
Alaeldein M. Abudabos
Department of Animal Production, College of Food and Agricultural Sciences, King Saud University, P.O. Box 2460, Riyadh 11451, Kingdom of Saudi Arabia
LiveDNA: 966.4788
ORCID: 0000-0001-7998-0344
Asian Journal of Animal and Veterinary Advances
Year: 2012 | Volume: 7 | Issue: 1 | Page No.: 20-29







ABSTRACT
Due to environmental and economic concerns there is a great interest in providing adequate Phosphorus (P) level to sustain performance of poultry while reducing P excretion with manure to the environment. The purpose of the present study was to determine the effect of dietary Total Phosphorus (TP) in birds sorted into 2 groups: Heavy (H) and Light (L) from three to seven weeks of age on performance, serum phosphorus (serum P) and intestinal phytase activity. From three to six weeks of age, diets contained 5.0, 5.5, 6.0 and 6.5 g TP kg-1. From six to seven weeks, birds received diets that contained 3.8, 5.5, 6.0 and 6.5 g TP kg-1. At six and seven weeks of age interactions were not detected and either dietary TP or weight group had a significant effect on performance. Serum P was affected by the amount of inorganic phosphorus in the diet. Phytase activity was dramatically affected by diet; birds that received 3.8 g TP kg-1 diet (no inorganic supplement) had the highest activity. The results obtained from this study indicate that it is possible to reduce P excretion with no adverse effect on the performance from three to seven weeks of age. In conclusion, dietary P could be reduced to 5.0-5.5 g kg-1 from 3 to 6 weeks and omitted from the withdrawal diets without adverse effects on live performance and this could decrease P intake and as a result P excretion.
PDF Abstract XML References Citation
Received: July 24, 2011;
Accepted: October 03, 2011;
Published: December 23, 2011
How to cite this article
Alaeldein M. Abudabos, 2012. Optimal Dietary Phosphorus for Broiler Performance Based on Body Weight Group. Asian Journal of Animal and Veterinary Advances, 7: 20-29.
DOI: 10.3923/ajava.2012.20.29
URL: https://scialert.net/abstract/?doi=ajava.2012.20.29
DOI: 10.3923/ajava.2012.20.29
URL: https://scialert.net/abstract/?doi=ajava.2012.20.29
INTRODUCTION
Cereal grains and oilseed meals are relatively high in P but a high percent of that P is present as Phytate Phosphorus (PP) (Wise, 1983; Bozkurt et al., 2006). Phytate, a salt of phytic acid, is the main storage form of phosphorus in plants has long been considered to be poorly utilized by chickens, due to the lack of endogenous phytase enzyme which hydrolyze phytate to myo-inositol and inorganic phosphate via intermediate myo-inositol phosphates (Nelson, 1976; Raboy, 1990; Ahmed et al., 2004). Several reports have shown that neither of these assumptions is correct; phytase is present in the mucosa of the small intestine of poultry (Bitar and Reinhold, 1972; Maenz and Classen, 1998; Abudabos et al., 2000) and PP utilization in chickens exceeds 50% (Edwards, 1983; Mohammed et al., 1991; Liem et al., 2009). There are several factors that affect the utilization and under P-deficient conditions, birds respond by increasing the utilization of PP to minimize decreases in performance (Punna and Ronald, 1999).
The lack of complete understanding of PP utilization and variation in bioavailability of P from inorganic sources may lead to over supplementation of P causing excessive amount of P to be excreted in poultry manure (Manangi and Coon, 2008; Bingol et al., 2009; Somkuwar et al., 2010). However, due to environmental concerns regarding the potential pollution from P in poultry excreta, there is great interest in providing adequate P to sustain productivity of poultry while reducing P excretion. Feeding broilers closer to requirement has been shown to reduce the concentration of P in the excreta (Sharpley, 1999; Waldroupet al., 2000; Leytem et al., 2008).
