
X.F. Yang
College of Animal Science, South China Agricultural University, Guangzhou 510642, People`s Republic of China
Z.Y. Jiang
Ministry of Agriculture, Key Laboratory of Animal Nutrition and Feed Science (South China), State Key Laboratory of Livestock and Poultry Breeding, Institute of Animal Science, Guangdong Academy of Agricultural Sciences, Guangzhou 510640, Poeple`s Republic of China
X.Y. Ma
Ministry of Agriculture, Key Laboratory of Animal Nutrition and Feed Science (South China), State Key Laboratory of Livestock and Poultry Breeding, Institute of Animal Science, Guangdong Academy of Agricultural Sciences, Guangzhou 510640, Poeple`s Republic of China
C.T. Zheng
Ministry of Agriculture, Key Laboratory of Animal Nutrition and Feed Science (South China), State Key Laboratory of Livestock and Poultry Breeding, Institute of Animal Science, Guangdong Academy of Agricultural Sciences, Guangzhou 510640, Poeple`s Republic of China
Y.C. Lin
Ministry of Agriculture, Key Laboratory of Animal Nutrition and Feed Science (South China), State Key Laboratory of Livestock and Poultry Breeding, Institute of Animal Science, Guangdong Academy of Agricultural Sciences, Guangzhou 510640, Poeple`s Republic of China
S.K. Wang
Ministry of Agriculture, Key Laboratory of Animal Nutrition and Feed Science (South China), State Key Laboratory of Livestock and Poultry Breeding, Institute of Animal Science, Guangdong Academy of Agricultural Sciences, Guangzhou 510640, Poeple`s Republic of China
S.Q. Jiang
Ministry of Agriculture, Key Laboratory of Animal Nutrition and Feed Science (South China), State Key Laboratory of Livestock and Poultry Breeding, Institute of Animal Science, Guangdong Academy of Agricultural Sciences, Guangzhou 510640, Poeple`s Republic of China
F. Chen
Ministry of Agriculture, Key Laboratory of Animal Nutrition and Feed Science (South China), State Key Laboratory of Livestock and Poultry Breeding, Institute of Animal Science, Guangdong Academy of Agricultural Sciences, Guangzhou 510640, Poeple`s Republic of China
G.L. Zhou
Ministry of Agriculture, Key Laboratory of Animal Nutrition and Feed Science (South China), State Key Laboratory of Livestock and Poultry Breeding, Institute of Animal Science, Guangdong Academy of Agricultural Sciences, Guangzhou 510640, Poeple`s Republic of China
Asian Journal of Animal and Veterinary Advances
Year: 2012 | Volume: 7 | Issue: 5 | Page No.: 412-419







ABSTRACT
The pork industry is dedicating to provide fast-growing pigs with high-grade meat; this study aimed to explore the relationship between dietary energy and growth output. Thirty two individually-penned male Lantang pigs (initially weighed 20.21±0.63 kg) were randomly assigned to one of four dietary treatments, with 4 replicates of 2 pigs each. The Digestive Energy (DE) of T (treatment) 1~4 were 9.24, 11.02, 12.66 and 14.29 MJ kg-1 feed, respectively. Feeding trail was terminated when all pigs within a treatment averaged 60±2 kg b.wt. and all pigs were slaughtered. The feeding time to reach terminal b.wt. were longer in both T1 and T2 in contrast with T3 and T4 (p<0.01). Average daily energy intake increased linearly with dietary energy concentration (p<0.01). Compared with the other three groups, average daily gain (p<0.05) and feed to gain ratio (p<0.05) of T4 were significantly increased and decreased, respectively. Relative contents of major white fat and perinephric fat, as well as adipocyte size of T4 were higher than those of the other groups (p<0.05). These results indicated that increasing dietary energy content improved fat accumulation in Lantang growing pigs possibly through direct deposition of fat in adipocyte. Higher content of dietary energy tend to increase growth rate of the pig, yet the over-deposition of body fat as well as its possible impact on carcass quality and marketing profit need to be further explored.
