
Wu-Yi Liu
Department of Animal Science, Chinese Agricultural University, Yuanmingyuan West Road No. 2, Beijing 100093, People�s Republic of China
Chun-Jiang Zhao
Department of Animal Science, Chinese Agricultural University, Yuanmingyuan West Road No. 2, Beijing 100093, People�s Republic of China
Asian Journal of Animal and Veterinary Advances
Year: 2010 | Volume: 5 | Issue: 6 | Page No.: 388-401







ABSTRACT
With the aim of testing whether there are possible genetic communications between Chinese gamecocks and their neighboring native chicken breeds, this study comprehensively analyzed an integrative mitochondrial DNA (mtDNA) data of 328 chicken (69 gamecocks belonging to 5 breeds and 259 domesticated chicken belonging to 18 breeds). Analysis of Molecular Variance (AMOVA) and phylogeny demonstrated Chinese gamecocks formed monophyletic groups that had been differentiated significantly with other native chicken breeds, though some of the clusters appeared to be an admixture of gamecocks and domesticated chicken. All the analysis demonstrated significant differentiation within chicken populations sampled and presented a gene flow mode of geographical distribution (p<0.05, AMOVA). In this study, we found most of the chicken breeds have been an admixture with possible gene flow among them, except for three gamecock breeds and three native chicken breeds, Chahua, Gushi and Tibetan chicken, whose races are relatively pure. Our study provided important clues for the research and understanding of the evolutionary and genetics relationship among Chinese native chicken breeds.
PDF Abstract XML References Citation
How to cite this article
Wu-Yi Liu and Chun-Jiang Zhao, 2010. Comprehensive Genetic Analysis with Mitochondrial DNA Data Reveals the Population Evolution Relationship Between Chinese Gamecocks and Their Neighboring Native Chicken Breeds. Asian Journal of Animal and Veterinary Advances, 5: 388-401.
DOI: 10.3923/ajava.2010.388.401
URL: https://scialert.net/abstract/?doi=ajava.2010.388.401
DOI: 10.3923/ajava.2010.388.401
URL: https://scialert.net/abstract/?doi=ajava.2010.388.401
INTRODUCTION
The domestication of chicken is often associated with the progression of human society and culture. Domestic chicken are now distributed all over the world and generally being classified into four categories, such as entertainment, ornamental, meat and egg production according to their utilities for human. Domestic chicken were believed to have been originated from the red jungle fowl (Gallus gallus) as early as 5400 B.C. (Zeuner, 1963; West and Zhou, 1989) with archaeological discoveries in the regions of Indus Valley and Hebei Province of China, respectively.
There are five possible progenitor subspecies of the red jungle fowl G.g. gallus (Gallus gallus gallus) in Thailand and its adjacent regions, G.g. spadiceus in Burma and Yunnan Province of China, G.g. jabouillei in Southern China and Vietnam, G.g. murghi in India and G.g. bankiva in the Java islands (Crawford, 1995; Howard, 1984) and presently researchers defend two hypotheses on the origination of domestic chicken: monophyletic origin of chicken from the G.g. gallus in Thailand and the Indus valley (Fumihito et al., 1994, 1996) and multiple origins of chicken from the G.g. gallus with independent domestication incidences in different regions (Crawford, 1990, 1995; Liu et al., 2006a). Liu et al. (2006a) made the first large scale analysis of the mitochondrial DNA hyper-variable segment I (HVS-I) from 834 domestic chickens (Gallus gallus domesticus) sampled across Eurasia with 66 wild red jungle fowls (Gallus gallus) from Southeast Asia and China. Their phylogenetic analysis revealed a dispersal pattern of nine highly divergent mtDNA clades (A-I) for both the red jungle fowls and domestic chickens, but there was no breed-specific clade in the study.
Although, the academic viewpoint of chicken multiple maternal origins seems ready to be acceptable, it is not yet known how many subspecies of the red jungle fowl have contributed to the origination of some particular chicken breeds, such as gamecocks. The authors also admitted that they failed to identify breed-specific matrilineal clades in their study (Liu et al., 2006a) and most of their samples from China were distributed in Yunnan Province and the adjacent provinces. Their speculation about the clade D which mainly contained red jungle fowl and gamecocks was still in need of verification. Until now, we have no direct or clear evidence about the gene flow between the wild red jungle fowl and gamecocks and it is natural to make a survey on the relationship between the Chinese gamecocks and other native chicken breeds in adjacent areas with integrative historical records and genetic data.
