
Kagan Kokten
Department of Field Crops, Faculty of Agriculture, Bingol University, Bingol, Turkey
Alpaslan Kocak
Department of Biology, Faculty of Science and Arts, Bingol University, Bingol, Turkey
Mahmut Kaplan
Department of Field Crops, Faculty of Agriculture, Erciyes University, Kayseri, Turkey
Mevlut Akcura
Department of Field Crops, Faculty of Agriculture, Bing�l University, Bing�l, Turkey
Adil Bakoglu
Program of Field Crops, Vocational School of Higher Education,Bingol University, Bingol, Turkey
Eyup Bagci
Department of Biology, Faculty of Science and Arts, Firat University, Elazig, Turkey
Asian Journal of Animal and Veterinary Advances
Year: 2011 | Volume: 6 | Issue: 1 | Page No.: 88-95







ABSTRACT
The seed oils of seven Trifolium species (Leguminosae) were investigated for their protein, tannin contents and fatty acid compositions. The fatty acid compositions of these seven different species were determined by gas chromatography of the methyl esters of their fatty acids. The seed protein contents were found to be between 23.57 and 36.60%. The seed tannin contents were found to be between 0.77 and 1.73%. The seed oils of Trifolium species contain palmitic and stearic acids as the major component fatty acids, among the saturated acids, with a small amount of myristic, pentadecanoic, arachidic and behenic acids. The major unsaturated fatty acids found in the seed oils were oleic, linoleic and linolenic acids. In this study, Total Saturated Fatty Acids (TSFA) of Trifolium species were between 18.5 and 24.4% while Total Unsaturated Fatty Acids (TUSFA) were between 71.1 and 80.3%. The results revealed that the seed oils of Trifolium species studied with a substantial amount of very long chain fatty acids might have attracted attention because of their value of nutritional, industrial and renewable resources.
PDF Abstract XML References Citation
How to cite this article
Kagan Kokten, Alpaslan Kocak, Mahmut Kaplan, Mevlut Akcura, Adil Bakoglu and Eyup Bagci, 2011. Tannin, Protein Contents and Fatty Acid Composition of the Seeds of Some Trifolium L. species from Turkey. Asian Journal of Animal and Veterinary Advances, 6: 88-95.
DOI: 10.3923/ajava.2011.88.95
URL: https://scialert.net/abstract/?doi=ajava.2011.88.95
DOI: 10.3923/ajava.2011.88.95
URL: https://scialert.net/abstract/?doi=ajava.2011.88.95
INTRODUCTION
The genus Trifolium (Fabaceae or Leguminosae) is distributed in temperate and subtropical regions of both hemispheres (Bisby et al., 1994). The Mediterranean region is very rich in Trifolium species. Especially, in Turkey where it is widely spread and represented by 103 species (Maxted et al., 1997).
Legumes are an important food source and play a significant role in traditional diets in many regions of the world. Among the legume seeds, some are used as vegetables and others as supplementary sources of protein in animal diets (Savage, 1988). The widespread use of legumes makes this food group an important source of lipid and fatty acids in animal and human nutrition. Some publications dealing with the total lipid and fatty acid composition are reviewed by a few researchers (Welch and Griffiths, 1984; Grela and Gunter, 1995; Bakoglu et al., 2009, 2010; Yoshida et al., 2009).
The possible role of forage legumes in modern livestock production is being reconsidered (Wachendorf et al., 2006). Greater use of these species in pastures provides economic and agronomic benefits to agropastoral systems (Rochon et al., 2004). Biological nitrogen fixation by legumes results in high protein herbage and improved nutrient balance of the sward (Pecetti et al., 2009). Legume-based pastures may increase the self provision of the protein sources, the feeding value of forages and sustainability of grazing systems (Pflimlin et al., 2003).
Grain legumes contain moderately high levels of protein and amino acids and are promising alternatives (Wiseman, 1987; Brand et al., 1995). An accurate assessment of the profile and percentage total fatty acids from fresh forage is crucial when studying the biohydrogenation of fatty acid from fresh plants (Ribeiro and Eastridge, 2006). After harvesting, loss of lipids in forages occurs due to the oxidation of unsaturated fatty acids through the activity of plant lipases (Fried et al., 1993) and lipoxygenases (Gardner, 1991), decreasing the amount of unsaturated fatty acids (Dewhurst et al., 2002).
