
Abdul-Reza Dabbagh
Department of Marine Biology, Faculty of Sciences, University of Hormozgan, P.O. Box 3995, Bandar Abbas, Iran
Ehsan Kamrani
Department of Marine Biology, Faculty of Sciences, University of Hormozgan, P.O. Box 3995, Bandar Abbas, Iran
Mohammad Reza Taherizadeh
Department of Marine Biology, Faculty of Sciences, University of Hormozgan, P.O. Box 3995, Bandar Abbas, Iran
Asian Journal of Animal and Veterinary Advances
Year: 2012 | Volume: 7 | Issue: 4 | Page No.: 361-365







ABSTRACT
Alpheus lobidens De Haan, 1849, like many other snapping shrimp, is a socially monogamous species typically found in male-female pairs in burrows under rocks. We evaluated mate guarding (detection of female receptivity) and shared resource defense as factors that could have led to social monogamy in Iranian populations of this common intertidal species. Males within a pair did not move more pieces of gravel than females during burrow building. A shared resource defense experiment revealed that individuals within a pair did not differ significantly in their activity in repelling intruders.
PDF Abstract XML References Citation
Received: August 03, 2011;
Accepted: December 05, 2011;
Published: January 19, 2012
How to cite this article
Abdul-Reza Dabbagh, Ehsan Kamrani and Mohammad Reza Taherizadeh, 2012. Studies on Social Monogamy in the Snapping Shrimp, Alpheus lobidens De Haan, 1849. Asian Journal of Animal and Veterinary Advances, 7: 361-365.
DOI: 10.3923/ajava.2012.361.365
URL: https://scialert.net/abstract/?doi=ajava.2012.361.365
DOI: 10.3923/ajava.2012.361.365
URL: https://scialert.net/abstract/?doi=ajava.2012.361.365
INTRODUCTION
Social monogamy has been defined as a situation when two heterosexual adults share a territory or otherwise live together in a pair which may or may not also involve exclusive reproductive behaviour (Wickler and Seibt, 1981), whereas mate guarding refers to males defending their mates either before or after mating. Social monogamy has evolved in many taxa (Wittenberger and Tilson, 1980; Bull et al., 1998; Mathews, 2002a), including those that exhibit biparental care of offspring (e.g., tenebrionid beetles and many species of birds) and those that do not (e.g., caridean shrimp) (Joseph et al., 2005; Rahman et al., 2003). Social monogamy has been studied extensively in crustaceans and insects (e.g., Grafen and Ridley, 1983; Dunham et al., 1986; Jivoff and Hines, 1998; Mathews, 2002a, b; Rahman et al., 2003; Detto and Backwell, 2009) as well as in some vertebrates (Bull et al., 1998; Gillette et al., 2000; Joseph et al., 2005). Male mate guarding and shared territoriality is likely to guarantee access to a receptive female (Jormalainen, 1998; Rahman et al., 2003) and may also be favored if territorial defense or maintenance is helped by the presence of two individuals. Physiological factors, such as cryptic female fertility, may also favour mate guarding or social monogamy since males would have regular access to the female throughout the reproductive cycle. Ecological factors, such as the wide dispersal of potential mates, may also favour mate guarding since it would limit the (energetic) costs of mate search (Rahman et al., 2003).
In addition, species with mate guarding may share the defence of their shelter; this could reduce the risk of being evicted and therefore represent a substantial benefit to social monogamy (Rahman et al., 2003).
Several alpheid shrimp species have been studied with respect to their mating system which often involves male-female pairs living together (Knolton, 1980; Rahman et al., 2003; Mathews, 2002a, b). Receptivity in female alpheid shrimps is restricted to the period immediately after moulting which might favor social monogamy or mate guarding as advantageous to both males and females for the reasons outlined above (Rahman et al., 2003). The ability of males to detect receptive females should also be advantageous.
In socially monogamous snapping shrimp, females often act territorially against other females and thus, no harem formation is possible. Furthermore, males would improve their mating success by defending their mates against other males (Knolton, 1980). Mathews (2002a) suggests that social monogamy in Alpheus angulatus has been selected due to advantages of territorial cooperation and burrow construction.
In the present study, we studied factors that could lead to social monogamy in Alpheus lobidens De Haan, 1849. This species makes burrows in soft sediments under rocks. The burrows offer the owner protection and are defended from conspecifics.
In this project, we tested three hypotheses: (1) Males can detect female receptivity, (2) pairs share their duties in building a burrow and (3) burrows are defended by both members of the pair.
MATERIALS AND METHODS
Collection and shrimp maintenance: We collected Alpheus lobidens De Haan, 1849 in pairs and singly on a rocky shore next to the Station for Molluscan Research in Bandar-e Lengeh, Hormozgan province, Iran. The shrimp were kept in pairs or individually in 72 l aquaria with 21 separate compartments which were partitioned by screens, about 7 cm in width and 35 cm in height. Shrimp were kept at 27°C on a 14 h L : 10 h D illumination cycle and were fed commercial fish pellets on a daily basis. Checks for moulting were made twice daily. Body length was measured from the end of the tail to the tip of the rostrum and snapping claw length was measured as the maximum length of the propodus. A given shrimp was only used once in each experiment. The research was conducted in summer.