The greatest portion of skeletal development of the bird takes place during the first 3 weeks of life; it is generally recognized that the P requirement of the chick decreases with age (Skinner and Waldroup, 1992). Numerous studies have been conducted to determine P requirement of broiler chicks from d-old to 3 week of age. However, it is during the latter part of growing and finishing periods that P consumption and excretion are the greatest due to the greatest percentage of total feed consumption by the broilers (Dhandu and Angel, 2003). Several reports have shown that 5 g kg-1 TP is required for maximum growth. However, a higher level was required for maximum bone ash (Dhandu and Angel, 2003; Abas et al., 2011). The evidence suggests that P excretion in manure could be decreased by formulating grower and finisher diets with reduced concentration of inorganic P. The purpose of the present study was to examine the effect of dietary TP on performance, bone integrity and P excretion of broilers from three to seven weeks of age based on body weight group.
MATERIALS AND METHODS
Day-old, mixed chicks (Ross x Arbor Acres), obtained from a commercial hatchery, were used in this experiments. The chicks were wing-banded and randomly placed into 24 pens in an open-sided poultry house with partial environment control. The growth experiment was conducted for 28 days from March to April, 2009. Each earth floor pen was covered with about eight cm of wood shavings. The pen measured 2.6 m2 and was stocked with 25 chicks. Birds were individually weighed at 3 weeks and sorted into two groups: Heavy (H) and Light (L), with average body weights of 712 and 531 g, respectively and stocking density adjusted to 22 birds per pen from three to six weeks. The birds were individually weighed at six weeks, sorted into heavy and light body weight classes with mean body weight of 2089 and 1733 g, respectively and pen size adjusted to 13 birds. The birds were exposed to 23 h light from three to seven weeks of age.
Ingredients were analyzed for TP and Ca content (AOAC, 1984) and the adjusted values were used to formulate the diets. Chicks were fed a standard corn-soybean meal starter mash ad libitum to three weeks of age. In the experiment TP was varied in the grower and finisher diets. From three to six weeks of age, the diets contained 5.0, 5.5, 6.0 and 6.5 g TP kg-1 (Table 1). From six to seven weeks of age birds received diets that contained 3.8 (diet devoid of supplemental phosphate), 5.5, 6.0 and 6.5 g TP kg-1. Each diet was fed to six replicate pens. Considerable variation in body weight seen in previous experiments prompted a repeat of the experiment with body weight as a source of variance.
Individual body weight and feed consumption were recorded at six and seven weeks of age and feed conversion computed. At both ages one female and one male was sampled from each pen. Blood samples were collected by cardiac puncture for serum phosphorus (Baginski et al., 1967) and after euthanasia; the breast muscle was dissected and weighed (Uittenboogaart and Gerrits, 1982). Intestinal phytase activity was measured, a total of 36 birds per treatment were sampled for intestinal phytase activity (Iqbal et al., 1994; Biehl and Baker, 1997). The duodenal loop mucosa from three males and three females per treatment was pooled. Six pooled samples per sex within each treatment were used for the assay.
Statistical analysis: All statistical analyses were performed using the Statistical Analysis System (SAS) statistical package (SAS, 1996). Pen of birds constituted the experimental unit.
| Table 1: | Dietary ingredients (g kg-1) and chemical composition of the broiler chick grower and finisher diets |
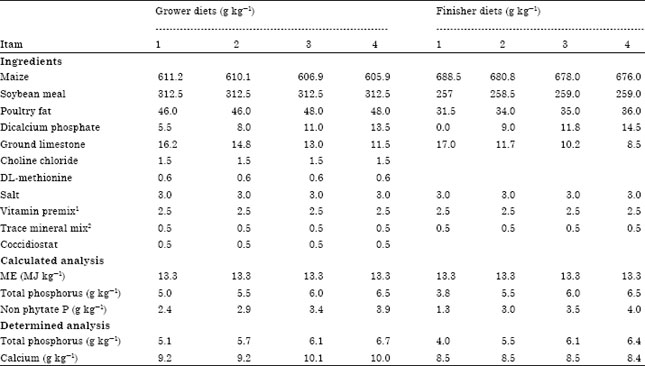 | |
| 1Vitamin mix provided per kg diet using Roche Broiler Premix: Retinyl acetate, 3.41 mg; cholecalciferol, 0.07 mg; DL-α-tocopherol acetate, 27.5 mg; menadione sodium bisulphate, 6 mg; riboflavin, 7.7 mg; niacin, 44 mg; pantothenic acid, cyanocobalamin, 0.02; choline 496 mg; folic acid, 1.32 mg; pyridoxine HCl, 4.82 mg; thiamine mononitrate, 2.16 mg; D-biotin, 0.11 mg. 2Mineral mix provided per kg diet using Mar-Jac Poultry Trace Mineral Mix: manganese, 67 mg; zinc, 54 mg; copper, 2 mg; iodine, 0.5 mg; iron, 75 mg; and selenium, 0.2 mg | |
The treatments were arranged in a split plot design. The whole plot factor was body weight class (H and L) and the sub-plot factor was level of TP. The experimental design for the whole plot was a randomized complete block design with 6 replications from 3 to 6 weeks and 3 replications from 6 to 7 weeks of age. The data were subjected to analysis of variance for a split-plot design using the Proc Mixed of SAS and the following statistical model:
Yijk = μ + αi + ρk + γik + βj + (αβ) j + εijk,
where, Yijk = variable measured, μ = overall mean, αi = effect for ith level of weight, ρk = the effect of the kth block, γik = random error for the weight, βj = effect for the jth level of phosphorus (αβ)ij = interaction effect for ith level of weight and jth level of phosphorus, εijk= random error term for subplot.
