
Abdolali Movahedinia
Department of Marine Biology, Faculty of Marine Sciences, Khorramshahr University of Marine Science and Technology, G.C., Khorramshahr, P.O. Box 669, Iran
Behrooz Abtahi
Department of Biology, School of Biological Sciences, Shahid Beheshti University, G.C., Tehran, Iran
Mahmood Bahmani
International Sturgeon Research Institute, Rasht, Iran
Asian Journal of Animal and Veterinary Advances
Year: 2012 | Volume: 7 | Issue: 8 | Page No.: 710-717







ABSTRACT
The present study, considered histopathological changes in the two fish species, Persian sturgeon Acipenser persicus and Stellate sturgeon Acipenser stellatus. The aim was to describe the different gill tissue changes in these two species of sturgeons. The prepared sections of gill were stained with haematoxylin and eosin (HE) and observed under a light microscope. According to the results of this case study, the important histological lesions observed in the gills of sturgeons were: (1) hyperplasia of secondary lamellar epithelium (2) epithelial lifting of secondary lamellae (3) leukocyte infiltration (4) hypertrophy of the respiratory epithelium (5) fusion of adjacent secondary lamellae and (6) lamellar aneurysm. Hyperplasia of secondary lamellar epithelium was the most frequent gill lesion and fusion of adjacent secondary lamellae as a result of hyperplasia, proliferation of epithelial cells or epithelial lifting and in some cases hypertrophy was the second frequent gill lesion.
PDF Abstract XML References Citation
Received: November 05, 2011;
Accepted: March 06, 2012;
Published: May 10, 2012
How to cite this article
Abdolali Movahedinia, Behrooz Abtahi and Mahmood Bahmani, 2012. Gill Histopathological Lesions of the Sturgeons. Asian Journal of Animal and Veterinary Advances, 7: 710-717.
DOI: 10.3923/ajava.2012.710.717
URL: https://scialert.net/abstract/?doi=ajava.2012.710.717
DOI: 10.3923/ajava.2012.710.717
URL: https://scialert.net/abstract/?doi=ajava.2012.710.717
INTRODUCTION
The gills consist of four branchial arches each bearing pairs of primary filaments upon which rows of secondary lamellae are situated. The filaments and lamellae are covered by epithelial cells (i.e., pavement cell, chloride cell (or Mitochondria rich cell) and mucous cell) supported by a complex system of blood vessels. Hence, the gill epithelia provide an extensive surface of contact with the environment (Wong and Wong, 2000; Mir and Channa, 2009; Movahedinia et al., 2009). The fish gill is a multifunctional organ involved not only in respiration but also in a variety of homeostatic activities such as osmoregulation, metabolism of circulating hormones, nitrogen excretion, ionic transport and acid-base balance (Haaparanta et al., 1997; Wood, 2001; Evans et al., 2005; Movahedinia et al., 2009). Fish gills typically respond rapidly to various irritants resulting gill lesions have a negative effect on the performance of fish and enhance its susceptibility to secondary diseases and potentially cause fish mortality (Haaparanta et al., 1997; Al-Attar, 2007).
A variety of factors, including environmental pollutants and many parasites can induce morphological anomalies in fish gills. Unfortunately, different irritants may cause almost identical lesion and gill structural damage may merely reflect a generalized stress response rather than toxicant, petroleum hydrocarbons or parasite-specific responses (Mallatt, 1985; Haaparanta et al., 1997; Osman et al., 2009).
Most studies of gill alterations have focused on farmed fish or fish from laboratory studies. Farmed fish are usually held under conditions favoring invasion by microbes and parasites, whereas laboratory studies often use concentrations of irritants orders of magnitude higher than those that occur in nature. Thus, laboratory tests do not reproduce real environmental conditions and may under-or overestimate effects and the responses of these fish may be exaggerated (Haaparanta et al., 1997; Alberto et al., 2005). Investigation on responses of natural populations of wild fish species would be desirable.
The present study described histopathological alterations of gills in two commercially, biologically and ecologically important fish species (that present only in the Caspian sea), Persian sturgeon, Acipenser persicus and Stellate sturgeon, A. stellatus.