Detection of female receptivity: We tested whether males can distinguish between pre-moult and inter-moult stage females. Two females of equal size along with a male of equal size were simultaneously placed in an experimental tank 60 cm length x 30 cm width x 40 cm height. As soon as the shrimps were introduced into the tank, one opaque plastic shelter was placed in a corner of the test chamber. One female had moulted 2-4 days previously and the second female was 1-2 days premoult (estimated by our observations that female moulting occurs every 21 days, on average. Pairing over a 2 h period was recorded. Twenty-five replicates were conducted (25 males and 50 females in total).
Burrow building: We used infrared-sensitive time-lapse video to record daytime and nighttime activities of the shrimp to test the hypothesis that individuals in pairs share their duties in building a burrow. Before each test, we put cleaned substrate, 15 cm deep, taken from the shrimps’ natural habitat into the test aquarium (35 cm lengthx25 cm widthx15 cm height) and filled the aquarium with sea water to a depth of ~25 cm. We also placed a bivalve shell in the center of the test chamber, to provide a focus for burrow building.
First, we recorded data pertaining to burrow building. We monitored burrow building of test shrimp (n = 20 pairs) for 30 min per day on day 1 immediately after the shrimp were introduced to the test chamber and 30 min per night on night 1, 2 h after introduction, using infrared sensitive video. We counted the number of pieces of gravel moved by both sexes.
The response of a pair to an intruder: To determine whether the pair can jointly defend the shelter against shrimps of different sizes, the response of a pair to an intruder was tested in an experimental aquarium identical to that in which the pair existed formerly within a constructed burrow. Six different intruders types were tested: a larger female, a larger male, a smaller male, a smaller female, a same-size male and a same-size female. The responses of each pair were observed for 30 min immediately following introduction of the intruder and their behaviour was videotaped. The frequency of snaps was recorded from the tapes. After a further 1 day, the animals’ pairing status and location were recorded. No pair was used more than once in each response. Twenty replicate trials were conducted for each of the six different intruders types tested.
Statistical analysis: Statistical analyses for all of treatments used SPSS. The Chi-square test was used to distinguish the ability of the male to detect female receptivity and the role of size in defense against intruders. The Mann-Whitney rank sum test was used to compare males and females with regard to numbers of moved pieces of gravel and activity levels in repelling intruders.
RESULTS
Detection of female receptivity: Of the 25 males of Alpheus lobidens tested, 11 paired with the female that had recently completed its moult, 12 paired with the female that was to moult soon and 2 males did not pair with either female. Two pairings were formed out of a burrow. This indicates that males did not distinguish between females that were early or very late in their moulting cycle (chi-square test, χ2 = 1.06, df = 1, p = 0.19) or did not exhibit a preference based on moult cycle.
Burrow building: Individuals within a pair did not differ in their movement of pieces of gravel per 30 min during the daytime observation period (Mann-Whitney rank sum test, T = 181, p = 0.80) and or the nighttime observation period (Mann-Whitney rank sum test, T = 157, p = 0.35) (Table 1).
The response of a pair to an intruder: For both males and female intruders, similar patterns were observed. There were significant differences associated with different sized intruders (chi-square test, for males: χ2 = 6.58, df = 2, p = 0.03; for females: χ2 = 6.02, df = 2, p = 0.04) (Table 2). Pairs were never disrupted when the intruder was smaller. The most common outcome when the intruder was larger was the eviction of the same sexed member of the pair and the pairing of the intruder with the opposite sexed resident.
Finally, the shared resource defence experiment revealed that individuals within a pair did not significantly differ in activity in repelling intruders by snapping (Mann-Whitney rank sum test, T = 129.5, p = 0.08).
| Table 1: | Mean (± SD) frequency of building by day and night Alpheus lobidens De Haan |
 | |
| Table 2: | Effects of different types of intruders on paired residents Alpheus lobidens De Haan |
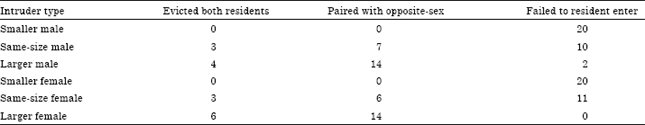 | |
| N = 20 for each intruder type which introduced with different pairs | |
DISCUSSION
Our study on the detection of female receptivity showed that males did not distinguish between females soon to be receptive and those not soon to be receptive or did not exhibit a preference based on moult cycle. One possible explanation is the fact that pair bonds may typically be of long duration and movement between burrows may be risky. A review of other studies of male mate guarding in mobile species by Rahman et al. (2003) has shown that recognition of receptivity commonly occurs but that the male typically forms a pair bond with a female for only a few days. Lack of recognition of female receptivity has been shown for other caridean shrimps such as Palaemonetes pugio Holthuis, 1949 (Bauer and Abdalla, 2001). It may also be the case that males simply preferred the female that was successful in defending the single burrow.