There are many historic records or biographies describing the ancient cockfighting in China (Xie, 1980, 1992; Zhu, 1999). It’s true that the Japanese envoi went to China and returned Japan with the gamecocks and spreading the cockfighting culture ever since in the Tang Dynasty, approximately 1300 years ago (Xie, 1980, 1992). It was believed that China was one of the earliest ancient nations raising cocks for cockfighting and the cockfighting culture originated in China and dispersed to East and Southeast Asia. Since, the introduction of gamecocks was believed to be associated with culture communication and the national historical stories, tracing the origin of gamecocks could thus provide key clues on the spreading of cockfighting games and its culture among different countries and/or regions.
According to cockfighting game players, ancient people used fighting cocks to resolve disputes at an earlier time. Later, people found it interesting to make cocks fight with each other and created the cockfighting games. Therefore, gamecock was bred from domestic chicken and isolated for special fighting trainings. There should not be any genetic communications between gamecocks and other chicken breeds. However, according to some historic documents or records, it was assumed that the famous Shouguan chicken and some other breeds were the decedents of Chinese Luxi gamecocks from ancient Qi kingdom at the Chun Qiu Period (770-476 B.C.) (Xie, 1980). The famous breed Shouguang chicken in Shandong Province of China had been formed and was once trained and exploited as cockfighting chicken during the Northern and Southern Dynasties Period of China (420-589 A.D.). It is also meaningful to trace the relationship between the Shouguang chicken and Luxi gamecock using mitochondrial DNA (mtDNA) data.
Recently, Komiyama et al. (2003) analyzed 47 gamecock mtDNA data (42 from Japan, 2 from Myanmar and 3 from China) and suggested a dual origin of Japanese gamecocks, with one arising from mainland China and the other from Southeast Asia (Komiyama et al., 2003). They further analyzed the relationship of Japanese domesticated chicken and gamecocks and suggested that most of the Japanese domesticated chicken have been derived from the ancestors of Japanese gamecocks (Komiyama et al., 2004). Liu et al. (2006b) also analyzed the 42 Japanese gamecock mtDNA data, adding 52 Chinese gamecock mtDNA sequences and advocated a single origin hypothesis of the Japanese gamecock from China (either directly from China or indirectly via Southeast Asia). Other than this, they had no more discussion on the population differentiation or gene flow among chicken populations in relation to Chinese gamecocks. Qu et al. (2009) analyzed the genetic relationships among Chinese gamecock breeds with mtDNA data and proposed that gamecock breeds might originated from domestic chicken or wild birds directly, but their suggestion was not convincing since no evidence from red jungle fowls and other domestic chicken was demonstrated. Moreover, these authors had not analyzed the relationship of Chinese chicken breeds and gamecocks, or testing the hypothesis that there are possible genetic communications or hybridizations between Chinese gamecocks and their neighboring chicken breeds, as Komiyama et al. (2004) did in Japanese indigenous chicken.
In this study, we compared the first hyper-variable segment I sequences of the mtDNA control region (HVS-I) of 69 gamecocks belonging to 5 breeds across China and 259 native chicken belonging to 18 breeds sampled from the neighboring regions or provinces, with sequences data retrieved from GenBank and previous works (Niu et al., 2002; Komiyama et al., 2003; Liu et al., 2004; Qu, 2004; Liu et al., 2006b; Song, 2006; Bao, 2007). First, we constructed phylogenetic trees and compared the mtDNA data of Chinese gamecocks with the neighboring native breeds using the red jungle fowls as the outgroup, in order to trace and learn more about the genetic information between Chinese gamecocks and the neighboring native chicken breeds and the followed artificial selection. Then, we analyzed the gene flow at an inter-group level and computed the corresponding population differentiation, particularly focusing on Chinese gamecocks. We found most of the chicken breeds have been an admixture with possible much gene flow among them, except for three gamecock breeds and three native chicken breeds. We also found there was no obvious genetic evidence for the assumption that the Shouguan chicken breed was descended form Chinese Luxi gamecocks.