PDF Abstract XML References Citation
Received: September 28, 2011;
Accepted: January 02, 2012;
Published: February 17, 2012
How to cite this article
X.F. Yang, Z.Y. Jiang, X.Y. Ma, C.T. Zheng, Y.C. Lin, S.K. Wang, S.Q. Jiang, F. Chen and G.L. Zhou, 2012. Effect of Dietary Energy on Growth Performance and Fat Deposition of Lantang Growing Pigs. Asian Journal of Animal and Veterinary Advances, 7: 412-419.
DOI: 10.3923/ajava.2012.412.419
URL: https://scialert.net/abstract/?doi=ajava.2012.412.419
DOI: 10.3923/ajava.2012.412.419
URL: https://scialert.net/abstract/?doi=ajava.2012.412.419
INTRODUCTION
The pork industry has made great efforts in increasing growth rate and reducing fat content in pigs (Li et al., 2010; Realini et al., 2010). However, this may lead to reduction in fat quality and contrarily affect meat flavor, further processing and storage stability (Gatlin et al., 2002). Existing reports regarding the correlation between dietary energy and biological outcomes have frequently been conducted on the controlled basis (Campbell and Taverner, 1988; Bikker, 1994). Evidence showed that, controlled feed intake generally led to predictable and even consistent outcomes of changing dietary energy (Bikker, 1994; Beaulieu et al., 2009). Energy intake on an ad libitum basis and the growth performance of pigs are dependent upon animals and environmental factors such as feed energy density (Noblet and Van Milgen, 2004). To determine the optimal content of dietary energy for pigs which are often highly diverse, it is essential to probe energy utilization in pigs and the effect of changing dietary energy concentration on growth and carcass composition of the pig (Beaulieu et al., 2009).
The Lantang pig is an indigenous breed (fat-type) of South China which prone to get sexually matures and become fat. This traditional breed grows much more slowly and their carcass contained more fat and less lean than modern breeds (such as Duroc and Large White) which makes the breed a desirable animal model for researches of lipid metabolism and deposition in pigs (Chen et al., 2010). What’s worth mentioning, the nutrient requirements of Lantang pig, including dietary energy level, are hitherto poorly reported either at home or abroad. This study aimed to explore the effect of dietary energy concentration on carcass composition and fat deposition of Lantang growing pigs and to provide reference for the determination of energy requirement of the pig.
MATERIALS AND METHODS
Experimental materials and procedures: Experimental procedures for this study were approved by Animal Experimental Committee of Guangdong Institute of Animal Science. The study was conducted from December, 2009 to March, 2010, in Guangdong Institute of Animal Science.
Thirty two castrated male Lantang pigs with similar parity and birth date (initial body weight (BW) = 20.21±0.63 kg) were obtained from Heyuan Lantang-pig Conservation Farm, Guangdong province. Pigs were randomly assigned to one of four dietary treatments, with 4 replicates of 2 pigs each. The Digestive Energy (DE) of T1~T4 were 9.24, 11.02, 12.66 and 14.29 MJ kg-1 feed, respectively. The ingredients of all diets were identical (main ingredients were corn, soybean meal and wheat bran), although in different proportions. Concentrations of protein, essential amino acids and other nutrient components in all diets were adjusted to meet required NRC 1998 minimal levels (Table 1). Throughout the research, pigs were kept in individual pens with feed and water provided ad libitum. Pigs were weighed weekly to determine the weight gain. Feed delivery was recorded daily and feed remaining in the feeder was weighed weekly to determine feed intake and feed efficiency. The feeding trial was terminated when pigs within the same group averaged about 60±2 kg b.wt. Pigs terminated were weighed and their Backfat Thickness (BFT) at the 1st, 10th and last ribs were determined using an ultrasonic instrument (Renco, USA) before slaughter. All pigs were killed for determination of body composition and tissue sampling.
Analysis: Pigs were electrically stunned, exsanguinated, hair-removed and processed following commercial procedures (Savell and Smith, 2000). The eviscerated carcass was split longitudinally through the vertebrae midline and the left side was weighed. The left-side carcass was dissected into lean mass, subcutaneous and intermuscular fat, bone and cuticle following the procedure of Walstra and Merkus (1995). Individual weight of the above tissues was measured and their respective proportions to the whole carcass were calculated. Dorsal Subcutaneous Adipose Tissue (SAT) and Longissimus Dorsi Muscle (LDM) between 4~5 lumbar vertebra were obtained from the right-side carcass and immediately stored in 10% formaldehyde solution for histological study.