Significant differences among weight class or total phosphorus means were determined using pair wise comparisons. Means±SEM are presented in the tables and differences were considered statistically significant at p<0.05.
RESULTS
Decreasing dietary TP 6.5 to 5.0 g kg-1 from three to six weeks had no effect on feed intake, body weight gain and feed conversion (Table 2). Breast muscle yield was not influenced by any dietary treatment (Table 3).
| Table 2: | Effect of dietary phosphorus and weight group on feed consumption, body weight change and feed conversion, from three to six weeks in broiler chicks |
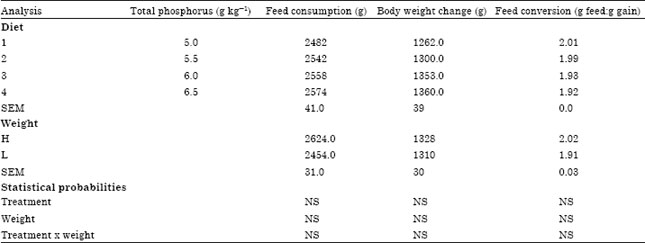 | |
| Within treatments means within columns followed by different superscripts are different (p<0.05). | |
| Table 3: | Effect of dietary phosphorus and weight group on breast muscle yield, serum P and phytase activity from three to six weeks in broiler chicks |
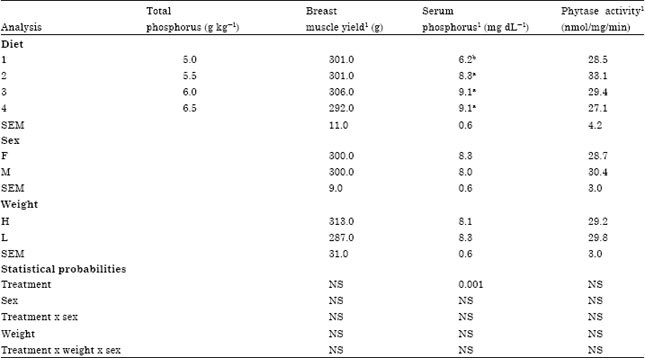 | |
| Within treatments means within columns followed by different superscripts are different (p<0.05). 1Each mean is based on a total of 12 birds per treatment at 42 day of age | |
A dietary TP x sex interaction was not detected for Serum P (Table 3), however, serum p values were lower (p<0.001) for birds fed 5.0 g kg-1 TP compared to the other 3 diets from three to six weeks. Serum P increased abruptly when TP was increased to 5.5 g kg-1 and as dietary TP increased from 5.5 to 6.5 g kg-1 serum P was not responsive. Intestinal phytase activity based on extracts from individual birds showed no significant differences between dietary treatments, weight groups and sex (p>0.05). Numerically, birds that received 5.5 g kg-1 TP had 22% higher phytase activity than those received 6.5 g kg-1 (Table 3).