MATERIAL AND METHODS
Data
The sample data were collected and the research project was conducted from October, 2007 to November, 2009, with two necessary updates in mid 2009. We retrieved 69 Chinese gamecock mtDNA data, in which 62 were from GenBank (accession No. AY588608-42, AB098664-6, AY465968-71, AF512108-16, DQ462521-5, DQ462547-9, DQ462551-2) and 8 were provided by Bao (2007), (Henan gamecock, recoded as HnDJ013-20). Then, we got a total of 258 Chinese native chicken mtDNAs belonging to 18 breeds in the neighboring regions, some from GenBank (accession nos. AY465960-6, AY465972-9, AY465981-007, AF512076-90, DQ462526-46, DQ462550, DQ462553-6, DQ462563-70, AF128315-24, AF128330-4, AF128340-4), the rest were provided by Bao (2007) and Qu (2004). The reported 15 mtDNAs of red jungle fowls (8 from China and 4 from Thailand, Bao 2007; another 3 strains are from Indonesian, accession No. AB007718, AB007720 and AB009431; Komiyama et al., 2003) were also compiled and put into analysis. Detailed sample information is presented in Table 1.
Among all the reported sequences, two (Accession No. AY588607 and AY465967) were deleted and not included in the following analysis as suspected of sequencing errors (Liu et al., 2006a). It should be noted that, in the study of Liu et al. (2006a), their samples from China were mainly from Yunnan Province and adjacent provinces (415 out of 595, or about 74.21%) and the clade D were mainly composed of red jungle fowl and gamecocks (Liu et al., 2006a).
| Table 1: | Genetic diversity of Chinese native chicken samples |
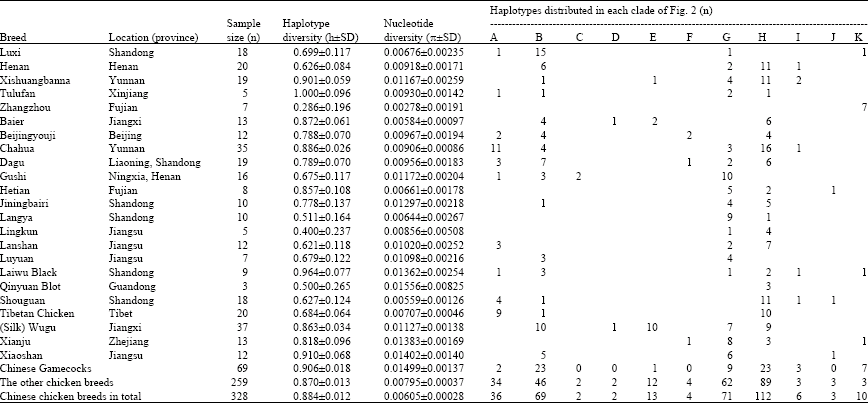 | |
| The estimation is restricted to the 516 bp fragment relative to sequence position 1-516 of mtDNA HVS-I region | |
Phylogenetic Analysis
Sequences (gamecocks and the other native chicken) were aligned and the nucleotide polymorphisms were reported using MEGA version 4.0 (Tamura et al., 2007). We discarded any insertions/deletion (e.g., indel) in the following analysis. First, we constructed rooted NJ (neighbor-joining) trees for Chinese gamecocks with Indonesian red jungle fowls as outgroup and unrooted NJ trees for the haplotypes shared by Chinese gamecocks and their neighboring native chicken breeds considered in this study by means of Kimura two-parameter model. We validated phylogenetic trees with interior branch bootstrap test of 1,000 replicates to create second trees respectively. We focused interests on the main clades emerging from the trees.
Meanwhile, the genetic relationships of tree clades were further explored by median-joining method (Bandelt et al., 1999). The graphic median-joining (MJ) networks were drawn according to Bandelt et al. (1999, 2000) and confirmed using Network (http://www.fluxus-engineering.com/sharenet.htm). To learn more about the genetic diversity of the gamecock, we estimated haplotype diversity (h) and nucleotide diversity (ð) for samples from each breed and two serials of samples from assembled breeds using DnaSP version 4.0 (Rozas et al., 2003).