Tissues for histological research were processed as described by Touchette et al. (2002).
| Table 1: | Ingredients and nutrient composition of research diets fed to Lantang growing pigs, % as-fed basis |
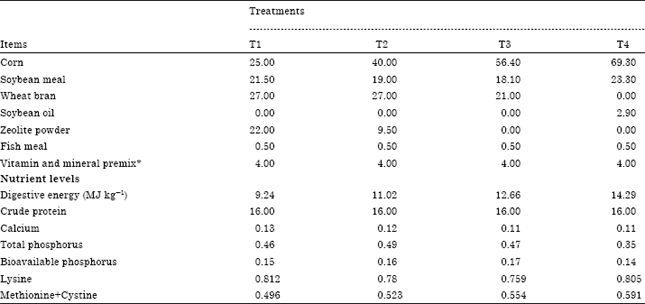 | |
| *Provided the following nutrients per kilogram of diet (mg); Retinol: 6.4; Cholecalciferol: 0.083; α-tocopherol: 22; Menadione: 0.60; Riboflavin: 3.3; Nicotinic acid: 16.5; Pantothenic acid: 5.5; Pyrodoxine: 1.1; Biotin: 0.56; Choline: 1100; Cyanocobalamin: 0.017; Fe: 88; Zn: 55; Mn: 22; Cu: 6.6; I: 0.22; Se: 0.1 | |
Briefly, SAT and LDM samples were stored in 10% neutral buffered formalin for 24 h, after which they were removed out of the buffer and processed in low-melt paraffin and stained with hematoxylin and eosin. The diameter and amount of adipocyte or muscle fiber were measured.
Statistical analysis: Data were expressed as Mean±SEM. Comparisons between groups were done by using GLM procedure of SAS 9.2 (SAS Institute, Cary, NC, USA) and a p-value<0.05 was considered significant.
RESULTS AND DISCUSSION
There was no significant difference among the four treatments in initial BW or final BW of the pigs (Table 2). However, to reach the similar final BW, the feeding time was markedly extended in both T1 and T2 (100 days) in contrast with T3 and T4 (72 days) (p<0.01). As previously reported, increasing dietary energy concentration for growing pigs typically improves Average Daily Gain (ADG) and gain: feed ratio (Pettigrew and Moser, 1991). Beaulieu et al. (2009) also reported that ADG, feed efficiency and daily caloric intake of pigs was significantly improved by energy content of diet. However, variability in growth was not affected by dietary energy treatment (Beaulieu et al., 2009). In this study, the overall ADG increased with dietary energy concentrations. ADG of T4 was notably higher than those of T1 and T2 (p<0.05), meanwhile, ADG of T3 was higher than T1 (p<0.05). Previous reports have assumed that growing pigs being imposed upon a wide range of dietary energy contents would adjust feed intake to maintain a fairly constant daily energy intake (Ellis and Augspurger, 2001). Black (1995) suggested that the critical lower limit of DE for pigs weighing between 20 to 50 kg would be 13.81 MJ kg-1 feed and another investigation indicated that growth of growing pigs would be normally performed at dietary DE concentrations above 14.55 MJ kg-1 (King, 1999). Nevertheless, the range of energy is poorly defined, especially in Lantang pigs. As this data revealed, the higher level of dietary energy resulted in greater ADG of pigs.