Tannins are general toxins. In addition, tannins act as feeding repellents to a great diversity of animals (Teiz and Zeiger, 2002). Condensed tannins, also known as flavolans, are polymers of flavanols (Sarkar et al., 1976) and are believed to have a major role in conferring bloat safeness on forage legumes by complexing with proteins. This renders proteins incapable of supporting foam (Jones et al., 1995). Astragalus cicer, for example, does not contain tannins, but is bloating safe (Jhonston et al., 1971), although it produces a greater volume of foam in vitro than Medicago sativa or Trifolium pratense (Cooper et al., 1966).
The objective of the present study was to determine the tannin and protein contents of the seeds of some Trifolium L. species (T. alexandrinum L., T. spumosum L., T. resupinatum L., T. lappaceum L., T. campastre Scrheb., T. angustifolium L. and T. purpureum Loisel.). In addition during the course of this study, it was aimed to characterize seed fatty acids used by animals in field, to establish the nutritional value and to do contributions as the renewable resources of Fatty Acids (FA) and other chemical patterns in these crops.
MATERIALS AND METHODS
Seed samples: The Trifolium species used in this study were T. alexandrinum L., T. spumosum L., T. resupinatum L., T. lappaceum L., T. campastre Scrheb., T. angustifolium L. and T. purpureum Loisel. Matured seeds of these species were collected from various locations in Adana (Turkey) between June and August 2009.
Oil extraction and preparation of Fatty Acid Methyl Esters (FAME): Impurities were removed from the seeds and the cleaned seeds were ground into powder using a ball mill. Lipids were extracted with hexane/isopropanol 2 v/v (Hara and Radin, 1978). The lipid extracts were centrifuged at 10.0 g for 5 min and filtered; then the solvent was removed on a rotary evaporator at 40°C.
Capillary GLC: Fatty acids in the lipid extracts were converted into methyl esters by means of 2% sulphuric acid (v/v) in methanol (Christie, 1990). The fatty acid methyl esters were extracted with hexane. Then the methyl esters were separated and quantified by gas chromatography and flame ionization detection (Schimadzu GC, 17 Ver.3) coupled to a glass GC 10 software computing recorder. Chromatography was performed with capillary column (25 m in length) and 0.25 mm in diameter, Permabound 25, Machery-Nagel, Germany using nitrogen as carrier gas (flow rate 0.8 mL min-1). The temperatures of the column, detector and injector valve were 130-220 and 240-280°C, respectively. Identification of the individual method was performed by frequent comparison with authentic standard mixtures that were analyzed under the same conditions.
Determination of protein and tannin contents: The extracts were prepared in 70% aqueous acetone (v/v). Total phenols were measured by using the Folin-Ciocalteu method and the results are expressed as tannic acid (Merck; Darmstadt, Germany) equivalent. Protein precipitation capacity of tannins was measured by the method of Makkar et al. (1995). For the protein precipitation assay, seed extracts were prepared in 50% aqueous methanol as presence of acetone intereferes with these assays. Nitrogen content (Crude protein = Nx6.25) in the seeds was determined by Kjeldahl method (AOAC, 1990). Protein and tannin analyses were carried out in triplicate.
Statistical analysis: The experimental design was completely randomized design with 3 replications. Data were analyzed SAS packet program. Biplot analysis was done to explain between fatty acids and Trifolium species by the method of Yan and Kang (2003).
RESULTS AND DISCUSSION
In this study, the total protein amount, fatty acid composition and tannin contents of some Trifolium species from Turkey were determined. The results of the fatty acid analysis are shown in Table 1 and Fig. 1 and total protein and tannin contents in Table 2.
