
Meryem Eren
Department of Biochemistry, Faculty of Veterinary Medicine, University of Erciyes, 38039 Melikgazi, Kayseri, Turkey
Fatma Uyanik
Vocational Health College, University of Onsekiz Mart Canakkale, Canakkale 17100, Turkey
Berrin Kocaoglu Guclu
Department of Animal Nutrition and Nutritional Diseases, Faculty of Veterinary Medicine, University of Erciyes, 38039 Melikgazi, Kayseri, Turkey
Ayhan Atasever
Department of Pathology, Faculty of Veterinary Medicine, University of Erciyes, 38039 Melikgazi, Kayseri, Turkey
Asian Journal of Animal and Veterinary Advances
Year: 2012 | Volume: 7 | Issue: 11 | Page No.: 1079-1089







ABSTRACT
This study was performed to investigate the effect of adding boron (B) to broilers’ diets on performance, some blood biochemical parameters and visual and histopathological examinations of organs in broilers. One hundred and forty four chicks were randomly assigned to 4 groups with 3 replicates. The broilers were fed commercial diets supplemented with 0 (control group) and 500, 750, 1000 mg kg-1 (diet) B from Boric Acid (BA) for 42 days. Live body weight, weight gain, food consumption and feed conversion ratio were recorded at weekly intervals. At the end of the experimental period (42 days of age), serum Aspartate aminotransferase (AST), Alanine aminotransferase (ALT), Gamma Glutamyl Transpeptidase (GGT), Alkaline Phosphatase (ALP), Creatine Kinase (CK), Lactate Dehydrogenase (LDH) enzyme activities and glucose, creatinine, total protein, albumin, globulin, triglyceride, total cholesterol, High Density Lipoprotein-cholesterol (HDL-C), Low Density Lipoprotein-cholesterol (LDL-C), calcium (Ca), phosphorus (P), magnesium (Mg) and B levels were determined. Performance parameters were affected negatively and significantly by B supplementation. Serum AST, ALT, CK, LDH activities and glucose, total protein, albumin, globulin, triglycerides, total cholesterol, LDL-C and Mg levels were significantly influenced by dietary B supplementation. Necrosis in muscle fibers was observed but no gross and microscopic changes were determined in liver, kidney and brain in histological examinations in B supplemented groups. In conclusion; add boron to broilers’ diets at levels 500, 750 and 1000 mg kg-1 (feed) did not cause mortality or result in histopathological lesions except for muscles. On the other hand, it resulted in a significant decrease in performance and changed some biochemical parameters in broilers.
PDF Abstract XML References Citation
Received: May 23, 2012;
Accepted: July 09, 2012;
Published: September 15, 2012
How to cite this article
Meryem Eren, Fatma Uyanik, Berrin Kocaoglu Guclu and Ayhan Atasever, 2012. The Influence of Dietary Boron Supplementation on Performance, Some Biochemical Parameters and Organs in Broilers. Asian Journal of Animal and Veterinary Advances, 7: 1079-1089.
DOI: 10.3923/ajava.2012.1079.1089
URL: https://scialert.net/abstract/?doi=ajava.2012.1079.1089
DOI: 10.3923/ajava.2012.1079.1089
URL: https://scialert.net/abstract/?doi=ajava.2012.1079.1089
INTRODUCTION
Boron (B) is an essential trace element for human, animals and higher plants and it is found in every part of the environment (McDowell, 1992; WHO, 1998; Laila and Adel, 2002; Tariq and Mott, 2007; Shaaban, 2010). However the exact metabolic and biochemical functions of B are not fully understood, it has been suggested that B have important effects on various metabolic and physiological systems (bone, mineral, lipid, energy metabolism, endocrine function etc.) of the in animal and human organism (Nielsen et al., 1988; Hall et al., 1989; Hunt and Herbel, 1991a, b; Kurtoglu et al., 2001, 2005; Gallardo-Williams et al., 2003; Karabulut and Eren, 2006; Eren and Uyanik, 2007; Basoglu et al., 2010). Boron, in the form of various inorganic borates, is widely distributed in low concentrations throughout nature and, used in industrial and domestic applications. It is used in the production of fibreglass, borosilicate glass, enamels, frits, glazes and as cosmetics, pharmaceuticals (as a pH buffer), boron capture therapy, insecticide, antiseptic and chemical preservative in food products (Woods, 1994; WHO, 1996, 1998; See et al., 2010).