| Table 4: | Effect of dietary phosphorus and weight group on feed consumption, body weight change and feed conversion from six to seven weeks in broiler chicks |
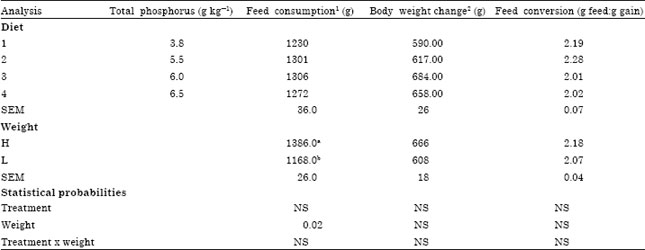 | |
| Within treatments means within columns followed by different superscripts are different (p<0.05). 1,2Feed consumption and body weight gain per bird during 8 d period. 1Each mean is based on 6 pens. 2Each mean is based on 13 bird’s weight in six pens | |
| Table 5: | Effect of dietary treatments on breast muscle yield, serum phosphorus and phytase activity from 6 to 7 weeks |
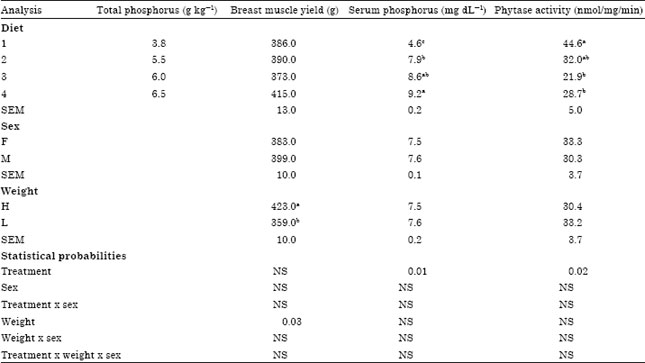 | |
| Within treatments means within columns followed by different superscripts are significantly different (p<0.05).1Each mean is based on 12 birds per treatment at 49 d of age | |
The responses to dietary TP at seven weeks (Table 4) were similar to those observed at six weeks for growth, feed conversion and breast muscle yield. When body weight group was used as a source of variance the only variable affected was feed consumption (Table 4). Birds in the heavy group consumed more feed than those in the light group. The gender effect on breast meat at seven weeks was not seen (Table 5). Serum P content was affected by dietary treatment (p<0.01) as shown in Table 5. Omitting the dicalcium phosphate from diet 1 resulted in 103% lower serum P compared to the control (6.5 g kg-1 TP). Phytase activity was affected by dietary treatment at 7 weeks (p<0.05). Birds that received a diet without inorganic P supplementation produced (39, 103 and 55%) higher phytase activity (p<0.02) than birds that received 5.5, 6.0 and 6.5 g TP kg-1, respectively (Table 5).
DISCUSSION
The results obtained from this study indicate that it is possible to reduce dietary P and as a result reduce P excretion with no adverse effect on the performance from three to seven weeks of age. During the growing period, there were no differences in body weight gain between birds fed the four levels of TP. Birds fed diets with 5.0 g kg-1 TP gained weight similarly to those fed the highest level 6.5 g kg-1. These results agreed with the results obtained when we measured the breast muscle yield, no significant effects were detected for the dietary treatments. Moreover, TP did not affect feed consumption or FCR. In agreement with the results obtained by Chen and Moran (1995) feed to gain ratio was not affected by the level of P in the diet, however, they reported 8 and 12% reduction in body weight and gain in Ross x Ross males from 3-6 weeks when they fed a low P growing feed (3.5 g kg-1) compared to control (6.3 g TP kg-1). In this experiment, we found 11.5% reduction in body weight gain from three to seven weeks period when the birds were fed 30% less TP but this was not a significant difference. Several studies reported that after 3 weeks of age P requirements are greatly reduced and there is little or no need for P supplementation (Skinner et al., 1992). Waldroup et al. (2000) found that available P could be reduced by 0.75 g kg-1 within an age period from zero to three, three to six and six to eight weeks of age without adverse effects on broiler growth. Several reports have shown that 5.0 g kg-1 P level was required for maximum growth (Skinner et al., 1992; Chen and Moran, 1995; Dhandu and Angel, 2003). Yan et al. (2001) did not find any significant difference in body weight gain of broilers which had fed levels of available P from 1.5 to 4.5 g kg-1 from three to seven weeks. However, recently, Shaw et al. (2010) found that body weight increased significantly with an increase in available P from 3.5 to 5.0 g kg-1 at four week. On the other hand, a higher level of P was required for maximum bone ash (Fritz et al., 1969; Dhandu and Angel, 2003). Yan et al. (2001) reported that tibia ash was significantly lower for of broilers fed 1.5 compared to those fed 4.5 g kg-1 available P; 3.3, 1.86 and 1.63 g kg-1 of available P were required to optimize tibia ash, body weight gain and FCR, respectively.