The fatty acid composition of the seeds: The fatty acid composition of some plants used as feed crops from Fabaceae family showed different saturated and unsaturated fatty acid concentrations. The main components in the seed oils of Trifolium are linoleic, oleic, palmitic and linolenic acids. In all Trifolium studied found linoleic acid as major component (47.16-60.82%). Oleic acid content was at the highest level in T. spumosum (23.19%), but found to be at the lowest level in T. lappaceum (13.58%) and T. alexandrianum (12.86%). Linoleic acid was found in the greatest proportion in the seed oil. Linoleic acid content was at the highest level in T. lappaceum (60.82%), but found to be at the lowest level in T. alexandrianum (47.16%). The seed oils of all the species were richer in linoleic than linolenic acid contents. Linolenic acid was also detected at low level in T. campestre (4.0%) (Table 1).
| Table 1: | Fatty acid composition of some Trifolium species from Turkey |
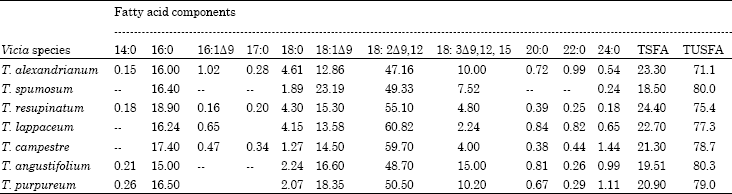 | |
| 14:0: myristic acid, 16:0: palmitic acid, 16:1Δ9: palmitoleic acid, 17:0: margaric acid, 18:0: stearic acid, 18:1Δ9: oleic acid, 18:2Δ9,12: linoleic acid, 18:3Δ9,12, 15: linolenic acid, 20:0: arachidic acid, 22:0: behenic acid, 24:0: lignoceric acid, TSFA: Total saturated fatty acid, TUSFA: Total unsaturated fatty acid | |
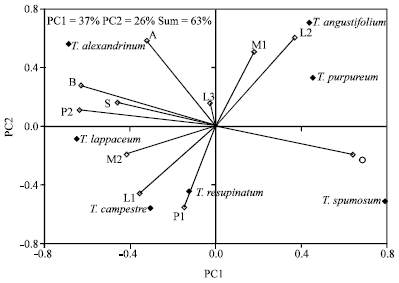 | |
| Fig. 1: | The biplot of 7 Trifolium species for fatty acid composition. Fatty acid: M1: Myristic acid, P1: Palmitic acid, P2: Palmitoleic acid, M2: Margaric acid, S: Stearic acid, O: Oleic acid, L1: Linoleic acid, L2: Linolenic acid, A: Arachidic acid, B: Behenic acid, L3: Lignoceric acid) |
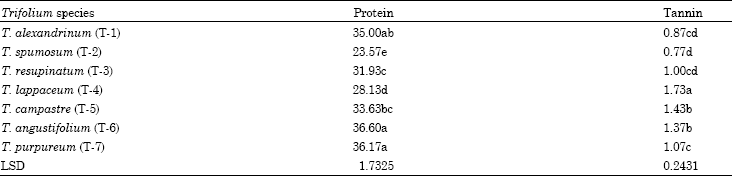
| Table 2: | Total protein (%) and tannin contents (%) of some Trifolium species from Turkey |
 | |
The oil of the five Trifolium species (T. balansae, T. stellatum, T. nigrescens subsp. petrisavii, T. constantinopolitanum and T. resupinatum var. resupinatum) was reported a similar fatty acid profile. The major fatty acids were linolenic acid (16.6-31.1%), linoleic acid (5-11.3%) and palmitic acid (11.1-18.3%) (Sabudak et al., 2009).
Linolenic acid was found to be the predominant free fatty acid and triacontanol was determinate as the principal free fatty alcohol in T. repens. The hydrocarbons of C29 and C31 were observed in the largest amount in T. repens (Body, 1974; Maffei, 1996). Also, linolenic acid (61.5%) was found the major component in T. resupinatum (Nazir and Shah, 1990).
Behenic acid (22:0) was not found (T. spumosum) or it was in very low amounts in the legume seed oils studied, T. alexandrianum (0.99%), T. resupinatum (0.25%), T. lappaceum (0.82%), T. campestre (0.44%), T. angustifolium (0.26%) and T. purpureum (0.29%). The low amounts behenic acid in legume seed oils is important because of some researchers have indicated that oils with have high levels of behenic acid may be difficult for digestive enzymes in humans and animals (Hilditch and Williams, 1964; Balogun and Fetuga, 1985). Despite its low bioavailability compared with oleic acid, behenic acid is a cholesterol-raising fatty acid in humans and therefore not a suitable for palmitic acid in manufactured triacylglycerols (Cater and Denke, 2001).