The oral toxicity of B is low (McDowell, 1992; WHO, 1998). On the other hand, B accumulation was shown in bone, muscle, adrenal tissue, brain, hypothalamus, liver, spleen, kidney, adrenals, lymph node, testis, seminal vesicles, prostate, large intestine and blood after exposure to high doses of B (Ku et al., 1991; Sander et al., 1991; Moseman, 1994; Tibbits et al., 2000) in various animals. It is reported that disorders such as edema and inflammation in the skin around the nail and legs, depressed live weight gains and feed consumption, low haematocrit, haemoglobin, triglyceride, protein, phosphorus and ALP levels, riboflavinuria, paralysis in toe, disorder of calcium metabolism, osteoporosis with the decreasing of the parathyroid activity, inhibition of sperm production, testicular atrophy, disorder of the ovarium development, alopecia, dermatitis, hypertonia and even death were seen depending on the excessive intake of the B element in several animals (Nielsen et al., 1988; Sander et al., 1991; Treinen and Chapin, 1991; McDowell, 1992; Naghii and Samman, 1993; Ku and Chapin, 1994; WHO, 1996, 1998; Bustos-Obregon and Ricardo Hartley, 2008; See et al., 2010). Geyikogglu and Turkez (2007) suggested that 3.375 and 4.5 mmol B kg-1 b.wt. (750 and 1000 ppm boric acid) caused decreased metabolite concentrations (glucose, glycogen, lactate and ATP) in muscle fibers as dependent on time in broiler chickens.
Boric acid is used in the poultry environment primarily to litter which could lead to accidental consumption of the chemical by poultry. Sander et al. (1991) reported that boric acid is highly efficient against cockroaches and darkling beetles determined in poultry houses. Several authors have investigated the effects of different doses of B up to 320 mg kg-1 supplementation to diet in broilers (Rossi et al., 1990, 1993; Qin and Klandorf, 1991; Elliot and Edwards, 1992; Kurtoglu et al., 2001, 2005; Fassani et al., 2004; Geyikogglu and Turkez, 2007; Yildiz et al., 2011; Bozkurt et al., 2012) but there are no studies about the effects of higher doses than 320 mg kg-1 on performance and blood biochemical parameters in broilers. The determination of maximum tolerable doses of boric acid in broilers is also important. Therefore, this study was performed to investigate the effects of adding 500, 750 and 1000 mg kg-1 boron supplementation to diet on performance, some biochemical parameters and organs in broilers.
MATERIAL AND METHODS
Animals and experimental design: One hundred and forty four, day old, broiler chicks were used in this study. Chicks were randomly assigned to 4 groups with 3 replicates (12 chicks each). Chicks of each replicate were kept in separate floor pens with 24 h constant light schedule. The broilers were fed commercial diets (starter and grower) supplemented with 0 (control group) and 500, 750 and 1000 mg kg-1 (diet) B from boric acid (H3BO3, Carlo Erba) from 0-42 days as experimental period. Ingredients and chemical composition of the basal diet fed to broilers was shown in Table 1. Feed and water was supplied ad libitum. This study was approved by the Ethics Committee of University of Erciyes, Faculty of Veterinary Medicine, Approval number, 2004-45-59.