Live performance was not affected when the inorganic P source was omitted from the finishing diets. No significant differences could be detected for any dietary treatment on feed intake, body weight gain and FCR. The body weight gain data agreed with the breast muscle weight data since no differences in performance could be detected between treatments at six to seven weeks of age. Previous investigators had removed inorganic P source from the withdrawal feed of broiler males without adverse effects on body weight gain, feed intake, feed utilization and carcass yield (Skinner et al., 1992; Chen and Moran, 1994, 1995). Waldroup et al. (1974) studied the requirement of broilers for P between four and eight weeks of age. They reported no more than 1.2 g kg-1 added inorganic P was needed in addition to the 4.2 g kg-1 P present in the corn-soy meal diet. Yan et al. (2000) reported that males fed diets with 0.75 g kg-1 less available P did not differ significantly in body weight (at six and eight weeks old) from those fed NRC level but they detected significant differences when the birds fed diets with 1.5 g kg-1 less than the NRC level.
Serum P showed treatment and sex effects during growing and finishing periods. Gardiner (1962) concluded that the level of plasma inorganic P responded very quickly to suboptimal dietary P levels and to feed withdrawal. During the growing period, birds that received 5.5 g kg-1 TP showed no significant differences than those that received the highest level of TP. During the finishing period, serum P was inconsistently responsive to dietary P levels. Birds that received any level of inorganic P supplementation showed no significant differences among them. Females in both periods had a higher serum P compared to males; this could be explained by the fact that males have more phosphate deposited in their bones since they have longer and heavier bones.
Dietary P levels have been shown to influence the utilization of PP (Temperton and Cassidy, 1964; Ballam et al., 1984; Simons et al., 1990; Mitchell and Edwards, 1996). These reports indicated that PP retention increased when low levels of dietary P were fed. According to Punna and Ronald (1999), dietary P level significantly affect PP utilization. Broiler that consumed the P-deficient diet utilized significantly more PP than birds fed the control diet at the end of second and fourth weeks (69.4 vs. 17.1% and 65.5 vs. 19.5%), respectively. In the current experiment PP utilization was not measured but intestinal phytase activity was measured.
Phytase enzyme improves performance by increasing the availability of PP; the ability of broilers to utilize PP becomes critically important, especially under P-deficient conditions. This increase in PP utilization is directly related to increased activity of intestinal phytase (Davies et al., 1970; McCuaig et al., 1972). Literature indicates that intestinal phytase activity is subject to regulatory mechanisms and it contributes to the utilization of PP. The activity of the enzyme may be regulated by the mineral status of the animal (Matterson, 1948; McCuaig et al., 1970; Davies et al., 1970). In this experiment, we found that birds that received the low P diets had double the phytase activity. The results of the present study agree with the reports of Davies et al. (1970) and McCuaig et al. (1972) in which they found that intestinal phytase activity was more than twice as high when the diet contained no supplemental phosphorus compared to a supplemented diet. At 42 day of age phytase activity showed no differences between treatments, weight and sex. Numerically, 22% higher phytase activity was measured in mucosal extracts from birds that received 5.5 g TP kg-1 compared to those fed 6.5 g kg-1.
Body weight group had little influence on criteria measured in this experiment from six to seven weeks of age. Birds from the heavy group consumed more feed and produced heavier breast muscles compared to the light group. Generally, it was found that variation in body weight between birds on the same treatment group had a little influence on P utilization. Punna and Ronald (1999) suggested that a great variation among birds in utilizing PP influences the phosphorus requirements of chickens. The larger birds retained more PP and used more TP than the smaller birds within the P-deficient group.
The results of this study, in which variation in body weight was included as a source of variation, indicated that the TP levels needed by broilers to maintain growth and feed utilization from three to seven weeks could be lower than that recommended by NRC (1994). On the other hand, retention of P will decrease as the P consumed surpasses the requirement. Birds utilized PP, especially those that received a low level of inorganic source during the growing period and no supplementation during finishing period.