MATERIALS AND METHODS
The samples ranging from 42.5 to 94.3 cm (n = 57) were collected from fishing ground along the southern coasts of Caspian Sea (Arjan (n = 12), Esmaili (n = 15), Kiashahr (n = 11) and Caviar Processing Centre (n = 19) in Gilan and Golestan provinces) (Fig. 1). For each fish, gill sections derived from the second arch of the gills on the right hand side of the head (Movahedinia et al., 2009) were taken in the field, at the collecting places and fixed in 10% formalin (pH = 7.2-7.4) for 24 h. After serial dehydration in ascending percentages of ethanol, the gills were infiltrated with xylene and embedded in paraffin. For each fish, two paraffin blocks were used. From each block, a set of serial sections (5-6 μm of thickness) were cut, parallel to the long axis of the filament. Sections (two slides from each block and two blocks per individual) were stained with haematoxylin and eosin (H and E) (Haaparanta et al., 1997). The prepared sections of gill were observed on a Nikon (Tokyo, Japan) light microscope. Slide preparations and microscopic evaluations were conducted in the laboratories of both International Sturgeon Research Institute (Rasht, Iran) and Tarbiat Modarres University (Noor, Iran), respectively. Different histological anomalies were investigated and photos were taken from some of them. Those lesions that rarely observed in slides were not described.
 | |
| Fig. 1: | Sturgeons sampling areas, southern Caspian sea (Gilan and Golestan provinces) |
RESULTS
Most gill changes were systemic and without any signs of causative agent. Hyperplasia of the lamellar epithelium was the most frequent observed gill lesion in both species (A. persicus and A. stellatus). The increase of cellular layers of the lamellar epithelium (Fig. 2) and also diffusion of distance between blood and aquatic medium was observed. Epithelial lifting of the secondary lamellae because of an infiltration of fluid made oedematous space between epithelium and basement membrane (Fig. 3). Similar to hyperplasia, this kind of lesion inhibited the respiratory gas exchange by increasing diffusion distance and decreasing interlamellar distance. There were blood cells (leukocytes) in leukocyte infiltration within the oedematous space (Fig. 4). The other observed lesion was Hypertrophy of the respiratory epithelium which referred to the numerous swollen epithelial cells related to the volume regulatory processes disorders, degeneration of cells in different phases, etc. (Fig. 5).
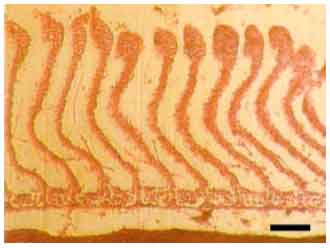 | |
| Fig. 2: | Distal hyperplasia of the secondary lamellar epithelium (A. persicus), The epithelial cell proliferation occurred preferentially at tip of lamella, Scale bar 100 μm |
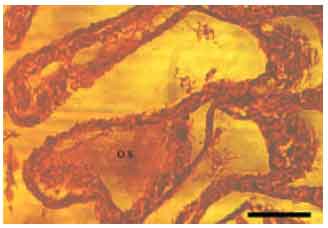 | |
| Fig. 3: | Epithelial lifting of secondary lamellae (A. stellatus), A space (named oedematous space (O.S.) growth between epithelial cells and beneath tissues (pillar cells and blood capillaries), Scale bar 50 μm |
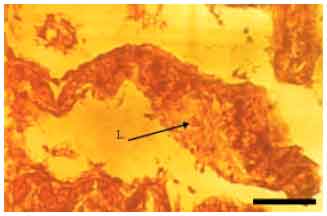 | |
| Fig. 4: | Leukocyte infiltration (A. stellatus), L.: Leukocyte, Scale bar 50 μm |
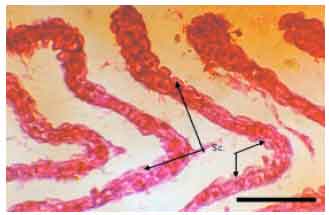 | |
| Fig. 5: | Hypertrophy of the respiratory epithelium (A. persicus), Sc.: Swollen cell, Scale bar 100 μm |
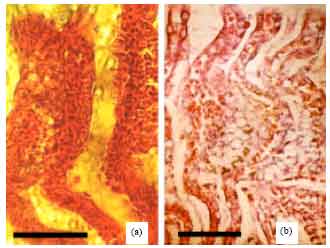 | |
| Fig. 6(a-b): | Fusion of adjacent secondary lamellae (a) Hyperplasia of the epithelial cells at tips caused fusion of the adjacent secondary lamellae (A. stellatus), Lamellae are free in the middle and base areas, Scale bar 50 μm, (b) Advanced cell proliferations and mucous secretion lead to fusion of secondary lamellae (A. persicus), Scale bar 50 μm |
Fusion of adjacent secondary lamellae (Fig. 6) was the second frequent gill lesion in the studied species. This was a result of hyperplasia of epithelial cells, lamellar epithelial lifting and mucous cell proliferation or in some cases hypertrophy of epithelial cells.