Population Differentiation and Gene Flow
Population comparison and population differentiation were executed in an integrated group or a few groups in different geographic regions with gamecocks and the neighboring native chicken breeds as populations or subgroups, computing gene flow and genetic differentiation with haplotype genotypic data in the software package ARLEQUIN version 3.11 (Excoffier and Schneider, 2005). Inter-group level analysis, such as analysis of molecular variance (AMOVA), estimates of pairwise genetic fixation indices (Fst) and gene flow indices (Nm), were calculated for evaluating the evolutionary relationship among different Chinese chicken breeds.
First, we used the structure package version 2.3.1 (Pritchard et al., 2000; Falush et al., 2003) to infer the proper number of populations and assumed a mode with population admixture and that the allele frequencies were correlated within populations (Falush et al., 2003). We conducted 10 independent runs for each value of K (the number of subpopulations) between 1 and 23. After conducting numerous runs to investigate the behaviour of the programme, we chose to use a burn-in period of 104 iterations and then collect data for 105 iterations. We ran the software also with the same parameters separately for both Gamecocks and their neighboring chicken samples. The presence of genetic structure among the Chinese native chicken populations was investigated also by an analysis of variance framework using analysis of molecular variance (AMOVA). We next used the ARLEQUIN package version 3.11 (Excoffier and Schneider, 2005) to perform the AMOVA analysis.
We used both softwares ARLEQUIN and DnaSP to estimate each pairwise genetic distances (Fst) and the corresponding estimate of the average effective number of migrants (Nm) exchanged among the Chinese 5 gamecock breeds and 23 native chicken populations. Exact tests of population differentiation between the chicken populations were conducted as described by Raymond and Rousset (1995) using ARLEQUIN with 10000 markov chain interactions.
It should be noticed that the estimates of Nm are based on the island mode of population structure (Wright, 1951; Nei, 1987), where N is the number of individuals in each subpopulation sampled and m is the fraction of migrants in each subpopulation in each generation. For instance, Fst = 1/(1 + 2Nm) on haploids (mitochondrial and chloroplast genomes) and Fst = 1/(1 + 4Nm) on diploids (autosome), or Fst = 1/(1 + 3Nm) on diploids (X-chromosome), or Fst = 1/(1 + Nm) on diploids (Y-chromosome). If Fst is less than or equal to 0, the estimator is considered undefined and not analyzed later.
RESULTS
Sequence Variation and Genetic Diversity
The mtDNA sequence variations of 69 Chinese gamecocks and 259 the other Chinese native chickens are shown Table 1. An integrated population group of 328 chicken belonging to 5 gamecock breeds and 18 native chicken breeds was further analyzed with phylogenetic tree and supported by graphic profiles of MJ network, although some distances between nodes were only one or two mutation steps.
In total, we had identified 59 variable sites which defined 60 haplotypes in the integrated large population group (detailed information could be showed if demanded), where single variable sites were identified in 32 loci. Statistics of all the haplotypes and nucleotide diversities were summarized in Table 1. Among five Chinese gamecock breeds, Tulufan and Banna (Xishuangbanna) samples have high haplotype diversities (h = 1.000±0.096 and h = 0.901±0.059, respectively), Banna gamecock samples have a high nucleotide diversity (π = 0.01167±0.00259), whereas Zhangzhou samples show the lowest haplotype and nucleotide diversities (h = 0.286±0.196, π = 0.00278±0.00191). The average genetic diversity of Chinese gamecock samples (h = 0.906±0.018, π = 0.01499±0.00137) is relatively higher than that of the other Chinese native chicken samples (h = 0.870±0.013, π = 0.00795±0.00037).
Phylogenetic Tree and MJ Network Profile Show Geographical Distributions
Phylogenetic trees were built with mtDNA haplotypes of all the Chinese native chicken samples and red jungle fowls. Clear clades could be figured out. Eleven main clades or nodes (A-K) are discerned in both the unrooted NJ tree and MJ network built with mtDNA haplotypes of Chinese gamecocks and the neighboring chicken breeds (Fig. 1, 2). The NJ tree was bootstrapped with 1000 replicates and the haplotype clades were further supported by the MJ network profile (Fig. 2), which has been widely used in reconstructing maximum parsimony phylogenies for intra-specific mtDNA data (Bandelt et al., 1999).