Measurements of performance: Live Body Weight (LBW), Body Weight Gain (BWG) and Feed Consumption (FC) were weekly recorded. Feed Conversion Ratio (FCR) was calculated by dividing feed consumption by live weight gain.
| Table 1: | Ingredients and chemical composition of basal diets fed to broilers |
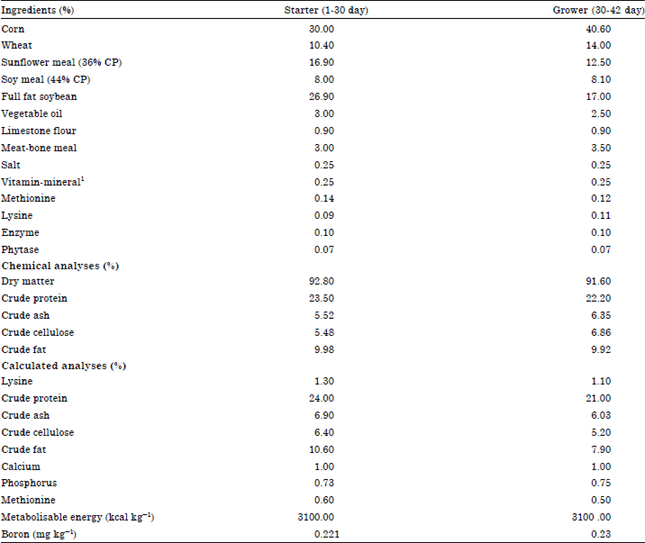 | |
| CP: Crude protein, 1Provided by per kg of diet: Vitamin A: 12000 IU, Vitamin D3: 1500 IU, Vitamin E: 30 mg, Vitamin K3: 5 mg, Vitamin B1: 3 mg, Vitamin B2: 6 mg, Vitamin B6: 5 mg, Vitamin B12: 0.03 μg, Nicotinamide: 40 mg, Vitamin C: 50 mg, Calcium D-pantothenate: 10 mg, Folic acid: 0.75 μg, Biotin: 0.075 μg, Choline chloride: 375 mg, Mn: 80 mg, Fe: 40 mg, Zn: 60 mg, Cu: 5 mg, I: 0.4 μg, Co: 0.1 μg, Se: 0.15 μg, Antioxidant: 10 mg | |
Sample collection and analysis: Blood samples were taken from V. brachialis of the animals (12 broilers from each group, 4 animals per replicates) at the end of the experimental period. Sera were separated by centrifugation at 3000 rpm for 10 min after 1 h incubation at room temperature and stored at -20°C until the analysis. Sera were analysed by a spectrophotometer (Shimadzu UV Model 1208) using commercial kits for AST, ALT, GGT, ALP, CK, LDH activities, glucose, creatinine, total protein, albumin, Ca, P, Mg (Biolabo, France), triglycerides, total cholesterol, HDL-C and LDL-C (AMP, Austria) levels. The serum globulin concentrations were calculated by subtracting the albumin values from the total protein values. Chemical composition of the diet was analysed by the method of AOAC (1984). Boron levels in sera and in basal diet were determined by ICP (Inductively coupled plasma atomic emission spectrometer; AES Varion Vista Model, Sydney, Australia). Twelve chicks from each group were killed by neck dislocation and gross visual examination of organs was made during the necropsy procedure to determine tissue damage and liver, kidney, brain and muscle samples were preserved in buffered formalin for histopathological evaluations. Tissue samples were embedded in paraffin, sectioned (5-6 μm), stained with hematoxylin and eosin (H and E) and then examined with light microscope (Luna, 1968).
Statistical analysis: Statistical analyses of data were performed by SPSS 15.0 version for Windows. One-way analysis of variance (ANOVA) was used for the differences between groups. When the F values were significant, Duncan’s Multiple Range Test was performed. All data were expressed as Mean±SEM. Results are considered as significant when the p-value was less than 0.05.
RESULTS AND DISCUSSION
In the present study, no mortality was seen in B supplemented groups as indicated by Hoffman et al. (1991) who supplemented diet of ducklings with 1000 mg kg-1 B. This can be attributed to the doses used in this study, as 500, 750 and 1000 mg kg-1 B were tolerated by the broilers because of B is absorbed rapidly and virtually completely from gastrointestinal tract and rapidly excreted (McDowell, 1992; WHO, 1998).