We have used the biplot to compare Trifolium species on the basis of fatty acid compositions and to identify species or groups of Trifolium that are particularly good in certain aspects (Fig. 1).
The GT biplot (Fig. 1) displays 63% of the information in the standardized data of the 7 Trifolium species for 11 fatty acids, which is partially presented in Table 1. This biplot can be visualized from two perspectives. First, it shows the associations among the traits across the 7 Trifolium species: (1) a positive correlation (acute angle) between linolenic acid and myristic acid; between arachidic acid, behenic acid, stearic acid and palmitoleic acid; between margaric acid, linoleic acid and palmitic acid (2) a negative correlation (obtuse angle) between oleic acid and behenic acid, stearic acid and palmitoleic acid and (3) a negative correlation (obtuse angle) between linolenic acid and : linolenic acid. Similar relationships among some legumes fatty acid composition of were reported by Kokten et al. (2010). Second, it shows the trait profiles of the Trifolium species, particularly those that are placed farther away from the biplot origin. For example, it shows that T. angustifolium had extremely high linolenic acid and high myristic acid but low palmitic acid, linoleic acid and margaric acid; T. spumosum had extremely high oleic acid but lower levels for other fatty acits and T. alexandrinum had high behenic acid, T. resupinatum had highest palmitic acid, T. campestre had higher linoleic acid (Table 1, Fig. 1).
Total Saturated Fatty Acid (TSFA) of Trifolium species were between 18.5 and 24.4%. T. spumosum has lowest level of saturated acid and T. resupinatum has highest amount of saturated fatty acid concentration (Table 1). On the other hand, the unsaturated fatty acid composition of Trifolium species were determined as high levels reported as other family members of Fabaceae (Bagci et al., 2004a; Masoud, 2009; Bakoglu et al., 2010), Lamiaceae (Bagci et al., 2004b; Bagci, 2007), Boraginaceae (Bagci et al., 2004c), Apiaceae (Bena et al., 1998) family patterns. T. angustifolium has highest level of unsaturated fatty acid (80.3%) and also T. spumosum (80.0%), T. purpureum (79.0%), T. campestre (78.7%), T. lappaceum (77.3%), T. resupinatum (75.4%) and T. alexandrianum (71.1%), respectively. Medicago sativa and M. lupiluna (Fabaceae) has 83.46 and 78.55% unsaturated fatty acid concentrations in their seed oils (Bakoglu et al., 2010).
Total protein amounts of the seeds: Total protein amounts of feed crops studied were between 23.57-36.60% in T. spumosum and T. angustifolium and 28.13, 31.93, 33.63, 35.00 and 36.17% in T. lappaceum, T. resupinatum, T. campastre, T. alexandrinum and T. purpureum, respectively. In the Bakoglu et al. (2009) study, the protein levels of T. pratense and T. aureum were determined which belong to seeds. According to the authors, the protein levels studied were found between 26.1 and 28.6% (Table 2). Many previous studies have reported the changes in protein contents for other forage legumes (Ukhun and Ifebigh, 1988; Bena et al., 1998; Djemel et al., 2005; Maheri-Sis et al., 2007; Bakoglu et al., 2009; Ciftci and Bagci, 2009).
Tannin contents of the seeds: Tannin amounts of feed crops studied were between 0.77-1.73% in Trifolium spumosum and Trifolium lappaceum and 0.87, 1.00, 1.07, 1.37 and 1.43% in, T. alexandrinum, T. resupinatum, T. purpureum, T. angustifolium and T. campastre, respectively (Table 2). Tannins may play a major role in limiting proteolysis in some legumes during ensiling (Albrecht and Muck, 1991), but it would appear that an enzyme, possibly a polyphenol oxidase, may act to inhibit proteolysis in red clover (Jones et al., 1995; Hatfield and Muck, 1999) and may account for the Crude Protein (CP) in red clover (T. pratense) being less degradable than that in Lucerne, both in the silo and in the rumen. Grain legumes are potential sources of energy and micronutrients but their use is still limited because of in certainty about the amount and the effect of antinutritional factors they may contain (Wiryawan and Dingle, 1999).