The first NJ phylogenetic tree was constructed after analyzing an integrative set of 328 sequences from Chinese gamecocks and the neighboring native chicken breeds (Fig. 1). This NJ phylogeny tree recovered 60 haplotypes for all the Chinese native chicken within 11 clades partitioned according to the cluster shapes. In general, the alleged Central Chinese gamecocks (including Luxi and Henan gamecock breeds) differ from other Chinese chicken breeds and cluster together by harboring a high amount of lineages in clades B, G and H, whereas the majority of lineages in Chinese gamecocks present (Fig. 1, Table 1). The Henan and Banna gamecocks got an admixture cluster in both two sets of trees, while the majority of Luxi gamecocks did not join in other chicken lineages except Henan gamecocks (Fig. 2, Table 1). It shows the Central Chinese gamecock lineages are relatively conserved. The NJ tree was supported by MJ network profiles very well (Fig. 2). It was also seen that Zhangzhou gamecocks were isolated from the other gamecocks (Fig. 2). Moreover, clades or nodes in Fig. 1 and 2 are in accord with the geographical distribution of the Chinese native chicken lineages. For instance, Luxi gamecocks from Shandong Province are clustered more close to chicken breeds from Shandong Province and the neighbor Jiangsu and Henan Provinces. Tibetan chicken comprise lineages close to Chahua chicken sampled from the neighboring Yunan Province.
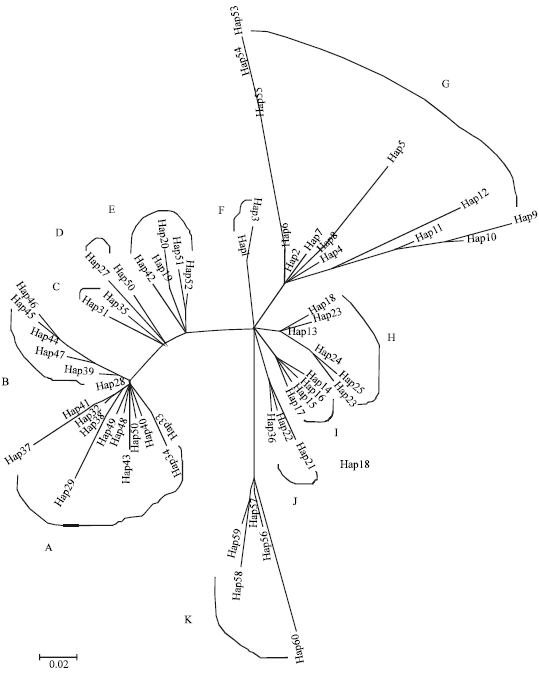 | |
| Fig. 1: | NJ tree of 60 mtDNA haplotypes for Chinese gamecocks and their neighboring native chicken. Eleven clades of mtDNA haplotypes (coded as hap) were found in the phylogenetic tree |
Genetic Structure and (AMOVA) Analysis of Chinese Chicken Population
Phylogenetic analysis can provided the clustering of clades with sequence similarity, but it brings no more population information to us, such as the population structure and differentiation. So, we analyzed the genetic structure among Chinese chicken populations, including gamecock breeds.
In the Bayesian analysis of population structure (significance tests at 1023 permutations) (Pritchard et al., 2000; Falush et al., 2003), the probabilities of six clusters (K = 1-6) were relatively high and the variants of LnP(D) with K cluster values belonging to different samples were the lowest ones (Fig. 3).