Cooperative burrow construction has been reported in many taxa (Mathews, 2002a). Cooperation in burrow building and construction usually will decrease investment in burrowing by each individual. Although, Mathews (2002a) showed that females Alpheus angulatus McClure, 1995 spent significantly more time in burrow construction than males, present results suggest that females are not more active in burrowing than males, perhaps because the both had the same role in defense.
In many taxa, shared territorial defence is associated with social monogamy (Wickler and Seibt, 1981) and both members of the mated pair usually defend the burrow (Wittenberger and Tilson, 1980). In Alpheus lobidens De Haan, 1950, as with other socially monogamous species such as Alpheus heterochaelis Say, 1818 (e.g., Rahman et al., 2003) the cost of defence is the snaps that are exchanged between intruder and resident, the risk of injury and the burden of maintaining a large weapon (the major chela). The fact that larger intruders often became the mates of opposite sexed residents suggests that shelter defence may be more important than maintaining the bond with a specific individual (Rahman et al., 2003).
ACKNOWLEDGMENTS
We thank Mahdi Dabbagh for providing us with snapping shrimp, Dr. Patricia R. Y. Backwell for providing us with articles and Dr. Arthur Anker for species identification. We also have special thanks to Dr. Nancy Knowlton for advice and comments on an earlier version of this manuscript.
REFERENCES
- Bauer, R.T. and J.H. Abdalla, 2001. Male mating tactics in the shrimp Palaemonetes pugio (Decapoda, Caridea): Precopulatory mate guarding vs. pure searching. Ethology, 107: 185-199.
CrossRefDirect Link - Bull, C.M., S.J.B. Cooper and B.C. Baghurst, 1998. Social monogamy and extra-pair fertilization in an Australian lizard, Tiliqua rugosa. Behav. Ecol. Sociobiol., 44: 63-72.
CrossRefDirect Link - Detto, T. and P.R.Y. Backwell, 2009. Social monogamy in a fiddler crab, Uca capricornis. J. Crustacean Biol., 29: 283-289.
CrossRefDirect Link - Dunham, P.J., T. Alexander and A. Hurshman, 1986. Precopulatory mate guarding in an amphipod, Gammarus lawrencianus bousfield. Anim. Behav., 34: 1680-1686.
CrossRefDirect Link - Jivoff, P. and A.P. Hines, 1998. Female behaviour, sexual competition and mate guarding in the blue crab, Callinectes sapidus. Animl. Behav., 55: 589-603.
CrossRefDirect Link - Gillette, J.R., R.G. Jaeger and M.G. Peterson, 2000. Social monogamy in a territorial salamander. Anim. Behav., 59: 1241-1250.
CrossRefDirect Link - Grafen, A. and M. Ridley, 1983. A model of mate guarding. J. Theor. Boil., 102: 549-567.
CrossRefDirect Link - Jormalainen, V., 1998. Precopulatory mate guarding in crustaceans: Male competitive strategy and intersexual conflict. Q. Rev. Biol., 73: 275-304.
Direct Link - Joseph, L.L., C.T. Cope, N.R. Kohn and R.G. Jaeger, 2005. The influence of familiarity on socially monogamous behavior on red-backed salamanders. Herpetoligica, 61: 343-348.
CrossRefDirect Link - Knolton, N., 1980. Sexual selection and dimorphism in two demes of a symbiotic, pair-bonding snapping shrimp. Evolution, 34: 161-173.
Direct Link - Mathews, L.M., 2002. Territorial cooperation and social monogamy: Factors affecting intersexual behaviours in pair-living snapping shrimp. Anim. Behav., 63: 767-777.
CrossRefDirect Link - Mathews, L.M., 2002. Tests of the mate-guarding hypothesis for social monogamy: Does population density, sex ratio, or female synchrony affect behaviour of male snapping shrimp (Alpheus angulatus)? Behav. Ecol. Sociobiol., 51: 426-432.
CrossRefDirect Link - Rahman,N., D.W. Dunham and C.K. Govind, 2003. Social monogamy in the big-clawed snapping shrimp, Alpheus heterochelis. Ethology, 109: 457-473.
CrossRefDirect Link - Wickler, W. and U. Seibt, 1981. Monogamy in crustacea and man. Zeitschrift Tierpsychol., 57: 215-234.
CrossRefDirect Link - Wittenberger, J.F. and R.L. Tilson, 1980. The evolution of monogamy: Hypothesis and evidence. Ann. Rev. Ecol., 11: 197-232.
CrossRefDirect Link