
Jing Liu
Key Laboratory of Food Nutrition and Safety, Ministry of Education, Tianjin University of Science and Technology, Tianjin 300457, China
Ying Miao
Key Laboratory of Food Nutrition and Safety, Ministry of Education, Tianjin University of Science and Technology, Tianjin 300457, China
Zheng Zhao
Key Laboratory of Food Nutrition and Safety, Ministry of Education, Tianjin University of Science and Technology, Tianjin 300457, China
Wenbo Zhao
Key Laboratory of Food Nutrition and Safety, Ministry of Education, Tianjin University of Science and Technology, Tianjin 300457, China
Asian Journal of Animal and Veterinary Advances
Year: 2011 | Volume: 6 | Issue: 12 | Page No.: 1214-1223







ABSTRACT
Protein and peptides are popular supplements in sports nutrition which is most often used to increase the exercise performance of human and animals. The antioxidant capacities and antifatigue properties of whey hydrolysates fermented by Lactobacillus plantarum TUST200209 and its purified whey peptides were studied. The treatment groups were orally administered with high dose (1.5 g/kg/day) and low dose (0.5 g/kg/day) of whey hydrolysates and its purified peptides in Kunming mice for four weeks, respectively. High dose (1.5 g/kg/day) of 0.9% saline (vehicle group) and mixture of whey and Lactobacillus plantarum TUST200209 were used as control. Compared with vehicle group, whey hydrolysates and its purified peptides could significantly prolong the swimming time to fatigue (p<0.05) and increase levels of superoxide dismutase and glutathione peroxidase. Moreover, contents of malondialdehyde in red cell, quadriceps femoris and mitochondria decreased and most of the changes were significant (p<0.05) for the high dose of whey peptides. The high antioxidant capacities were consistent with high antifatigue ability. Compared with vehicle group, the group oral administered high dose of whey peptides demonstrated high levels of glucose, non-esterified fatty acid and liver glycogen (p<0.05). It demonstrated that the whey hydrolysates could significantly alleviate exercise fatigue of the mice induced by free radical, especially for its purified peptides.
PDF Abstract XML References Citation
Received: August 21, 2011;
Accepted: October 01, 2011;
Published: November 05, 2011
How to cite this article
Jing Liu, Ying Miao, Zheng Zhao and Wenbo Zhao, 2011. Retard of Whey Hydrolysates Supplementation on Swimming Exercise-induced Fatigue in Mice. Asian Journal of Animal and Veterinary Advances, 6: 1214-1223.
DOI: 10.3923/ajava.2011.1214.1223
URL: https://scialert.net/abstract/?doi=ajava.2011.1214.1223
DOI: 10.3923/ajava.2011.1214.1223
URL: https://scialert.net/abstract/?doi=ajava.2011.1214.1223
INTRODUCTION
According to Harmans, classical radical theory, intense exercise caused homeostasis disruptions between the body’s oxidation system and its antioxidation system (Harman, 1956). During swimming exercise, the oxygen consumption increased tremendously, 20-fold for mitochondria (Ji and Leichtweis, 1997) and 100-200 times for muscle (Atamaniuk et al., 2004) which caused production of Reactive Oxygen Species (ROS). The accumulation of reactive free radical was considered responsible for muscle fatigue during exercise (Powers et al., 2004) which limited exercise performance. Hence oxidative stress plays a significant role in the initiation and progression of fatigue (Tharakan et al., 2005). Malondialdehyde (MDA), a product of lipid peroxidation, increased during the strenuous exercise causing physical fatigue (Richards et al., 2007). Intracellular antioxidant enzymes, including superoxide dismutase (SOD) and glutathione peroxidase (GSH-Px), could reduce oxidative stress mediated body fatigue (Singh et al., 2002). The antifatigue function could be demonstrated by the improvement of plasma biochemical parameters and storage of glycogen (Prasad and Khanum, 2012).
In food and Pharmaceutical industry, there has been considerable interest in how to look for the natural materials, especially for proteins and peptides, to inhibit production of free radical and postpone the sports fatigue (Ali, 2011; Geethalakshmi et al., 2010; Prasad and Khanum, 2012). Proteins and peptides injected intraperitoneally or oral administered into mice decreased levels of blood urea nitrogen and lactic acid and increased swimming endurance and the storage of liver glycogen (Wang et al., 2008; Takenaka et al., 2003; Elia et al., 2006). Peptides were superior to the protein (Ren et al., 2011; Yu et al., 2008).