| Table 2: | Effect of dietary energy on growth performance of Lantang growing pigs |
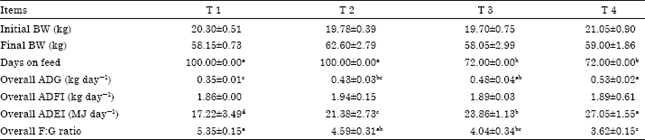 | |
| Values are Mean±SEM. T1, T2, T3 and T4 represent experimental treatments of dietary DE 9.24, 11.02, 12.66 and 14.29 MJ kg-1 feed, respectively. a,b,c Means in a row without a common letter differ, p<0.05. BW: Body weight; ADG: Average daily gain; ADFI: Average daily feed intake; ADEI: Average daily energy intake; F:G ratio: Feed to gain ratio | |
| Table 3: | Effect of dietary energy on BFT of Lantang growing pigs |
 | |
| Values are Mean±SEM. T1, T2, T3 and T4 represent experimental treatments of dietary DE 9.24, 11.02, 12.66 and 14.29 MJ kg-1 feed, respectively. a,b,cMeans in a row without a common letter differ, p<0.05. BFT: Back fat thickness | |
Adeniji (2008) found that decreasing dietary energy content might lead to the decrease of feed intake in pigs. However, Average Daily Feed Intake (ADFI) of pigs in this study was identical between different dietary treatments. Beaulieu et al. (2009) clarified that increasing energy density of the diet prominently improved energy intake. The overall average daily energy intake (ADEI, MJ of DE/day) in the present research increased remarkably with dietary energy concentration (linear, p<0.01). These results are also consistent with the finding of Williams et al. (1994) in which ADG of male pigs linearly increased with increasing energy intake. Nevertheless, report on chicken has showed negative effect of dietary energy on growth rate (Vahdatpour et al., 2008), the difference between the studies could be ascribe to the species difference of animals.
Adding fat/oil to the diets has previously been found to improve feed efficiency in pigs (Pettigrew and Moser, 1991). Gilts fed a fat-blended diet had an improved feed:gain ratio than gilts fed diet with no fat added (Realini et al., 2010). Increasing added dietary fat linearly increased ADG and gain to feed ratio of pigs during 36 and 59 kg b.wt. (Stahly et al., 1981; Campbell and Taverner, 1988). In this study, the overall feed to gain (F:G) ratio of T4 was 32.34% (p<0.05), 21.18% (p<0.05) and 10.45% (p>0.05) lower than those of T1, T2 and T3, respectively, showing that elevated dietary energy (attained by adding soybean oil) actually improved feed efficiency. These results were different from chicken study in which the optimal feed conversion rate was found in a lower dietary energy level rather than the higher levels (Mbajiorgu et al., 2011).
The increment of DE by supplementing fat to diets generally improves carcass fat content in growing-finishing pigs (Pettigrew and Moser, 1991). Reducing the proportion of protein:energy in the diet typically increases fat deposition (Wood et al., 2004). Likewise, higher level of dietary energy led to higher content of body fat in Japanese Quail (Parizadian et al., 2011). This study disclosed that, BFT at the 10th (p<0.05) and last (p<0.05) rib locations as well as the average BFT (p<0.05) were higher in T4 than T3 (Table 3), whereas, no obvious difference was noted among the other groups or rib locations.
| Table 4: | Effect of dietary energy on carcass composition of Lantang growing pigs |
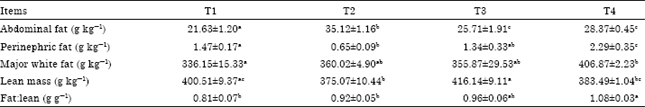 | |
| Values are Mean±SEM. T1, T2, T3 and T4 represent experimental treatments of dietary DE 9.24, 11.02, 12.66 and 14.29 MJ kg-1 feed, respectively. a,b,c Means in a row without a common letter differ, p<0.05 | |
| Table 5: | Effect of dietary energy on histological figures of Lantang growing pigs |
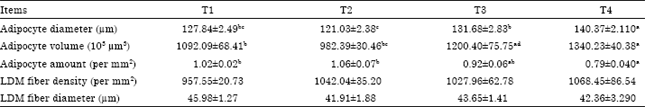 | |
| Values are Mean±SEM. T1, T2, T3 and T4 represent experimental treatments of dietary DE 9.24, 11.02, 12.66 and 14.29 MJ kg-1 feed, respectively. a,b,cMeans in a row without a common letter differ, p<0.05. LDM: Longissimus dorsi muscle | |
Previous studies showed that BFT differed markedly between high-fat and low-fat diet groups in genetically lean pigs (Pond et al., 1985), also, genetically obese pigs on high-fat diet were found to gain body-fat contents 4-fold higher than those of low-fat pigs (Diersen-Schade et al., 1985) which are consistent with our data.
