. The degree of grassland coverage had been decreased by 30~50%, compared with the 1950s in Alasan Left League. 96.9% of its grassland is desert or semi-desert (Fan et al., 2006). About 0.3 million sheep are poisoned and 0.03 million die every year.
SW, namely 1, 2, 8-trihydroxy indolizidine alkaloid, is the primary toxic component in locoweeds (Dorling et al., 1980; Molyneux and James, 1982). It was first isolated and identified from Swainsona canescens in 1979 (Colegate et al., 1979) and then from the locoweed A. lentiginosus in 1982 (Molyneux and James, 1982). Colegate et al. (1979) first let the aqueous fraction flow through the strongly acidic cation exchange resin and collected the component eluted by weak aqua ammonia. The component was concentrated and gradient eluted by aqueous sodium chloride through medicinal agarose gel CL-6B column, which was then extracted in ammonia chloroform, until finally a white needle crystal was acquired through crystallization and recrystallisation using chloroform and diethyl ether after recovering the solvent. Molyneux et al. (1991) extracted a white sera solid from the dry powder of the aerial part of spotted locoweed (Astragalus lentiginosus) by using acetone which was heated then cooled. The aqueous fraction of the solid was purified with a column of cation exchange resin AG50W2X8, during which some alkaline matter was recovered. The alkaline matter was detected to contain two components through TLC detection, both of which showed positive in the reaction with an alkaloids-specific reagent. The two components were isolated in chloroform and the undissolved part was precipitated in the form of white needle crystal, which was identified as SW through TLC, MS and NMR detection. Cao et al. (1989) isolated SW using cation exchange column. Li et al. (2005) acquired SW through ultrasonic oscillation, extracting in n-butanol and chloroform and sublimation. This method is easier and the extraction rate is quite high. In this study, the plant sample was thermal refluxed in 95% ETOH, the extraction was dissolved in 1% aqueous HCl and extracted in chloroform, ethyl acetate and n-butyl alcohol successively. The n-butanol extraction was then eluted by the eluant (chloroform:methanol: ammonia:water = 70:26:2:2) in the column. When the similar components, which had been detected by TLC detection, were combined and concentrated, SW was finally isolated. The extraction rate was 0.002%. The paste-forming rate was high but the sample should have strong resistance to high temperature.
The symptoms of nosotoxicosis caused by O. glabra are extremely similar to that caused by locoweed. After ingesting it, livestock show chronic symptoms such as loss of weight and rough hair, neurological symptoms such as astasia, tetraplegia, abortion and death in the most severe cases. Thus it has been assumed that SW is responsible for the poisonous effects of O. glabra. Toxic components of O. glabra were detected and isolated from the quinolizidine alkaloids, such as thermopsine, anagyrine, sperteine but the indolizidine alkaloids remained unidentified (Yang et al., 1989; Yu et al., 1991). The histopathological lesions caused by quinolizidine alkaloids are characterized by desmoplasia in liver, swelling and degeneration in fibers of myocardium and skeletal muscle instead of neurological symptoms. According to these symptoms, it’s assumed that quinolizidine alkaloids are not the main toxic ingredient of O. glabra. In this experiment, we have detected SW in chloroform extractum, ethyl acetate extractum and n-butanol extractum from O. glabra using TLC and GC detection, among which the SW content in n-butanol extractum was found to be the highest. We isolated a white needle crystal from the gel column chromatograph from n-butanol extractum, containing SW detected by TLC detection and subliming; we identified this compound to be SW through IR and MS analysis. By referencing previous reports on locoweed poisoning, we demonstrated that the main toxic component of O. glabra is indolizidine alkaloids instead of quinolizidine alkaloids.
SW can cause an accumulation of oligosaccharides and result in significant cellular vacuolation in parenchymatous organs (Elbein et al., 1981; Tulsiani et al., 1990; Stegelmeier et al., 2005). A previous study on the toxicity of O. glabra in mice (Chen et al., 2004) has shown a disordered activity of the CNS and cellular vacuolation in most organs, especially brains, after ingestion of this poisonous plant. This is similar to SW toxicity. In this study, SW was isolated from O. glabra, thus, it can be inferred that the principal toxin of this plant responsible for its toxicity is SW. SW can also be used as an immune modulator, inhibitor of neoplasm metastasis, antiviral drug and cellular protector. Moreover, SW can also promote the proliferation of bone marrow, as well as reduce the inhibition of bone marrow and neutrophilopenia caused by high-dose chemotherapy or lethal irradiation and it has been demonstrated that SW has a therapeutic effect on malignant tumor in humans, which makes it a promising drug in the medical area (Lu et al., 2009).
ACKNOWLEDGMENTS
This study was co-financed by the grants from the National Natural Science Foundation (No.31072175) and the Ph.D. Programs Foundation of Ministry of Education of China (No.20100204120018) and the Basic scientific research project of Northwest A and F University (No.QN2011108).
REFERENCES
- Boliang, D., W. Jianchen and X. Dengmin, 1995. A pathological study on ovary and placenta in dairy goats intoxicated through grazing Oxytropis kansuensis. Chin. J. Vet. Sci., 15: 27-32.
Direct Link - Dorling, P.R., C.R. Huxtable and S.M. Colegate, 1980. Inhibition of lysosomal α-mannosidase by swainsonine, an indolizidine alkaloid isolated from Swainsona canescens. Biochem. J., 191: 649-651.
Direct Link - Elbein, A.D., R. Solf, P.R. Dorling and K. Vosbeck, 1981. Swainsonine: An inhibitor of glycoprotein processing. Proc. Natl. Acad. Sci. USA., 78: 7393-7397.
PubMedDirect Link - Fan, Z.F., J.J. Wang, B.Y. Zhao, Q. Dong and N.T. Da, 2006. Locoweed harm investigation in Alasan League of Inner Mongolia. Chin. J. Grassland, 28: 56-59.
Direct Link - Martyn, A., J. Tyler, C. Offord and R. McConchie, 2003. Swainsona sejuncta: A species of ornamental promise or a potential weed? Aust. J. Exp. Agric., 43: 1369-1381.
CrossRefDirect Link - Medeiros, R.M.T., R.C. Barbosa, F. Riet-Correa, E.F. Limaa and I.M. Tabosaa et al., 2003. Tremorgenic syndrome in goats caused by Ipomoea asarifolia in Northeastern Brazil. Toxicon, 41: 933-935.
CrossRefDirect Link - Molyneux, R.J. and L.F. James, 1982. Loco intoxication: Indolizidine alkaloids of spotted locoweed (Astragalus lentiginosus). Science, 216: 190-191.
PubMed - Molyneux, R.J., L.F. James, K.E. Panter and M.H. Ralphs, 1991. Analysis and distribution of swainsonine and related polyhydroxyindolizidine alkaloids by thin-layer chromatography. Phytochem. Anal., 2: 125-129.
CrossRefDirect Link - Stegelmeier, B.L., L.F. James, D.R. Gardner, K.E. Panter and S.T. Lee et al., 2005. Locoweed (Oxytropis sericea)-induced lesions in mule deer (Odocoileius hemionus). Vet. Pathol., 42: 566-578.
PubMed - Taylor, J.B. and J.R. Strickland, 2002. Appearance and disappearance of swainsonine in serum and milk of lactating ruminants with nursing young following a single dose exposure to swainsonine (locoweed; Oxytropis sericea). J. Anim. Sci., 80: 2476-2484.
PubMed