As shown in Table 2 and 3, LBW and BWG (p<0.001) were decreased significantly in 500, 750 and 1000 mg kg-1 B groups compared with control group. In regard to B treated groups, there was a significant decrease in LBW which was B-doses dependent. Feed consumption (p<0.05) was depressed insignificantly by 500 and 750 mg kg-1 B and significantly by 1000 mg kg-1 B. Similar results were obtained in broilers fed 300 (Rossi et al., 1993) and 320 mg kg-1 B (Rossi et al., 1990), in ducklings fed 1000 mg kg-1 B (Hoffman et al., 1991) and layer (Wilson and Ruszler, 1996, 1998; Eren et al., 2004) fed 400 mg kg-1 B. Feed efficiency (p<0.01) was negatively affected by 1000 mg kg-1 B as indicated by Rossi et al. (1990) who supplemented the diet of broiler chicks with 320 mg kg-1 B. Decreased LBW and BWG that observed in B treated groups may be resulted from the reduction feed intake.
Our results showed that decreases were determined in serum total protein in 750 and 1000 mg kg-1 B groups, albumin in all of the B groups (p<0.01) and globulin (p<0.05) levels in 1000 mg kg-1 B group compared with control group in the present study (Table 4). This result is in agreement with the findings of Hoffman et al. (1991) who reported that plasma protein concentration decreased in the presence of 1000 mg B kg-1 feed in ducklings. The decreases in protein levels may attribute to the decrease in feed consumption and thus the decrease of the amount of protein which absorbed through small intestine.
| Table 2: | Effect of adding different levels of boron to broilers’ diet on live body weight (LBW) |
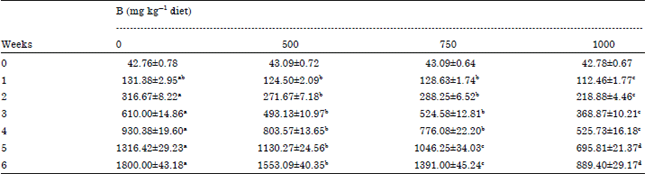 | |
| Values (Mean±SEM) within each row with different superscripts differ significantly at p<0.001 | |
| Table 3: | Effect of adding different levels of boron to broilers’ diet on feed consumption (FC) (g), body weight gain (BWG) (g) and feed conversion ratio (FSR) (g feed g-1 LBW) |
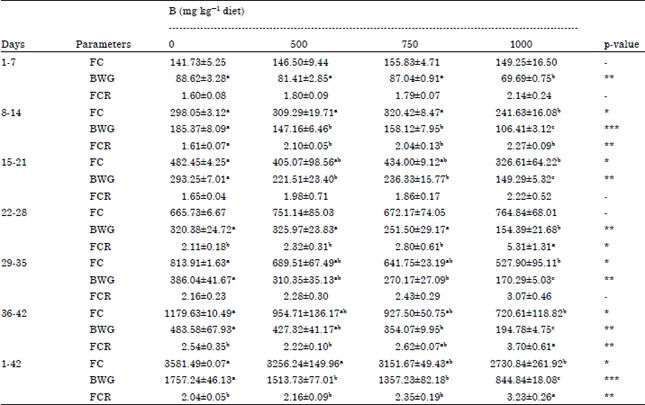 | |
| Values (Mean±SEM) within each row with different superscripts differ significantly at *p<0.05, **p<0.01 and ***p<0.001 | |
Serum B level was increased (p<0.001) in B treatment groups compared to the control group and this effect was B-doses dependent in this study (Table 4). In spite of, feed consumption of 1000 mg B treated group was reduced by (-16.15%) compared to the 500 mg B group and by (-13.36%) compared to the 750 mg B group, the serum B concentration was higher in the 1000 mg treated group compared to the other two B levels. This effect attributed to the increase of B absorption from intestine to the blood because increase B level in this group by 100 and 50% compared to the other two B levels, respectively.