Although, many researches focused on the enhanced capacity of bioactive peptides on antioxidant activity or exercise-induced fatigue (Kamau and Lu, 2011; Ren et al., 2011) the direct relationship between the antioxidant capacity and antifatigue ability of whey hydrolysates was seldom studied (Misra et al., 2006). Whey protein is the main factor that was responsible for the antioxidant activity of whey (Tong et al., 2000). Antioxidant properties of whey protein could increase by hydrolysis with proteinase (Hernandez-Ledesma et al., 2005). But, if antioxidant ability and antifatigue capacity were improved by whey hydrolysates fermented by lactic acid bacterial and its purified peptides was not reported. Addition of Lactobacillus plantarum TUST200209 could increase the antioxidant capacity of whey hydrolysates. But the antifatigue function and relationship between antifatigue and antioxidation were not studied. In this paper, after oral administration of whey hydrolysates and peptides to mice, swimming time, activity of antioxidant enzymes, plasma biochemical parameters and storage of glycogen were explored.
MATERIALS AND METHODS
Raw milk was purchased from local farm. Chymotrypsin was procured from Novo Nordisk, Denmark in 2010. Lactobacillus plantarum TUST200209 was stored by Laboratory of Cheese Science and Technology, Tianjin University of Science and Technology, Tianjin, China. Kunming mice were provided by the animal center of Academy of Military Sciences, China People’s Liberation Army in 2011. Commercial kits to determine the levels of SOD, GSH-Px, MDA, glucose, lactate, non-esterified fatty acid (NEFA), blood urine nitrogen (BUN) and glycogen content were got from Nanjing Jiancheng Bioengineering Center, China in 2011.
Whey hydrolysates preparation: Whey was made using raw skim milk via adjusting pH to 4.6, removal of casein by centrifugation (3000 rpm, 10 min), readjusting to pH 7.0 and recentrifugation (3000 rpm, 10 min). Whey was condensed by vacuum rotary evaporation to 10 mg protein/mL. Chymotrypsin was used to hydrolyze whey and the enzymolysis condition was: temperature 37°C, the ratio of enzyme/substrate was 2:100. After 2 h, solution was pasteurized. Lactobacillus plantarum TUST200209 was added to the solution 3% (v/v) and fermented 24 h. The Whey Hydrolysates (WH) were lyophilized and stored at-80°C until further use. Macroporous resin was used to purify WH to remove lactose, lactic acid and salt. Chromatography column 2.5x30 cm; Resin DA201-C; adjusted pH 6.0, the sample flow rate 0.75 BV/h and deionized water were used to elute lactose, lactic acid and salt. Seventy five present ethanol elution flow rate was 0.5 BV/h and absorbance of elution was measured at 220 nm with a UV detector. Elution peak materials, whey peptides (WP), were collected and evaporated to get rid of ethanol. WP was lyophilized and stored at-80°C until further use.
Animals: Four-week-old male Kunming mice were used. They were housed in standard cages (21.5x32x14, 5 mice/cage) under controlled conditions of temperature (24±1°C), humidity (about 60%) and 12 h light-dark cycle. They were given free access to water and laboratory standard diet. The care and treatment of experimental animals conformed to the guidelines for the ethical treatment of laboratory animals, Principles of Laboratory Animal Care (WHO, 1985). The mice were allowed to adapt to the laboratory housing for at least 1 week.
Sample and mice treatments: The forty mice were divided into six groups (n = 8 per group). The mice were given different oral administered treatments: (1) vehicle (0.9% saline, 1.5 g kg-1 body weight); (2) low dose of WH (WHL, 0.5 g kg-1 body weight); (3) high dose of WH (WHH, 1.5 g kg-1 body weight); (4) low dose of WP (WPL, 0.5 g kg-1 body weight); (5) high dose of WP (WP, 1.5 g kg-1 body weight) and (6) mixture of whey and Lactobacillus plantarum TUST200209 (MAL, 1.5 g kg-1 body weight), respectively. All the substrates above were dissolved in 0.9% saline and were oral administered in doses of 200 μL every day for four weeks.