CONCLUSION
The oil contents of the studied legumes belonging to the Trifolium genus, showed quantitative differences but the seed oils showed uniform fatty acid composition. The results of Trifolium patterns in this study, its chemotaxonomy are noteworthy. Total protein contents have found high level for studied of Trifolium species. By whatever mechanism protein-tannin binding occurs, this process has a negative impact on herbivore nutrition. Tannins can inactivate herbivore digestive enzymes and create complex aggregates of tannins and plant proteins that are difficult to digest. The results revealed that the seed oils of Trifolium species studied with a substantial amount of very long chain fatty acids might have attracted attention because of their value of nutritional, industrial and renewable resources.
REFERENCES
- Albrecht, K.A. and R.E. Muck, 1991. Proteolysis in ensiled forage legumes that vary in tannin concentration. Crop Sci., 31: 464-469.
Direct Link - Bagci, E., 2007. Fatty acids and tocochromanol patterns of some Turkish Apiaceae (Umbelliferae) plants: A chemotaxonomic approach. Acta Botancia Gallica, 154: 143-152.
Direct Link - Bagci, E., L. Bruehl, H. Ozcelik, K. Aitzetmuller, M. Vural and A. Sahin, 2004. A study of the fatty acid and tocochromanol patterns of some Fabaceae (Leguminosae) plants from Turkey I. Grassas Y Aceites, 55: 378-384.
Direct Link - Bagci, E., L. Bruehl, K. Aizetmuller, Y. Altan, 2004. Fatty acid and tocochromanol patterns of some Turkish Boraginaceae: A chemotaxonomic approach. Nordic J. Bot., 22: 719-726.
CrossRef - Bagci, E., M. Vural, T. Dirmenci, L. Bruehl and K. Aitzetmuller, 2004. Fatty acid and tocochromanol patterns of some Salvia L. (Lamiaceae) sp. from Turkey. Zeitschrift Fur Naturforsch, 59: 305-309.
Direct Link - Bakoglu, A., E. Bagci, A. Kocak and E. Yuce, 2010. Fatty acid composition of some Medicago L. (Fabaceae) species from Turkey. Asian J. Chem., 22: 651-656.
Direct Link - Bakoglu, A., E. Bagci and H. Ciftci, 2009. Fatty acid, protein contents and composition of some feed crops from Turkey. J. Food Agric. Environ., 7: 343-346.
Direct Link - Balogun, A.M. and B.I. Fetuga, 1985. Fatty acid composition of seed oils of some members of the leguminosae family. Food Chem., 17: 175-182.
CrossRef - Bena, G., J.M. Prosperi, B. Lejeune and I. Olivieri, 1998. Evolution of annual species of the genus Medicago: A molecular phylogenetic approach. Mol. Phisiogenet. Evol., 9: 552-559.
CrossRef - Body, D.R., 1974. Neutral lipids of leaves and steams of Trifolium repens. Phytochemistry, 13: 1527-1530.
CrossRef - Brand, T.S., R.C, Olckers and J.P. Van der Merwe, 1995. Evaluation of faba beans (Vicia faba cv Fiord) and sweet lupins (Lupinus albus cv Kiev) as protein source for growing pigs. South Afr. J. Anim. Sci., 25: 31-35.
Direct Link - Cater, N.B. and M.A. Denke, 2001. Behenic acid is a cholesterol-raising saturated fatty acid in humans 1-3. Am. J. Clin. Nutr., 73: 41-44.
Direct Link - Ciftci, H. and E. Bagci, 2009. Protein contents and metal composition of some feed crops from Turkey. Int. J. Chem. Tech. Res., 1: 577-580.
Direct Link - Cooper, C.S., R.F. Eslick and P.W. McDonald, 1966. Foam formation from extracts of 27 legume species in vitro. Crop Sci., 6: 215-216.
Direct Link - Dewhurst, R.J., J.M. Moorby, M.S. Dhanoa and W.J. Fisher, 2002. Effect of level concentrate feding during the second gestation of Holstein-Friesian dairy cows. 1. Feed intake and milk production. J. Dairy Sci., 85: 169-177.
Direct Link - Djemel, N., D. Guedon, A. Lechevalier, C. Salon and M. Miquel et al., 2005. Development and composition of the seeds of nine genotypes of the Medicago truncatula species complex. Plant Phys. Biochem., 43: 557-566.