As shown in Table 4, serum AST activity was highly significant decreased (p<0.01) in all groups fed diets supplemented with different levels of B and this effect was B-doses dependent and ALT enzyme activity was decreased (p<0.001) in a like manner with the AST but the significant was with the 750 and 1000 mg B only. Whereas significant increases in CK enzyme activity (p<0.01) with 750 and 1000 mg B doses and LDH enzyme activity (p<0.05) with 500, 750 and 1000 mg B doses were observed in B treated groups than the control group. It was reported that serum AST activity was decreased by 100, 200 and 400 mg kg-1 B supplementation in laying hens (Eren and Uyanik, 2007). The AST activity in rats (Hunt and Herbel, 1991a) and both AST and ALT activities in mice (Ince et al., 2011) were also decreased by B in previous studies. The decreases in AST and ALT enzyme activities by B supplementation in this study may result from the protective effects of the B on normal liver metabolism (Hunt and Herbel, 1991a; Ince et al., 2011). Skeletal muscle contains high activities of CK and LDH. Cell injury with subsequent leakage of the enzymes is the most common cause of their increased activities.
| Table 4: | Effect of adding different levels of boron to broilers’ diet on some serum enzyme activities and metabolite levels in broilers (Mean±SEM) |
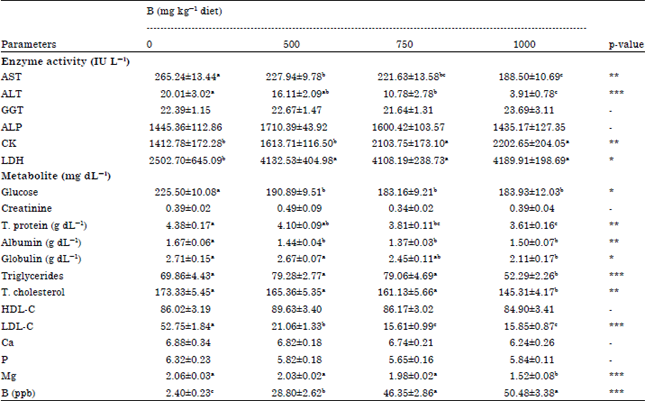 | |
| Values (Mean±SEM) within each row with different superscripts differ significantly at *p<0.05, **p<0.01 and ***p<0.001, n = 12 | |
Serum CK and LDH enzymes are useful diagnostically for identifying skeletal muscle disease (Meyer and Harvey, 1998), therefore, the increases in serum CK and LDH activities may be due to muscle damage characterised by necrosis in muscle fibers in B supplemented groups (Fig. 1-3) in the present study. On the other hand, no significant differences in serum GGT and ALP activities, creatinine, HDL-C, Ca and P levels were found between control and B treatment groups (Table 4) and no gross and microscopic changes were observed in histological examinations in liver, kidney and brain.
Boric acid supplementation has induced changes in serum glucose and lipid profiles (Table 4). In compared with control, serum glucose level was gradually and significantly decreased (p<0.05) in B groups. There was no change in glucose level among B treated groups. The decrease of serum glucose levels with supplementation of B consistent with the result of the studies that tested B in broiler (Hunt, 1989), in laying hens (Eren and Uyanik, 2007), in rats (Hunt and Herbel, 1991a), in dogs (Basoglu et al., 2000) and the decreases in serum glucose levels may be result of the binding of B to hydroxyl groups of α-glucose (Hunt, 1989; WHO, 1998) in the present study. Decreased total cholesterol significantly (p<0.01) in all B treated groups and triglycerides (p<0.001) in 1000 mg kg-1 B supplemented group and LDL-C (p<0.001) in all of the B treated groups levels may indicate that B is a hypolipidemic agent in this study. Our findings are in agreement with that of the previous studies in rat (Hall et al., 1989), rabbit (Basoglu et al., 2010) and laying hens (Eren and Uyanik, 2007).