Swimming endurance protocol: The method was used according to Bostrom et al. (1974) and Kamakura et al. (2001) and a slight modification was made. The swimming capacity of the mice was measured using an acrylic plastic pool (90x45x45 cm) filled with water to a depth of 35 cm. The temperature of the water was maintained at 34±1°C. The mice were loaded with a lead sheath, weighing approximately 8% of their body weight, attached to their tails. The mice were determined to be fatigued when they failed to rise to the surface to breathe within a 5 sec period. The swimming exercise was performed the seventh day every week. To avoid circadian variations in physical activity, swimming exercise was performed between 11:00 and 17:00 which has been confirmed the minimal variation on endurance capacity in mice. The swimming time to fatigue was used as the index of the forced swimming capacity. At the last day of the 4th week, the mice were killed immediately after the swimming exercise for further research.
Analysis of SOD, GSH-Px and MDA in red cell and tissues: The mice were killed by removing the eyeballs and collecting blood after 12 h fasting, red cell was collected from blood (Miki et al., 1987). Quadriceps femoris and liver were quickly dissected out and mitochondria was separated from the liver, frozen in liquid nitrogen and kept at -70°C until further analysis. The levels of SOD, GSH-Px and MDA were analyzed with commercial kits.
Analysis of plasma biochemical parameters: Whole blood samples were collected in heparinized tubes. Plasma was prepared by centrifugation at 1000 rpm at 4°C for 10 min and then stored at -70°C (Ikeuchi et al., 2006; Jung et al., 2007). The levels of glucose, lactate, Non-Esterified Fatty Acid (NEFA) and Blood Urine Nitrogen (BUN) were analyzed with commercial kits.
Analysis of tissue glycogen contents: Immediately after the blood was collected, the liver and gastrocnemius muscles were quickly dissected out, frozen in liquid nitrogen and kept at -70°C until an analysis for glycogen content. The glycogen content was measured with commercial kits.
Statistical analysis: Date was presented as mean value with standard deviations. The SPSS 11.5 software (SPSS Inc., Chicago, IL, USA) was used to make one-way ANOVA(S-N-K) analysis. In all the tests, the criterion for statistical significance was p<0.05.
RESULTS
Effect on swimming time to fatigue: Swimming times to fatigue for the WHH and WPH groups were observed to be longer (p<0.05) than the vehicle group at the third and fourth weeks but there were no significant differences (p>0.05) at the first and second weeks (Table 1). The maximum performance was observed in WPH group and swimming time increased from 8.7±2.8 min (vehicle group) to 19.9±2.1 min at 4th week. Other groups couldn’t prolong the swimming time significantly, except for the WPL group (14.9±1.5 min) at 4th week.
For the WHL, WHH, WTH and WPL groups, swimming times increased significantly only after three weeks (p<0.05). But for WPH group, swimming time increased significantly after two weeks (p<0.05) (Table 1).
Levels of SOD, GSH-Px and MDA in red cell and tissue: The level of SOD in WPH group (8627.3±976.2 U mg-1 pro) increased (p<0.05) in red cell of mice compared to other groups except for WHH group (7123.5±653.4 U mg-1 pro) but the content of GSH-Px didn’t change obviously (p>0.05). MDA in WHH (0.62±0.22 nmol mg-1 pro), WPL (0.51±0.09 nmol mg-1 pro) and WPH groups (0.42±0.14 nmol mg-1 pro) decreased significantly (p<0.05) but there were no significant differences (p>0.05) in WHL, MAL and vehicle groups (Table 2).
The levels of SOD and GSH-Px in quadriceps femoris of each group had no significant difference (p>0.05) between these groups and vehicle group and both WPL and WPH groups could decrease the content of MDA significantly (p<0.05). For WPH group, the level of MDA was 9.12±2.47 nmol mg-1 pro, about half of the vehicle group (16.87±3.45 nmol mg-1 pro) (Table 3).