 | |
| Fig. 2: | MJ network profile of 60 mtDNA haplotypes of the Chinese gamecock and the neighboring native chicken. The codes of LxDJ, HnDJ, BnDJ, TlfDJ and ZzhDJ stand for five gamecock breeds in China, while the codes of RDch, RDth and RDini mean red jungle fowls from China, Thailand and Indonesian, respectively. The codes of Ber, BJY, ChH, DaG, GuS, HeT, JnBr, LanY, LinK, LSH, LuY, LwuB, QYB, ShG, Tib, WuG, XJu, XSH are the other 18 Chinese native chicken breeds, i.e., Baier, Beijingyouji, Chahua, Dagu, Gushi, Hetian, Jiningbairi, Langya, Lingkun, Lanshan, Luyuan, Laiwu Black, Qinyuan Blot, Shouguan, Tibetan Chicken, Wugu (Silk), Xianju, Xiaoshan stand for five gamecock breeds, respectively |
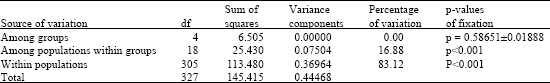
According to the plot Figure (Fig. 3), the most probable number and sub-structuring mode of population groups was five. AMOVA analysis with five geographical distributing sub-groups (we selected five vicinal areas, e.g., Shandong and neighboring regions, Fujian and neighboring regions, Yunnan, Xinjiang and Tibietan) suggested no significant differentiation among the five population groups of Chinese gamecock and their neighbor native chicken (p = 0.58651±0.01888), but significant differentiations within populations (p<0.001) and among populations within groups (p<0.001) (Table 2). Most of the variation (83.12%) appeared to be within populations and the rest appeared to be among populations within groups (16.88%) (Table 2). No variation was found to be within groups since the F statistics of variation base on haplotype frequencies turned out to be negative and was denied. According to the plot of structure mode choice criterion LnP(D) (Fig. 3), the STRUCTURE analysis with both Chinese gamecocks and the native chicken might suggest some other possible substructures grouping in all the chicken populations with lower LnP(D) values too.
We also did the AMOVA analysis of two groups of Chinese gamecocks and the other native chicken, assuming Chinese gamecocks were isolated from the other populations sampled. The overall genetic differences of Chinese gamecocks and their neighbor chicken populations suggested a near significant differentiation among these two population groups (p = 0.05963±0.00883, F-statistics of variation based on haplotype frequencies and 1023 permutations, Table 2).
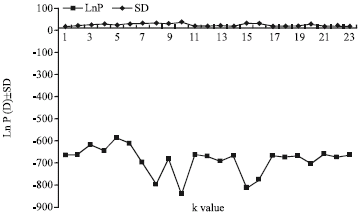 | |
| Fig. 3: | Mode choice criterion Ln P(D) of the structure for each k cluster value. The k cluster value is an index for mode selection in STRUCTURE. The structure algorithm starts at a random place in parameter space and then converges towards a mode of the parameter space. In this context, a mode can be thought of as a clustering solution that has high posterior probability |
| Table 2: | AMOVA Analysis of five groups computed based on conventional F-Statistics of haplotype frequencies (significance tests at 1023 permutations) |
 | |
| Table 3: | AMOVA Analysis of two groups computed based on conventional F-Statistics of haplotype frequencies (significance tests at 1023 permutations) |
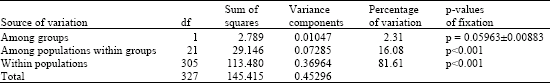 | |
Most of the variation (81.61%) appeared to be within populations and the rest appeared to be within groups (2.31%) and among populations within groups (16.08%) (Table 3). However, the variation between population groups was near significant (p = 0.05963±0.00883) and explained 2.31% of the total variation and the Fst estimate of 0.26258 would suggest Nm = 1.40418 as the average effective number of migrants exchanged per generation between the population groups.
The pairwise estimates of Fst among chicken breeds or populations suggested that the chicken populations are genetically close to each other (Fst and Nm, Appendix 1). Some Fsts are less than 0, these estimators are undefined and not considered. The pairwise estimates and non-differentiation exact test (see the p-values matrix, significance level at 0.05, markov chain length 10000 steps) provided us useful clues to infer the population differentiation and gene flow.
Simulation Analysis of Gene Flow and Population Differentiation Test
The pairwise Fst matrixes estimated by ARLEQUIN and DnaSP are similar (Weir and Cockerham, 1984; Wright, 1951), but Fst values provided by ARLEQUIN are much lower than those by DnaSP and probably would lead a set of higher Nm estimates. Here, we consecutively took the pairwise Fst and Nm estimates matrix computed by ARLEQUIN to infer whether there is any possible genetic spreading or gene flow between Chinese gamecocks with the other Chinese native chicken breeds and made the non-differentiation exact test (p<0.05, p-values can be found in Appendix 2).