Boron has an essential function that regulates parahormone activity (McDowell, 1992) and therefore, effects Ca, P, Mg and cholecalciferol metabolisms (McDowell, 1992, Miljkovic et al., 2004; Kurtoglu et al., 2001, 2005) and it has been shown to prevent bone demineralization and affect cartilage of bone and normal development and plasma mineral levels (Ca, P, Mg) (Hunt, 1989; Nielsen et al., 1988; Wilson and Ruszler, 1998; Kurtoglu et al., 2001).
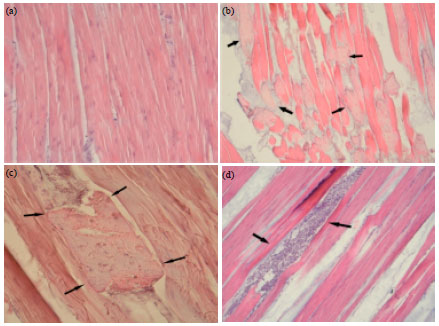 | |
| Fig. 1(a-d): | Broiler muscles with (a) Normal appearance in control group, (b) Homogeneous pink-coloured necrotic areas (arrows), (c) Breakdown of muscle fiber depending on necrosis (arrows) and (d) Foci of mononuclear cell infiltration in necrotic areas between in muscle fibers (arrows), H and E, x400 |
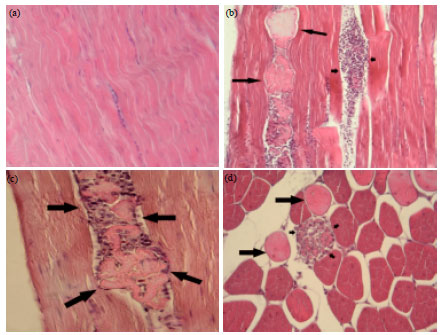 | |
| Fig. 2(a-d): | Broiler muscle with (a) Normal appearance, control group, (b) Necrosis of muscle fibers (large arrows) and mononuclear cell infiltrations (small arrows), (c) Mononuclear cells in necrotic area of muscle fibers (arrows) and (d) Necrotic areas (large arrows) and mononuclear cell infiltrations (small arrows) in transverse cross section of muscle fibers, H and E, x200 |
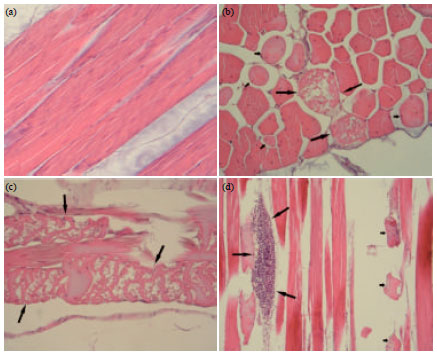 | |
| Fig. 3(a-d): | Broiler muscle with (a) Normal appearance, control group, (b) Homogeneous pink-coloured necrotic areas (small arrows) and fragmented muscles (large arrows) in transverse cross section of muscle fibers, (c) Fragmented muscles as a result of necrosis in longitudinal cross section of muscle fibers (arrows) and (d) Necrotic muscles (small arrows) and mononuclear cell infiltrations (large arrows) in necrotic areas between in muscle fibers, H and E, x200 |
In this study, serum Mg levels decreased significantly (p<0.001) in 1000 mg kg-1 B treated chickens (Table 4). The decreases in the levels of Mg may result from either the reductions of utilisation in the gastrointestinal tract or the improvement of urinary losses (Hunt and Herbel, 1991b; Chapin et al., 1997; Gallardo-Williams et al., 2003) due to B supplementation as reported by the previous studies in broilers (Qin and Klandorf, 1991; Kurtoglu et al., 2001), in quails (Karabulut and Eren, 2006) and in rats (Nielsen et al., 1988; Hunt and Herbel, 1991b; Chapin et al., 1997).