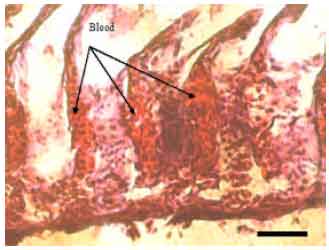 | |
| Fig. 7: | Lamellar aneurysm (A. stellatus), Mucous secreted in a large amount too and occupied the interlamellar space, Scale bar 100 μm |
Lamellar aneurysm characterized by accumulation of the blood cells in the secondary lamellae (Fig. 7). All explained lesions were observed in both A. persicus and A. stellatus.
DISCUSSION
The histopathological changes observed in the gills of Acipenser persicus and A. stellatus have been previously described in other wild fish (Laurent and Perry, 1991; Haaparanta et al., 1997; Wood, 2001; Handy et al., 2002) or exposed to different pollutants (Arellano et al., 1999; Van den Heuvel et al., 2000; De Boeck et al., 2001; Thophon et al., 2003; Pane et al., 2004; Monteiro et al., 2008; Abdel-Moneim et al., 2008; Bais and Lokhande, 2012). In these earlier studies, no clear relationship was found between the different histopathological changes nor the prevalence of lesions and the exposure of animals to a specific pollutant at the different concentrations and times of exposure (Monteiro et al., 2008). The sturgeons studied in the present study were usually caught when they were several years old and they were exposed to various irritants during their lifetime. It is known that hyperplasia is a lesion that appears as a response to many different irritants. As it is expected, the hyperplasia was the most frequent lesion found in the gill tissue of these fishes (both species from each area).
Copious mucous secretion of the gills with hyperplasia and hypertrophy of mucous-producing epithelial cells were widely reported as histopathological effects of toxicant such as crude oils and their products. The epithelial lifting and fusion of the secondary lamellae were previously described as the structural effects of petroleum hydrocarbons on marine fish, too. It was mentioned that fusion of adjacent secondary lamellae was frequent because some anomalies like hyperplasia and proliferation of epithelial cells in the acute phase resulted in lamellar fusion. This anomaly (fusion) may be parasitic (if only two or three neighboring lamellae fuse together) or systemic (if the fusion occurred in more vast areas). The hyperplasia, hypertrophy and fusion of adjacent lamellae appear to be a protective response to toxicants that causes less load of the toxicant to the blood (Kantham and Richards, 1995; Singhadach et al., 2009).
The appearance of extended lamellar oedematous spaces, sometimes containing infiltrated cells, seems to implicate a protective and osmoregulatory manifestation of the gills. The infiltration of eosinophilic granular cells and other mononuclear granular cells into the gill epithelium has been described as an inflammatory reaction by various workers in the response to different toxic chemical substances (Kantham and Richards, 1995; Authman and Abbas, 2007). These granular cells might have an antitoxic effect although the exact mechanism involved is unknown.
The epithelial lifting appears to be one of the initial reactions of the gill to a variety of pollutants (Smart, 1976) and previously described as a frequent gill lesion of fish exposed to toxicants (Mallatt, 1985; Arellano et al., 1999; Pane et al., 2004; Schwaiger et al., 2004; Authman and Abbas, 2007; Monteiro et al., 2008; Singhadach et al., 2009). It may act as a protective measure by increasing the pollutant-blood diffusion distance. Such changes are typical of an acute inflammatory response.