Compared to the vehicle group, the levels of SOD and GSH-Px, in WH and WP groups could increase significantly (p<0.05) and the content of MDA decrease significantly (p<0.05) in mitochondria of mice. The highest levels of SOD and GSH-Px were 557.2±23.1 U mg-1 pro and 503.2±33.2 U mg-1 pro, respectively in WHH group. Compared with Vehicle group (146.3±26.8 nmol mg-1 pro), WP groups could effectively decrease the levels of MDA, 63.7±21.4 nmol mg-1 pro for WPL group and 62.8±19.7 nmol mg-1 pro for WPH group. The differences between high and low dose were not significant for all parameters (Table 4).
| Table 1: | Changes of swimming exercise time to fatigue in mice |
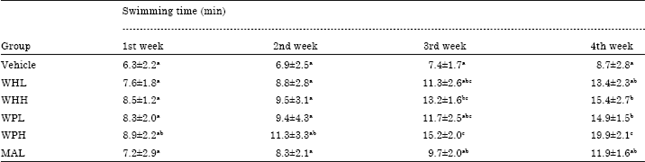 | |
| Data are Mean±SD values. Values in the same column with different letter superscripts indicate significant difference among groups (p<0.05). WHL, WHH: low dose (0.5 g kg-1 body weight) and high dose (1.5 g kg-1 body weight) of whey hydrolysates fermented by Lactobacillus plantarum TUST200209, WPL, WPH: low dose (0.5 g kg-1 body weight) and high dose (1.5 g kg-1 body weight) of whey peptides. MAL: High dose (1.5 g kg-1 body weight) of mixture of whey and Lactobacillus plantarum TUST200209 | |
| Table 2: | Levels of SOD, GSH-Px and MDA in red cell of mice after forced swimming to fatigue |
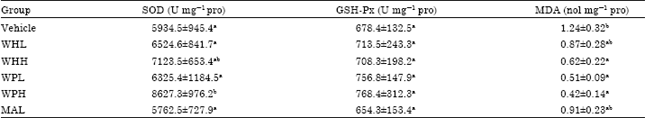 | |
| Data are Mean±SD values. Values in the same column with different letter superscripts indicate significant difference among groups p<0.05). WHL, WHH: low dose (0.5 g kg-1 body weight) and high dose (1.5 g kg-1 body weight) of whey hydrolysates fermented by Lactobacillus plantarum TUST200209, WPL, WPH: low dose (0.5 g kg-1 body weight) and high dose (1.5 g kg-1 body weight) of whey peptides. MAL: high dose (1.5 g kg-1 body weight) of mixture of whey and Lactobacillus plantarum TUST200209 | |
| Table 3: | Levels of SOD, GSH-Px and MDA in Quadriceps femoris of mice after forced swimming to fatigue |
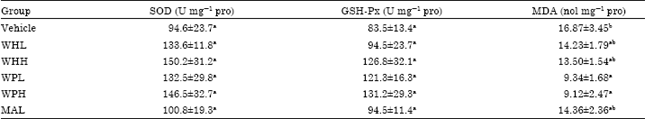 | |
| Data are Mean±SD values. Values in the same column with different letter superscripts indicate significant difference among groups (p<0.05). WHL, WHH: low dose (0.5 g kg-1 body weight) and high dose (1.5 g kg-1 body weight) of whey hydrolysates fermented by Lactobacillus plantarum TUST200209, WPL, WPH: low dose (0.5 g kg-1 body weight) and high dose (1.5 g kg-1 body weight) of whey peptides. MAL: high dose (1.5 g kg-1 body weight) of mixture of whey and Lactobacillus plantarum TUST200209 | |
| Table 4: | Levels of SOD, GSH-Px and MDA in mitochondria of mice after forced swimming to fatigue |
 | |
| Data are Mean±SD values. Values in the same column with different letter superscripts indicate significant difference among groups (p>0.05). WHL, WHH: low dose (0.5 g kg-1 body weight) and high dose (1.5 g kg-1 body weight) of whey hydrolysates fermented by Lactobacillus plantarum TUST200209, WPL, WPH: low dose (0.5 g kg-1 body weight) and high dose (1.5 g kg-1 body weight) of whey peptides, MAL: high dose (1.5 g kg-1 body weight) of mixture of whey and Lactobacillus plantarum TUST200209 | |
Levels of plasma glucose, lactate, NEFA and BUN: Plasma glucose was highest in the WPH group (129.8±11.8 mg dL-1) than in other group (p<0.05) and there is also significant difference between WPL, WHL, WHH groups and vehicle group, whereas no differences between MAL group (83.2±10.3 mg dL-1) and vehicle group (68.3±18.2 mg dL-1) (p>0.05) (Table 5). The levels of plasma lactate decreased in the WP, WH and MAL groups compared to the vehicle group but there was no significant difference (p>0.05) between these two parts (Table 5). In the WPH group (0.69±0.18 mEq L-1), plasma NEFA significantly increased compared with vehicle group (0.38±0.12 mEq L-1) (p<0.05) but for other group, the changes were not significant (p>0.05) (Table 5). In WHH group, BUN concentration was highest compared with other groups but there was no significant difference (p>0.05) between WPPH and vehicle group except for MAL group (342.8±42.5 mg L-1) (Table 5).