CONCLUSION
The results of this study clearly show that dietary P could be reduced to 5.0-5.5 g kg-1 from three to six weeks and omitted from the withdrawal diets without adverse effects on live performance and this could decrease P intake and as a result P excretion.
ACKNOWLEDGMENTS
This project was supported by King Saud University, Deanship of Scientific Research, College of Food and Agriculture Sciences, Research Center.
REFERENCES
- Ahmed, F., M.S. Rahman, S.U. Ahmed and M.Y. Miah, 2004. Performance of broiler on phytase supplemented soybean meal based diet. Int. J. Poult. Sci., 3: 266-271.
CrossRefDirect Link - Baginski, E.S., P.P. Foa and B. Zak, 1967. Determination of phosphate: Study of labile organic phosphate interference. Clin. Chem. Acta, 15: 155-158.
CrossRef - Ballam, G.C., T.S. Nelson and L.K. Kirby, 1984. Effect of fiber and phytate source and of calcium and phosphorus level on phytate hydrolysis in the chick. Poult. Sci., 63: 333-338.
PubMed - Biehl, R.R. and D.H. Baker, 1997. 1α-hydroxycholecalciferol does not increase the specific activity of intestinal phytase but does improve phosphorus utilization in both cecectomized and sham-operated chicks fed cholecalciferol-adequate diets. J. Nutr., 127: 2054-2059.
Direct Link - Bitar, K. and J.G. Reinhold, 1972. Phytase and alkaline phosphatase activities in intestinal mucosae of rat, chicken, calf and man. Biochim. Biophys. Acta, 268: 442-452.
PubMed - Chen, X. and E.T. Moran Jr., 1994. Response of broilers to omitting dicalcium phosphate from the withdrawal feed: Live performance, carcass downgrading and further-processing yields. J. Appl. Poult. Res., 3: 74-79.
Direct Link - Chen, X. and E.T. Moran Jr., 1995. The withdrawal feed of broilers: Carcass responses to dietary phosphorus. J. Applied Poult. Res., 4: 69-82.
Direct Link - Davies, M.I., G.M. Ritcey and I. Motzok, 1970. Intestinal phytase and alkaline phosphatase of chicks influence of dietary calcium inorganic and phytate phosphorus and vitamin D3. Poult. Sci., 49: 1280-1286.
PubMed - Dhandu, A.S. and R. Angel, 2003. Broiler nonphytin phosphorus requirement in the finisher and withdrawal phases of a commercial four-phase feeding system. Poult. Sci., 82: 1257-1265.
Direct Link - Edwards, H.M. Jr., 1983. Phosphorus 1. Effect of breed and strain on utilization of sub-optimal levels of phosphorus in the rations. Poult. Sci., 62: 77-84.
PubMed - Fritz, J.C., T. Roberts, J.W. Boehne and E.L. Hove, 1969. Factors affecting the chick's requirement for phosphorus. Poult. Sci., 48: 307-320.
PubMed - Iqbal, T.H., K.O. Lewis and B.T. Cooper, 1994. Phytase activity in the human and rat small intestine. Gut, 35: 1233-1236.
Direct Link - Leytem, A.B., G.P. Widyaratne and P.A. Thacker, 2008. Phosphorus utilization and characterization of ileal digesta and excreta from broiler chickens fed diets varying in cereal grain, phosphorus level and phytase addition. Poult. Sci., 87: 2466-2476.
CrossRef - Liem, A., G.M. Pesti, A. Atencio and H.M. Edwards, Jr., 2009. Experimental approach to optimize phytate phosphorus utilization by broiler chickens by addition of supplements. Poult. Sci., 88: 1655-1665.
PubMed - Maenz, D.D. and H.L. Classen, 1998. Phytase activity in the small intestinal brush border membrane of the chicken. Poult. Sci., 77: 557-563.
CrossRefDirect Link - Manangi, M.K. and C.N. Coon, 2008. Phytate phosphorus hydrolysis in broilers in response to dietary phytase, calcium and phosphorus concentrations. Poult. Sci., 87: 1577-1586.