Marked dilation of the blood spaces in the secondary lamellae is an interesting histological feature. The decrease of branchial vascular resistance was accompanied by increases in the heart rate and blood pressure suggesting an increased rate of blood flow through the gills (Smart, 1976). Aneurysms were previously observed using a variety of toxicants and their severity was demonstrated to vary with pollutant levels and exposure times (Van den Heuvel et al., 2000; Thophon et al., 2003; Schwaiger et al., 2004; Al-Attar, 2007; Bais and Lokhande, 2012). Aneurysms might be used as a sensitive and reliable biomarker of acute toxicant exposure e.g., copper (Monteiro et al., 2008). In the present study, aneurysms were found in a few slides.
Numerous studies have indicated that the stress plays a significant role in increasing the fish susceptibility to diseases. The factors such as overcrowding, handling and poor water quality constitute various types of stress (Smart, 1976; Osman et al., 2009). The histopathological changes in the gill structure which occurred after prolonged exposure to toxicants may seriously reduce the oxygen diffusing capacity of the gills so that death may result primarily from insufficient oxygen uptake. Alternatively, the fish may be able to maintain their oxygen uptake over the long exposure period in the nature by compensatory cardiovascular and respiratory changes, though their activity may be curtailed.
CONCLUSION
In conclusion, the exposure of sturgeons to various biotic and abiotic stressors during several years through their life span can cause remarkable general anomalies especially the hyperplasia, hypertrophy and related lesions like fusion of adjacent lamellae. Although these histopathological alterations are protective responses at the initial phase but they can cause reduction of the fish gill capabilities and finally follow more acute diseases and death of fish. Observed and described histopathological lesions, were similar in both species.
ACKNOWLEDGMENTS
The authors are grateful to staff of the International Sturgeon Research Institute (Rasht, Iran) for help with sampling and field assistance and marine biology laboratory of Tarbiat Modares University (Noor, Iran) for providing technical assistance. The authors would like to thank Dr. Behrooz Heidari and Dr. Saeed Keyvanshokooh for their help and critical review of the manuscript.
REFERENCES
- Abdel-Moneim, A.M., N.M. Abou Shabana, S.E.M. Khadre and H.H. Abdel-Kader, 2008. Physiological and histopathological effects in catfish (Clarias lazera) exposed to dyestuff and chemical wastewater. Int. J. Zool. Res., 4: 189-202.
CrossRefDirect Link - Al-Attar, A.M., 2007. The influences of nickel exposure on selected physiological parameters and gill structure in the teleost fish, Oreochromis niloticus. J. Biol. Sci., 7: 77-85.
CrossRefDirect Link - Alberto, A., A.F.M. Camargo, J.R. Verani, O.F.T. Costa and M.N. Fernandes, 2005. Health variables and gill morphology in the tropical fish Astyanax fasciatus from a sewage-contaminated river. Ecotox. Environ. Safe, 61: 247-255.
CrossRef - Authman, M.M.N. and H.H.H. Abbas, 2007. Accumulation and distribution of copper and zinc in both water and some vital tissues of two fish species (Tilapia zillii and Mugil cephalus) of Lake Qarun, Fayoum Province, Egypt. Pak. J. Biol. Sci., 10: 2106-2122.
CrossRefPubMedDirect Link - Bais, U.E. and M.V. Lokhande, 2012. Effect of cadmium chloride on histopathological changes in the freshwater fish Ophiocephalus striatus (Channa). Int. J. Zool. Res., 8: 23-32.
CrossRefDirect Link - De Boeck, G., A. Vlaeminck, P.H. Balm, R.A. Lock, B. De Wachter and R. Blust, 2001. Morphological and metabolic changes in common carp, Cyprinus carpio, during short-term copper exposure: Interactions between Cu2+ and plasma cortisol elevation. Environ. Toxicol. Chem., 20: 374-381.
Direct Link - Evans, D.H., P.M. Piermarini and K.P. Choe, 2005. The multifunctional fish gill: Dominant site of gas exchange, osmoregulation, acid-base regulation and excretion of nitrogenous waste. Physiol. Rev., 85: 97-177.
CrossRefDirect Link - Haaparanta, A., E.T. Valtonen and R.W. Hoffman, 1997. Gill anomalies of perch and roach from four lakes differing in water quality. J. Fish Biol., 50: 575-591.