| Table 5: | Levels of plasma biochemical parameters of mice after forced swimming to fatigue |
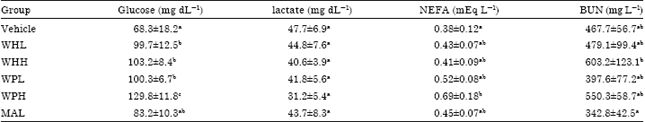 | |
| Data are Mean±SD values. Values in the same column with different letter superscripts indicate significant difference among groups (p<0.05), WHL, WHH: low dose (0.5 g kg-1 body weight) and high dose (1.5 g kg-1 body weight) of whey hydrolysates fermented by Lactobacillus plantarum TUST200209, WPL, WPH: low dose (0.5g kg-1 body weight) and high dose (1.5 g kg-1 body weight) of whey peptides, MAL: high dose (1.5 g kg-1 body weight) of mixture of whey and Lactobacillus plantarum TUST200209 | |
| Table 6: | Levels of liver and muscle glycogen of mice after forced swimming to fatigue |
 | |
| Data are Mean±SD values. Values in the same column with different letter superscripts indicate significant difference among groups (p<0.05). WHL, WHH: low dose (0.5 g kg-1 body weight) and high dose (1.5 g kg-1 body weight) of whey hydrolysates fermented by Lactobacillus plantarum TUST200209, WPL, WPH: low dose (0.5 g kg-1 body weight) and high dose (1.5 g kg-1 body weight) of whey peptides, MAL: high dose (1.5 g kg-1 body weight) of mixture of whey and Lactobacillus plantarum TUST200209 | |
Levels of liver and muscle glycogen: Oral administer different sample could not affect the levels of muscle glycogen significantly but could improve liver glycogen storage (p<0.05). The most effective material was WPH, then WHH and WPL and the levels of liver glycogen were 7.24±1.21, 6.12±1.03 and 6.25±2.15 mg g-1, respectively (Table 6).
DISCUSSION
Swimming times were significantly prolonged by administering whey hydrolysates which were dose-dependent and category-dependent. Compared with MAL group, WP and WH groups could prolong significantly swimming time, demonstrating that whey hydrolysates were more suitable than whey to relieve fatigue. Purification of whey hydrolysates could increase the content of protein in WPH group which was contributed to extending the swimming time of mice. Swimming time of WPH group was 29.2% longer than that of WHH group. Ren et al. (2011) found the similar result that grass carp peptide administration increased significantly the swimming endurance capacity compared with protein. Tong et al. (2000) divided whey into two parts, High-molecular-weight (HMW) fraction and low-molecular-weight (LMW) fraction. The HMW fraction represents the protein and phospholipid fraction of the whey, while the LMW fraction represents the lactose, mineral and other low-molecular-weight compounds. Because phospholipids had no effect on the oxidation, whey proteins are primarily responsible for the antioxidant activity of the HMW fraction. HMW fraction was the main antioxidative material compared with LMW fraction in whey, so the whey protein was the main factor that was responsible for the antioxidant activity of whey. It could be the main reason why purification of WH could significantly increase swimming time. The high antioxidant capacity meant the high scavenging activity of ROS which could delay fatigue induced by free radical. The longer swimming time was observed at high-dose oral administration and the WPH group demonstrated the longest swimming time.