BFT is positively correlated with direct measures of subcutaneous, retroperitoneal and visceral fat in pigs (Witczak et al., 2005; Dyson et al., 2006), thicker backfat directly suggested fatter signs in pigs. As this study shown, the relative abdominal fat content of T2 was greater than those of other groups (p<0.05) (Table 4); although, relative abdominal fat contents of T3 and T4 did not differ, both of the two groups were higher than T1 (p<0.05). The relative perinephric fat content of T4 was higher than the other groups (p<0.05) and that of T1 was higher than T2 (p<0.05). Compared with T1, the relative content of major white fat was higher in T4 (p<0.05). The relative lean mass of T3 was significantly higher than those of T2 and T4 (p<0.01) while that of T1 was significantly higher than T2 (p<0.05). In contrast with T4, the fat:lean ratio were markedly lower in both T1 (p<0.01) and T2 (p<0.05). Beaulieu et al. (2009) declared that elevating dietary energy content increased BFT while having no influence on carcass index and longissimus dorsi muscle thickness. This contradiction may be ascribe to the possible difference of lipid deposition and metabolism between the two dissimilar type of pigs (genetically lean pigs vs. genetically fat-type pig) used in the two studies.
The results of histological observation on SAT and LDM were shown in Table 5, Fig. 1 and 2. The SAT adipocyte diameter of pigs in T4 was higher than those of the other groups (p<0.05), likewise, adipocyte diameter of T3 was greater than T2 (p<0.01). SAT adipocyte volume of T4 was higher in contrast with T1 and T2 (p<0.01) and that of T3 was higher than T2 (p<0.05). Inversely, SAT adipocyte amount of T4 was lower than both T1 and T2 (p<0.01). No obvious difference was detected among the four groups in either LDM fiber density or LDM fiber diameter (Table 5, Fig. 2). Previous studies have shown that, body fat accumulation was elevated over high levels of dietary energy (attained mainly by supplementing fat in the diets) in both young and growing pigs (Frobish et al., 1970; Allee et al., 1971).
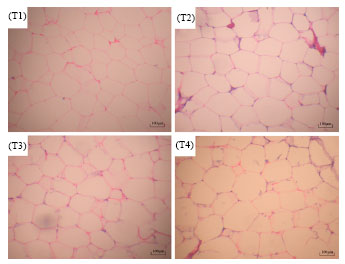 | |
| Fig. 1: | Hematoxylin and eosin staining of SAT adipocytes in Lantang growing pigs. T1, T2, T3 and T4 represent experimental treatments of dietary DE 9.24, 11.02, 12.66 and 14.29 MJ kg-1 feed, respectively. SAT: Dorsal subcutaneous adipose tissue |
 | |
| Fig. 2: | Hematoxylin and eosin staining of LDM fiber in Lantang growing pigs. T1, T2, T3 and T4 represent experimental treatments of dietary DE 9.24, 11.02, 12.66 and 14.29 MJ kg-1 feed, respectively. LDM: Longissimus dorsi muscle |
Indicated that elevating dietary fat (corn oil) concentration from 1-13% notably depressed (60-70% ) in vitro lipogenesis while increasing contents of carcass fat, presumably owing to the direct deposition of dietary fat which is in line with the results of adipocyte augmentation in this study.
CONCLUSION
Lantang growing pigs respond to elevated dietary energy levels by raising body fat deposition. Pigs under dietary DE level of 14.29 MJ kg-1 feed maintain the greatest growth rate, along with the highest relative content of body fat.
ACKNOWLEDGMENT
This study was supported by National Natural Science Foundation of China (30771562). We thank our staff fellows of Institute of Animal Science, Guangdong Academy of Agricultural Sciences, for technical assistance.
REFERENCES
- Adeniji, A.A., 2008. The feeding value of rumen content-maggot meal mixture in the diets of early weaned piglets. AJAVA., 3: 115-119.
CrossRefDirect Link - Parizadian, B., M.S. Shargh and S. Zerehdaran, 2011. Study the effects of different levels of energy and l-carnitine on meat quality and serum lipids of japanese quail. Asian J. Anim. Vet. Adv., 6: 944-952.
CrossRef - Diersen-Schade, D.A., M.J. Richard, D.C. Beitz and L.N. Jacobson, 1985. Effects of beef, soy and conventional diets on body composition and plasma lipids of young pigs fed restricted or liberal amounts of diet. J. Nutr., 115: 1016-1024.