CONCLUSION
The result of this study have shown that supplementation of 500, 750 and 1000 mg kg-1 boron in the form of boric acid did not cause mortality or result in histopathological lesions except for muscles but did cause a significant decrease in performance and changed some blood biochemical parameters in broilers. Especially, 1000 mg kg-1 B had harmful effect than 500 and 750 mg kg-1 B on growth performance and some blood chemistry. Generally, serum glucose, lipid profiles, AST and ALT enzyme activities as indicator of liver function were positively affected by dietary B supplementation. However, adverse effects in growth performance parameters, serum proteins, CK and LDH enzyme activities as sign of muscle damage and pathological lesion in the muscle were observed. In conclusion, further more detailed investigations with B at various levels or different B compounds are needed to better understand the biochemical functions of boron in different animal species.
ACKNOWLEDGMENT
This research was supported by Erciyes University Research Fund. Project No. VA-05-01.
REFERENCES
- Basoglu, A., M. Sevinc, H. Guzelbektas and T. Civelek, 2000. Effect of borax on serum lipid profile in dogs. Online J. Vet. Res., 4: 153-156.
Direct Link - Basoglu, A., N. Baspinar, S.A. Ozturk and P.P. Akalin, 2010. Effects of boron administration on hepatic steatosis, hematological and biochemical profiles in obese rabbits. Trace Elements Electrolytes, 27: 225-231.
CrossRefDirect Link - Bozkurt, M., K. Kucukyılmaz, A.U. Catlı, M. Cınar, M. Cabuk and E. Bintas, 2012. Effects of boron supplementation to diets deficient in calcium and phosphorus on performance with some serum, bone and fecal characteristics of broiler chickens. Asian-Aust. J. Anim. Sci., 25: 248-255.
Direct Link - Elliot, M.A. and H.M. Edwards, Jr., 1992. Studies to determine whether an interaction exists among boron, calcium and cholecalciferol on skeletal development of broiler chickens. Poult. Sci., 71: 677-690.
PubMedDirect Link - Fassani, E.J., A.G. Bertechini, J.A.K. Brito, R.K. Kato, E.T. Fialho and A. Geraldo, 2004. Boron supplementation in broiler diets. Revista Brasileira Ciencia Avicola, 6: 231-237.
CrossRef - Hall, I.H. B.F. Spielvogel, T.S. Griffin, E.L. Docks and R.J. Brotherton, 1989. The effects of boron hypolipidemic agents on LDL and HDL receptor binding and related enzyme activities of rat hepatocytes, aorta cells and human fibroblasts. Res Comm.Chem. Pathol. Pharmacol., 65: 297-317.
PubMed - Hoffman, D.J., C.J. Sanderson, L.J., Le Captain, E. Cromatie and G.W. Pendleton, 1991. Interactive effects of boron, selenium and dietary protein on survival, growth, and physiology in mallard ducklings. Arch. Environ. Con. Tox., 20: 288-294.
CrossRef - Hunt, C.D. and J.L. Herbel, 1991. Boron affects energy metabolism in the streptozotocin-injected, vitamin D3-deprived rat. Magnes. Trace Elem., 10: 374-386.
PubMed - Hunt, C.D. and J.L. Herbel, 1991. Effects of dietary boron on calcium and mineral metabolism in the streptozotocin-injected, vitamin D-3-deprived rat. Magnes. Trace Elem., 10: 387-408.
PubMed - Hunt, C.D., 1989. Dietary boron modified the effects of magnesium and molybdenum on mineral metabolism in the vitamin D3-deficient chick. Biol. Trace Elem. Res., 22: 201-220.
PubMedDirect Link - Ku, W. and R. Chapin, 1994. Mechanism of the testicular toxicity of boric acid in rats: In vivo and in vitro studies. Environ. Health Perspect., 102: 99-105.
PubMed - Ku, W.W., R.E. Chapin, R.F. Moseman, R.E. Brink, K.D. Pierce and K.Y. Adams, 1991. Tissue disposition of boron in male Fischer rats. Toxicol. Applied Pharmacol., 11: 145-151.
CrossRef - Kurtoglu, F., V. Kurtoglu, I. Celik, T. Kececi and M. Nizamlıoglu, 2005. Effects of dietary boron supplementation on biochemical parameters, peripheral blood lymphocytes, splenic plasma cells and bone characteristics of broiler chicks given diets with adequate or inadequate cholecalciferol (vitamin D3) content. Brit. Poult. Sci., 46: 87-96.