Intracellular antioxidant enzymes, SOD and GSH-Px, could quickly remove oxygen free radicals in body and the activities of enzymes were weaker during fatigue. Administration of antioxidants, whey hydrolysates or whey peptides, reduced lipid peroxidation and restored or increased the levels of antioxidant enzymes. Singh et al. (2002) found the similar conclusion that several natural and synthetic antioxidants increased the content of SOD and catalase in mice. High levels of SOD and GSH-Px and low level of MDA in WPH group demonstrated that antioxidant enzyme could effectively scavenge oxygen free radicals to inhibit lipid peroxidation induced by free radicals. WHH and WPL didn’t increase significantly the levels of SOD and GSH-Px in red cell and quadriceps femoris but decreased significantly the content of MDA. It meant there was other mechanism to inhibit lipid peroxidation (Halliwell and Chirico, 1993). It was also the reason why levels of SOD or GSH-Px in all groups were not different significantly in Quadriceps femoris but content of MDA decreased significantly. Purification of Whey hydrolysates, whey peptides, could improve the antioxidative activity of whey hydrolysates, especially in red cell and mitochondria. For WPH group, the high levels of SOD and GSH-Px in red cell meant free radicals could be scavenged in time to avoid the harmful effect on cell which were contribute to maintain the relative stable circumstance in blood.
Antioxidant enzyme activities in skeletal muscle are one of the lowest organizations in body and free radicals in muscle can not be scavenged in time and muscle damage happened (Davies et al., 1982). Compared to the red cell, levels of SOD and GSH-Px in quadriceps femoris were low which would cause the increase of MDA.
Swimming exercise, led to hypoxia, affected mitochondrial function. The electron transportation associated with the mitochondrial respiratory chain is considered the major process leading to ROS production during exercise. At end stage of mitochondrial electron transport chain, the supply of oxygen to accept electrons stopped. Ischemia-reperfusion which recovered the supply of oxygen could induce oxygen free radicals burst increase at short time (Di Meo and Venditti, 2001) which resulted in highest level of MDA compared with red cell and quadriceps femoris. Compared with quadriceps femoris, mitochondrial with higher activity of antioxidant enzyme could not scavenge the free radical in time and produced more MDA. Antioxidant enzyme activity can be modified differently (Finaud et al., 2006) which was one of the reasons that enzyme activity varied in different researches (Singh et al., 2002; Ho et al., 2003; Wang et al., 2008).
High-intensity exercise, swimming, could induce the accumulation of glycolytic stimulant (such as ADP, AMP, Pi) which accelerate glycolytic rate and the production of pyruvate, resulting increased production of lactate. The content of lactate varied quickly according to the exercise intensity, rest time, determination methods and physical states of subject. To guarantee the consistence of the determination, the blood sample must be got at the same time. This could explain why several researches had obvious different results in levels of plasma biochemical parameters (Klein et al., 1996; Prasad and Khanum, 2012; Wang et al., 2008). Increased fatty acid utilization meant the enhanced NEFA concentration which was thought the greater fat metabolism and in turn decreased glucose depletion and led to increased exercise capacity (Klein et al., 1996). The reason might be that uptake whey hydrolysates increased swimming exercise capacity of mice which stimulated the utilization of lipid to spare the glucose. High level of glucose was consistent with low lactate and high NEFA for WHL, WHH, WPL, WPH and WTH groups compared with vehicle group. For WPH group, the difference was significant in the levels of glucose and NEFA which demonstrated that WPH group could accelerate effectively the utilization of lipid to increase the storage of glucose. Intense exercise and excessive protein intake could increase the content of blood BUN. High dose of WH and WP could increase the levels of BUN in WHH and WPH groups which was consistent with the result that more peptides uptake, more absorption of amino acid and more content of BUN but the differences between vehicle group and WHH or WPH group were not significant. Compared with peptides, protein was not suitable for the absorption which maybe the reason of low BUN content in MAL group.
Low levels of muscle glycogen and liver glycogen due to swimming exercise have been shown to cause fatigue and thus decrement in performance. Low muscle glycogen levels could also lead to oxidation of the branched chain amino acids and central fatigue (Snyder, 1998). Liver glycogen in body is limited and its depletion is not only the cause of fatigue but also the determinants of fatigue occurrence time. During the course of glycogen recovery process after exercise, muscle glycogen restored at first, then the liver glycogen which was affected by exercise time and intensity (Clark and Conlee, 1979). To avoid the error, the glycogen content must be determined at the same time. Although the levels of muscle glycogen in all groups were not significant different, WHH, WPL and WPH increased significantly the liver glycogen content, especially for WPH which was consistent with the longer swimming time. Glycogen was the important resource of energy during swimming exercise, the increasing of liver glycogen storage was advantage to improve the endurance of the exercise.