CrossRef - Handy, R.D., T. Runnalls and P.M. Russell, 2002. Histopathological biomarkers in three spined sticklebacks, Gasterosteus aculeatus, from several rivers in southern England that meet the freshwater fisheries directive. Ecotoxicology, 11: 467-479.
CrossRef - Kantham, K.P.L. and R.H. Richards, 1995. Effect of buffers on the gill structure of common carp, Cyprinus carpio L., and rainbow trout, Oncorhynchus mykiss (Walbaum). J. Fish Dis., 18: 411-423.
CrossRef - Laurent, P. and S.F. Perry, 1991. Environmental effects on fish gill morphology. Physiol. Zool., 64: 4-25.
Direct Link - Mallatt, J., 1985. Fish gill structural changes induced by toxicants and other irritants: A statistical review. Can. J. Fish. Aquat. Sci., 42: 630-648.
CrossRefDirect Link - Mir, I.H. and A. Channa, 2009. Gills of the snow trout, Schizothorax curvifrons Heckel: A SEM study. Pak. J. Biol. Sci., 12: 1511-1515.
CrossRefPubMedDirect Link - Monteiro, S.M., E. Rocha, A. Fontainhas-Fernandes and M. Sousa, 2008. Quantitative histopathology of Oreochromis niloticus gills after copper exposure. J. Fish Biol., 73: 1376-1392.
CrossRef - Movahedinia, A.A., A. Savari, H. Morovvati, P. Kochanian, J.G. Marammazi and M. Nafisi, 2009. The effects of changes in salinity on gill mitochondria-rich cells of juvenile yellowfin seabream, Acanthopagrus latus. J. Biol. Sci., 9: 710-720.
CrossRef - Osman, M.M., S.A. El-Fiky, Y.M. Soheir and A.I Abeer, 2009. Impact of water pollution on histopathological and electrophoretic characters of Oreochromis niloticus fish. Res. J. Environ. Toxicol., 3: 9-23.
CrossRefDirect Link - Pane, E.F., A. Haque and C.M. Wood, 2004. Mechanistic analysis of acute, Ni-induced respiratory toxicity in the rainbow trout (Oncorhynchus mykiss): An exclusively branchial phenomenon. Aquat. Toxicol., 69: 11-24.
CrossRef - Schwaiger, J., H. Ferling, U. Mallow, H. Wintermayr and R.D. Negele, 2004. Toxic effects of the non-steroidal anti-inflammatory drug diclofenac. Part I: Histopathological alterations and bioaccumulation in rainbow trout. Aquat. Toxicol., 68: 141-150.
CrossRef - Singhadach, P., W. Jiraungkoorsku, T. Tansatit, P. Kosa and C. Ariyasrijit, 2009. Calcium pre-exposure reducing histopathological alteration in Nile Tilapia (Oreochromis niloticus) after lead exposure. J. Fish. Aquat. Sci., 4: 228-237.
CrossRefDirect Link - Smart, G., 1976. The effect of ammonia exposure on gill structure of the rainbow trout (Salmo gairdneri). J. Fish. Biol., 8: 471-475.
CrossRefDirect Link - Van den Heuvel, M.R., M. Power, J. Richards, M. MacKinnon and D.G. Dixon, 2000. Disease and gill lesions in yellow perch (Perca flavescens) exposed to oil sands mining-associated water. Ecotox. Environ. Safe, 46: 334-341.
CrossRef - Wong, C.K.C. and M.H. Wong, 2000. Morphological and biochemical changes in the gills of Tilapia (Oreochromis mossambicus) to ambient cadmium exposure. Aquat. Toxicol., 48: 517-527.
Direct Link - Thophon, S., M. Kruatrachue, E.S. Upatham, P. Pokethitiyook, S. Sahaphong and S. Jaritkhuan, 2003. Histopathological alterations of white seabass, Lates calcarifer, in acute and subchronic cadmium exposure. Environ. Pollut., 121: 307-320.
CrossRefDirect Link - Arellano, J.M., V. Storch and C. Sarasquete, 1999. Histological changes and copper accumulation in liver and gills of the Senegales sole, Solea senegalensis. Ecotoxicol. Environ. Saf., 44: 62-72.
CrossRefDirect Link