PubMed - Allee, G.L., E.K. Ohea, G.A. Leveille and D.H. Baker, 1971. Influence of dietary protein and fat on lipogenesis and enzymatic activity in pig adipose tissue. J. Nutr., 101: 869-878.
Direct Link - Beaulieu, A.D., N.H. Williams and J.F. Patience, 2009. Response to dietary digestible energy concentration in growing pigs fed cereal grain-based diets. J. Anim. Sci., 87: 965-976.
CrossRef - Mbajiorgu, C.A., J.W. Ngambi and D. Norris, 2011. Effect of varying dietary energy to protein ratio level on growth and productivity of indigenous venda chickens. Asian J. Anim. Vet. Adv., 6: 344-352.
CrossRefDirect Link - Campbell, R.G. and M.R. Taverner, 1988. Genotype and sex effects on the relationship between energy intake and protein deposition in growing pigs. J. Anim. Sci., 66: 676-686.
Direct Link - Dyson, M.C., M. Alloosh, J.P. Vuchetich, E.A. Mokelke and M. Sturek, 2006. Components of metabolic syndrome and coronary artery disease in female Ossabaw swine fed excess atherogenic diet. Comp. Med., 56: 35-45.
PubMed - Frobish, L.T., V.W. Hays, V.C. Speer and R.C. Ewan, 1970. Effect of fat source and level on utilization of fat by young pigs. J. Anim. Sci., 30: 197-202.
Direct Link - Gatlin, L.A., M.T. See, J.A. Hansen, D. Sutton and J. Odle, 2002. The effects of dietary fat sources, levels and feeding intervals on pork fatty acid composition. J. Anim. Sci., 80: 1606-1615.
Direct Link - Li, D.Z., J.X. He, M.G. Lei, D.Q. Xu, S.W. Jiang and Y.Z. Xiong, 2010. Polymorphism in exon 2 of pig FIT1 gene and its association with fat-deposition-related traits. Yi Chuan, 32: 375-380.
PubMed - Noblet, J. and J. van Milgen, 2004. Energy value of pig feeds: Effect of pig body weight and energy evaluation system. J. Anim. Sci., 82: E229-E238.
Direct Link - Realini, C.E., P.D. Montge, R. Lizardo, M. Gispert, M.A. Oliver and E.E. Garcia, 2010. Effect of source of dietary fat on pig performance, carcass characteristic and carcass fat content, distribution and fatty acid composition. Meat Sci., 85: 606-612.
PubMed - Stahly, T.S., G.L. Cromwell and J.R. Overfield, 1981. Interactive effects of season of year and dietary fat supplementation, lysine source and lysine level on the performance of swine. J. Anim. Sci., 53: 1269-1277.
Direct Link - Touchette, K.J., J.A. Carroll, G.L. Allee, R.L. Matteri, C.J. Dyer, L.A. Beausang and M.E. Zannelli, 2002. Effect of spray-dried plasma and lipopolysaccharide exposure on weaned pigs: I. Effects on the immune axis of weaned pigs. J. Anim. Sci., 80: 494-501.
Direct Link - Vahdatpour, T., K. Nazer-Adl, Y. Ebrahim-Nezhad, N. Maheri-Sis and S. Vahdatpour, 2008. The effects of energy increasing and protein lowering by addition of fats to diet on broiler chickens: performance and serum lipids. AJAVA., 3: 286-292.
CrossRefDirect Link - Williams, N.H., T.R. Cline, A.P. Schinckel and D.J. Jones, 1994. The impact of ractopamine, energy intake and dietary fat on finisher pig growth performance and carcass merit. J. Anim. Sci., 72: 3152-3162.
Direct Link - Witczak, C.A., E.A. Mokelke, R. Boullion, J. Wenzel, D.H. Keisler and M. Sturek, 2005. Noninvasive measures of body fat percentage in male Yucatan swine. Comp. Med., 55: 445-451.
PubMed - Wood, J.D., G.R. Nute, R.I. Richardson, F.M. Whittington and O. Southwood et al., 2004. Effects of breed, diet and muscle on fat deposition and eating quality in pigs. Meat Sci., 67: 651-667.
CrossRef