Addition of antioxidant lactic acid bacterial, Lactobacillus plantarum TUST200209, could increase the antioxidant activity (free radical-scavenging activity and metal ion-chelating capacity) of whey hydrolysates (data not shown) which was consistent with research of Amadou et al. (2009) on other lactic acid bacteria. Both intact cells and intracellular cell-free extracts of some lactic acid demonstrated antioxidant activity. But antioxidant ability and antifatigue capacity of protein hydrolysates with Lactobacillus plantarum TUST200209 were less than that of whey peptides. It might be superior free radical-scavenging activity and metal ion chelating capacity of whey peptides.
CONCLUSIONS
This research first reported that whey hydrolysates fermented by Lactobacillus plantarum TUST200209 could delay fatigue. Purification of WH improved the levels of SOD, GSH-Px and decreased MDA in red cell, mitochondria and quadriceps femoris which demonstrated good oxygen free radical-scavenging capacity. High content of glucose and NEFA, low levels of lactate and more storage of liver glycogen also indicated in WPH group. All the results demonstrated that there was positive relationship between antioxidant capacity and antifatigue. Therefore, WP could effectively retard fatigue induced by ROS and whey hydrolysates could used as promising ingredients in food industry.
REFERENCES
- Ali, R.F.M., 2011. Antioxidative effects of pomposia extract, on lipid oxidation and quality of ground beef during refrigerated storage. Am. J. Food Technol., 6: 52-62.
CrossRefDirect Link - Atamaniuk, J., C. Vidotto, H. Tschan, N. Bachl, K.M. Stuhlmeier and M.M. Muller, 2004. Increased concentrations of cell-free plasma DNA after exhaustive exercise. Clin. Chem., 50: 1668-1670.
PubMed - Bostrom, S., M. Fahlen, A. Hijalmarson and R. Johansson, 1974. Activities of rat muscle enzymes after acute exercise. Acta Physiol. Scand., 90: 544-554.
PubMed - Clark, J.H. and R.K. Conlee, 1979. Muscle and liver glycogen content: Diurnal variation and endurance. J. Applied Physiol., 47: 425-428.
Direct Link - Davies, K.J., A.T. Quintanilha, G.A. Brooks and L. Packer, 1982. Free radicals and tissue damage produced by exercise. Biochem. Biophys. Res. Commun., 107: 1198-1205.
PubMed - Di Meo, S. and P. Venditti, 2001. Mitochondria in exercise-induced oxidative stress. Biol. Signals Recept., 10: 125-140.
PubMed - Elia, D., K. Stadler, V. Horvath and J. Jakus, 2006. Effect of soy-and whey protein-isolate supplemented diet on the redox parameters of trained mice. Eur. J. Nutr., 45: 259-266.
CrossRefPubMedDirect Link - Finaud, J., G. Lac and E. Filaire, 2006. Oxidative stress relationship with exercise and training. Sports Med., 36: 327-358.
PubMed - Geethalakshmi, R., D.V.L. Sarada and P. Marimuthu, 2010. Evaluation of antimicrobial and antioxidant potentials of Trianthema decandra L. Asian J. Biotechnol., 2: 225-231.
CrossRefDirect Link - Halliwell, B. and S. Chirico, 1993. Lipid peroxidation: Its mechanism, measurement and significance. Am. J. Clin. Nutr., 57: 715S-724S.
Direct Link - Harman, D., 1956. Aging: A theory based on free radical and radiation chemistry. J. Gerontol., 11: 298-300.
Direct Link - Hernandez-Ledesma, B., A. Davalos, B. Bartolome and L. Amigo, 2005. Preparation of antioxidant enzymatic hydrolysates from α-lactalbumin and β-lactoglobulin. Identification of active peptides by HPLC-MS/MS. J. Agric. Food Chem., 53: 588-593.
CrossRefDirect Link - Ho, S.C., J.H. Liu and R.Y. Wu, 2003. Establishment of the mimetic aging effect in mice caused by D-galactose. Biogerontology, 4: 15-18.
CrossRefDirect Link - Ikeuchi, M., T. Koyama, J. Takahashi and K. Yazawa, 2006. Effects of astaxanthin supplementation on exercise-induced fatigue in mice. Biol. Pharm. Bull., 29: 2106-2110.