CrossRef - McCuaig, L.W., M.I. Davies and I. Motzok, 1972. Intestinal alkaline phosphatase and phytase of chicks: Effect of dietary magnesium, calcium, phosphorus and thyroactive casein. Poult. Sci., 51: 526-530.
PubMed - Mitchell, R.D. and H.M. Jr. Edwards, 1996. Effects of phytase and 1,25 dihydroxycholecalciferol on phytate utilization and the quantitative requirement for calcium and phosphorus in young broiler chickens. Poult. Sci., 75: 95-110.
PubMed - Mohammed, A., M.J. Gibney and T.G. Taylor, 1991. The effects of dietary levels of inorganic phosphorus, calcium and cholecalciferol on the digestibility of phytate-P by the chick. Br. J. Nutr., 66: 251-259.
PubMed - Nelson, T.S., 1976. The hydrolysis of phytate phosphorus by chicks and laying hens. Poult. Sci., 55: 2262-2264.
PubMed - NRC., 1994. Nutrient Requirements of Poultry. 9th Edn., National Academy Press, Washington, DC., USA., ISBN-13: 9780309048927, Pages: 176.
Direct Link - Punna, S. and D.A. Ronald, 1999. Variations in the phytate phosphorus utilization within the same broiler strain. J. Applied Poult. Res., 8: 10-15.
CrossRefDirect Link - Sharpley, A., 1999. Agricultural phosphorus, water quality and poultry production: Are they compatible? Poult. Sci., 78: 660-673.
CrossRefDirect Link - Shaw, A.L., J.P. Blake and E.T. Moran, 2010. Effects of flesh attachment on bone breaking and of phosphorus concentration on performance of broilers hatched from young and old flocks. Poult. Sci., 89: 295-302.
CrossRefDirect Link - Simons, P.C.M., H.A.J. Versteegh, A.W. Jongbloed, P.A. Kemme and P. Slump et al., 1990. Improvement of phosphorus availability by microbial phytase in broilers and pigs. Br. J. Nutr., 64: 525-540.
CrossRefDirect Link - Skinner, J.T., A.L. Izat and P.W. Waldroup, 1992. Effects of removal of supplemental calcium and phosphorus from broiler finisher diets. J. Applied Poult. Res., 1: 42-47.
Direct Link - Skinner, J.T. and P.W. Waldroup, 1992. Effects of calcium and phosphorus levels in starter and grower diets on broilers during the finisher period. J. Applied Poult. Res., 1: 273-279.
Direct Link - Somkuwar, A.P., K. Ravikanth, S. Maini, D.S. Rekhe, M.K. Patil and B. Ballurkar, 2010. Phytase with synergistic herbs: An option to reduce environmental pollution by partial replacement of inorganic phosphorus in broiler ration. Int. J. Poult. Sci., 9: 390-394.
CrossRefDirect Link - Bingol, N.T., M.A. Karsli, D. Bolat, I. Akca and T. Levendoglu, 2009. Effects of microbial phytase on animal performance, amount of phosphorus excreted and blood parameters in broiler fed low non-phytate phosphorus diets. Asian J. Anim. Vet. Adv., 4: 160-166.
CrossRefDirect Link - Waldroup, P.W., J.H. Kersey, E.A. Saleh, C.A. Fritts and F. Yan et al., 2000. Nonphytate phosphorus requirement and phosphorus excretion of broiler chicks fed diets composed of normal or high available phosphate corn with and without microbial phytase. Poult. Sci., 79: 1451-1459.
CrossRefPubMedDirect Link - Waldroup, P.W., R.J. Mitchell and K.R. Hazen, 1974. The phosphorus needs of finishing broilers in relationship to dietary nutrient density levels. Poult. Sci., 53: 1655-1663.
PubMed - Yan, F., J.H. Kersey, C.A. Fritts, P.W. Waldroup and H.L. Stilborn et al., 2000. Evaluation of normal yellow dent corn and high available phosphorus corn in combination with reduced dietary phosphorus and phytase supplementation for broilers grown to market weights in litter pens. Poult. Sci., 79: 1282-1289.
Direct Link - Yan, F., J.H. Kersey and P.W. Waldroup, 2001. Phosphorus requirements of broiler chicks three to six weeks of age as influenced by phytase supplementation. Poult. Sci., 80: 455-459.
PubMedDirect Link