CrossRef - Fried, A., D. Benayahu and S. Wientroub, 1993. Marrow stroma derived osteogenic clonal cell lines: Putative stages in osteoblastic differentiation. J. Cell Physiol., 155: 472-482.
CrossRef - Gardner, S.M., 1991. Ground beetle (Coleoptera, Carabidae) communities on upland heath and their association with heathland flora. J. Biogeogr., 18: 281-289.
Direct Link - Grela, E.R. and K.D. Gunther, 1995. Fatty acid composition and tocopherol content of some legume seeds. Anim. Feed Sci. Technol., 52: 325-331.
Direct Link - Hedayatifard, M., 2009. Comparative study of fatty acid composition of golden mullet fillet and roe oil (Liza aurata Risso, 1810). Aian J. Anim. Vet. Adv., 4: 209-213.
CrossRefDirect Link - Jones, B.A., R.E. Muck and R.D. Hatfield, 1995. Red clover extracts inhibit legume proteolysis. J. Sci. Food Agric., 67: 329-333.
Direct Link - Maheri-sis, N., A. Mirzaei-Aghsaghali, A. Mirza-Aghazadeh and A.R. Safaei, 2007. Determination of digestibility and nutritive value of Iranian alfalfa varieties using in vivo technique in sheep. Asian. J. Anim. Vet. Adv., 2: 239-243.
CrossRefDirect Link - Nazir, M. and F.H. Shah, 1990. Stability of fatty acids and leaf protein concentrate of Persian clover (Trifolium resupinatum). Plant Foods Hum. Nutr., 40: 283-288.
CrossRef - Pecetti, L., P. Annicchiarico, F. Battini and S. Cappelli, 2009. Adaptation of forage legume species and cultivars under grazing in two extensive livestock systems in Italy. Eur. J. Agron., 30: 199-204.
CrossRefDirect Link - Pflimlin, A., J.D. Arnaud, D. Gautier and A. Le Gall, 2003. Forage legumes, a means to conciliate protein self-sufficiency and preservation of the environment. Fourrages, 174: 183-204.
Direct Link - Ribeiro, C.V.D.M. and M.L. Eastridge, 2006. Short communication: Effect of number of extractions on percentage of long-chain fatty acids from fresh alfalfa. J. Dairy Sci., 89: 3505-3507.
Direct Link - Rochon, J.J., C.J. Doyle, J.M. Greef, A. Hopkins and G. Molle et al., 2004. Grazing legumes in Europe: A review of their status, management, benefits, research needs and future prospects. Grass Forage Sci., 59: 197-214.
CrossRefDirect Link - Sabudak, T., M. Ozturk, A.C. Goren, U. Kolak and G. Topcu, 2009. Fatty acids and other lipid composition of five Trifolium species with antioxidant activity. Pharm. Biol., 47: 137-141.
CrossRef - Sarkar, S.K., R.E. Howarth and B.P. Goplen, 1976. Condensed tannins in herbaceous legumes. Crop Sci., 16: 543-546.
Direct Link - Teiz, L. and E. Zeiger, 2002. Plant Physiology. 3rd Edn., Sinauer Associates, Sunderland, Massachusetts, ISBN: 9780878938230, Pages: 690.
Direct Link - Ukhun, M.E. and E.O. Ifebigh, 1988. Compositional chemistry of Cassia alata seeds. Food Chem., 30: 205-210.
CrossRef - Welch, R.W. and D.W. Griffiths, 1984. Variation in the oil content and fatty acid composition of field beans (Vicia faba) and peas (Pisum spp.). J Sci. Food Agric., 35: 1282-1289.
CrossRef - Wiryawan, K.G. and J.G. Dingle, 1999. Recent research on improving the quality of grain legumes for chicken growth. Anim. Feed. Sci. Technol., 76: 185-193.
CrossRefDirect Link - Yoshida, H., M. Saiki, N. Yoshida, Y. Tomiyama and Y. Mizushina, 2009. Fatty acid distribution in triacylglycerols and phospholipids of broad beans (Vicia faba). Food Chem., 112: 924-928.
CrossRef - Makkar, H.P.S., M. Blummel and K. Becker, 1995. Formation of complete between Polyvinyl pyrolidones or polyethylene glycol and tannins and their implication in gas production and true digestibility in in vitro technique. Br. J. Nutr., 73: 897-913.
CrossRef