PubMed - Amadou, I., S. Jin, M.T. Kamara, Y.H. Shi, O.S. Gbadamosi and L. Guo-Wei, 2009. Characterization in vitro trypsin digestibility and antioxidant activity of fermented soybean protein meal with Lactobacillus plantarum Lp6. Am. J. Food Technol., 4: 268-276.
CrossRefDirect Link - Jung, K.A., D. Han, E.K. Kwon, C.H. Lee and Y.E. Kim, 2007. Antifatigue effect of Rubus coreanus miquel extract in mice. J. Med. Food, 10: 689-693.
PubMed - Kamakura, M., N. Mitani, T. Fukuda and M. Fukushima, 2001. Antifatigue effect of fresh royal jelly in mice. J. Nutr. Sci. Vitaminol., 47: 394-401.
Direct Link - Kamau, S.M. and R.R. Lu, 2011. The effect of enzymes and hydrolysis conditions on degree of hydrolysis and DPPH radical scavenging activity of whey protein hydrolysates. Curr. Res. Dairy Sci., 3: 25-35.
CrossRefDirect Link - Klein, S., J.M. Weber, E.F. Coyle and R.R. Wolfe, 1996. Effect of endurance training on glycerol kinetics during strenuous exercise in humans. Metabolism, 45: 357-361.
PubMed - Ji, L.L. and S. Leichtweis, 1997. Exercise and oxidative stress: Sources of free radicals and their impact on antioxidant systems. Age, 20: 91-106.
CrossRef - Misra, S.D., R. Maiti, C. Mallick and D. Ghosh, 2006. Protective response of methanolic extract of Ocimum sanctum, Withania somnifera and Zingiber officinalis on swimming-induced oxidative damage on cardiac, skeletal and brain tissues in male rat: A duration dependent study. Int. J. Pharmacol., 2: 647-655.
CrossRefDirect Link - Miki, M., H. Tamai, M. Mino, Y. Yamamoto and E. Niki, 1987. Free-radical chain oxidation of rat red blood cells by molecular oxygen and its inhibition by α-tocopherol. Arch. Biochem. Biophys., 258: 373-380.
PubMed - Powers, S.K., K.C. DeRuisseau, J. Quindry and K.L. Hamilton, 2004. Dietary antioxidants and exercise. J. Sports Sci., 22: 81-94.
PubMed - Ren, J., M. Zhao, H. Wang, C. Cui and L. You, 2011. Effects of supplementation with grass carp protein versus peptide on swimming endurance in mice. Nutrition, 27: 789-795.
CrossRef - Singh, A., P.S. Naidu, S. Gupta and S. K. Kulkarni, 2002. Effect of natural and synthetic antioxidants in a mouse model of chronic fatigue syndrome. J. Med. Food, 5: 211-220.
PubMed - Snyder, A.C., 1998. Overtraining and glycogen depletion hypothesis. Med. Sci. Sports Exercise, 30: 1146-1150.
PubMed - Takenaka, A., H. Annaka, Y. Kimura, H. Aoki and K. Igarashi, 2003. Reduction of paraquat-induced oxidative stress in rats by dietary soy peptide. Biosci. Biotechnol. Biochem., 67: 278-283.
PubMed - Tharakan, B., M. Dhanasekaran and B.V. Manyam, 2005. Antioxidant and DNA protecting properties of anti-fatigue herb trichopus zeylanicus. Phytother. Res., 19: 669-673.
CrossRefPubMedDirect Link - Tong, L.M., S. Sasaki, D.J. McClements and E.A. Decker, 2000. Antioxidant activity of whey in a salmon oil emulsion. J. Food Sci., 65: 1325-1329.
CrossRef - Wang, L., H.L. Zhang, R. Lu, Y.J. Zhou and R. Ma et al., 2008. The decapeptide CMS001 enhances swimming ndurance in mice. Peptides, 29: 1176-1182.
PubMed - Yu, B., Z.X. Lu, X.M. Bie, F.X. Lu and X.Q. Huang, 2008. Scavenging and anti-fatigue activity of fermented defatted soybean peptides. Eur. Food Res. Technol., 226: 415-421.
CrossRefDirect Link - Prasad, M.P.V. and F. Khanum, 2012. Antifatigue activity of ethanolic extract of Ocimum sanctum in rats. Res. J. Med. Plant, 6: 37-46.
CrossRef