CrossRefPubMed - Kurtoglu, V., F. Kurtoglu and B. Coskun, 2001. Effects of boron supplementation of adequate and inadequate vitamin D3-containing diet on performance and serum biochemical characters of broiler chickens. Res. Vet. Sci., 71: 183-187.
CrossRefDirect Link - Moseman, R.F., 1994. Chemical disposition of boron in animals and humans. Environ. Health Persoect., 102: 113-117.
PubMed - Naghii, M.R. and S. Samman, 1993. The role of boron in nutrition and metabolism. Prog. Food Nutr. Sci., 17: 331-349.
PubMed - Nielsen, F.H., T.R. Schuler, T.J. Zimmerman and E.O. Uthus, 1988. Dietary magnesium, manganese and boron affect the response of rats to high dietary aluminum. Magnesium, 7: 133-147.
PubMed - Qin, X. and H. Klandorf, 1991. Effects of dietary boron supplementation on egg production, shell quality and calcium metabolism in aged broiler breeder hens. Poult. Sci., 70: 2131-2138.
PubMed - Rossi, A.F., R.D. Miles, B.L. Damron and L.K. Flunker, 1993. Effects of dietary boron supplementation on broilers. Poult. Sci., 72: 2124-2130.
PubMed - Sander, J.E., L. Dufour, R.D. Wyatt, P.B. Bush and R.K. Page, 1991. Acute toxicity of boric acid and boron tissue residues after chronic exposure in broiler chickens. Avian Dis., 35: 745-749.
Direct Link - See, A.S., A.B. Salleh, F.A. Bakar, N.Z. Yusof, A.S. Abdulamir and L.Y. Heng, 2010. Risk and health effect of boric acid. Am. J. Applied Sci., 7: 620-627.
CrossRef - Shaaban, M.M., 2010. Role of boron in plant nutrition and human health. Am. J. Plant Physiol., 5: 224-240.
CrossRefDirect Link - Tariq, M. and C.J.B. Mott, 2007. The significance of boron in plant nutrition and environment-A review. J. Agron., 6: 1-10.
CrossRefDirect Link - Treinen, K.A. and R.E. Chapin, 1991. Development of testicular lesions in F344 rats after treatment with boric acid. Toxicol. Appl. Pharm., 107: 325-335.
PubMed - Wilson, J.H. and P.L. Ruszler, 1996. Effects of dietary boron supplementation on laying hens. Br. Poult. Sci., 37: 723-729.
CrossRef - Wilson, J.H. and P.L. Ruszler, 1998. Long term effects of boron on layer bone strength and production parameters. Br. Poult. Sci., 39: 11-15.
PubMed - Woods, W.G., 1994. An introduction to boron: History, sources, uses and chemistry. Environ. Health Perspect., 102: 5-11.
PubMed - Yildiz, A.O., O. Olgun and Y. Cufadar, 2011. Effects of boron supplementation to diet on performance and boron deposition in broilers. Arch. Zoo., 14: 32-36.
Direct Link - Laila, E.A.N. and E.A.A. Adel, 2002. Effect of boron deficiency on some physiological and biochemical aspects during the developmental stages of wheat (Triticum aestivum L.) plant. J. Biol. Sci., 2: 470-476.
CrossRefDirect Link - Bustos-Obregon, E. and B. Ricardo Hartley, 2008. Ecotoxicology and testicular damage (environmental chemical pollution). A review. Int. J. Morphol., 26: 833-840.
CrossRefDirect Link - Geyikogglu, F. and H. Turkez, 2007. Acute toxicity of boric acid on energy metabolism of the breast muscle in broiler chickens. Biologia, 62: 112-117.
CrossRef - Ince, S., H. Keles, M. Erdogan, O. Hazman and I. Kucukkurt, 2011. Protective effect of boric acid against carbon tetrachloride-induced hepatotoxicity in mice. Drug Chem. Toxicol.
CrossRef