Finally, we tested the hypothesis of random distribution of the individuals between pairs of populations in the simulation as described in Raymond and Rousset (1995) and Goudet et al. (1996). This test is analogous to Fisher’s exact test on a two-by-two contingency table, but extended to a contingency table of size two (Raymond and Rousset, 1995). All pairs of samples were taken into a non-differentiation test with 10000 markov chain steps done and the exact simulated p-value matrix of pairwise Fst estimates was produced with 6000 markov chain steps. The p-values with sign + showed differentiation and p-values with sign - showed non-differentiation at the significance level of 0.0500.
DISCUSSION
Liu et al. (2004) and Song (2006) analyzed the genetic diversities with mtDNA data of a number of Chinese native chicken breeds, including 4 Henan and 9 Luxi gamecocks, but their sample sizes were small and could not resolved the issue whether there are possible genetic communications between Chinese gamecocks and their neighboring chicken breeds. Liu et al. (2006a) made a large scale phylogenetic analysis of the mtDNA data from 834 domestic chickens, but there were no breed-specific clades. The authors acknowledged that they failed to identify breed-specific matrilineal clades in their study (Liu et al., 2006a). Their speculation about the clade D which mainly contained red jungle fowl and gamecocks was still in need of verification. Later, Liu et al. (2006b) analyzed 42 Japanese gamecock mtDNA data and 52 Chinese gamecock mtDNA sequences and advocated a single origin hypothesis of the Japanese gamecock from China. Qu et al. (2009) proposed that gamecock breeds might originated from domestic chicken or wild birds directly, but their suggestion was not convincing since no evidence from the other domestic chicken was demonstrated. However, these authors had no more discussion on the population structure or gene flow within Chinese gamecocks. They also had not fully analyzed the relationship of Chinese gamecocks and other native chicken breeds, as Komiyama et al. (2004) did in Japanese indigenous chicken. In the present study, we comprehensively tested the hypothesis whether there are possible genetic communications between Chinese gamecocks and their neighboring native chicken breeds and found a worrying situation for Chinese gamecocks. Moreover, we found there was no obvious genetic evidence to verify the relationship that the Shouguan chicken breed was probability descended form Luxi gamecocks.
In the present analysis, we focused on the impact of population differentiation and admixture of Chinese gamecocks with other chicken breeds, which could be inferred from the AMOVA and phylogeny and population simulation analysis with significance level of the p-values. Analysis of the matrix data in Appendix 1 and 2 provided a perspective on the pattern of gene flow among Chinese native chicken breeds. Besides the Central Chinese gamecocks, there are some chicken breeds also show independent differentiation, such as Chahua and Gushi chicken. Apart from the Chinese gamecocks, most of the p-values of non-differentiation in the neighboring native chicken demonstrated a mode of geographical distribution. In the five regional group AMOVA analysis, observed haplotype frequencies are well according with expected haplotype frequencies under the infinite-allele model (data not showed). At present, we can not confirm the Shouguan chicken were the decedents of Chinese Luxi gamecocks. As expected, Luxi gamecocks are significantly differentiated from the other chicken breeds and the hypothesis that Shouguan chicken are the descendent of ancient Luxi gamecocks thus found no evidence in our analysis. The Central Chinese gamecock were independently raised and mated without admixture, which was supported by both phylogenetic analysis and pairwise population differentiation tests.
Another obviously independently differentiated breed is Zhangzhou gamecocks whose exact p-values appear to be significantly differentiated from other chicken breeds. Different from Bannan and Tulufang gamecocks, the Zhangzhou gamecocks were original from Southeast Asian by trade ships. The Zhangzhou trade ships and ports, thriving in the Ming Dynasty, linked the region to many areas through trade. These trade links allowed for gamecock transport and admixture with many Southeast Asian gamecock breeds (Wu, 1983), contributing to the unique genetic background of Zhangzhou gamecocks. Bannan gamecocks were reported to be frequently mixed other fighting cocks or roosters, such as Thailand and Myanmar (Burma) gamecocks. Tulufang gamecocks were thought to be the intercrossing descendent of Central Chinese gamecocks and Russian chicken. Origins of these three gamecocks are rather complex. Now, Tulufang gamecocks are becoming extinct. These analysis also suggest these two gamecock breeds are badly in need of conservation, though the Central Chinese gamecocks are relatively isolated and prosperous populations.
In China, gamecock (called Douji) is reared solely for use in cockfighting and as pets. The tradition of cockfighting can be traced to as early as 2500 years ago (Xie, 1980, 1992; West and Zhou, 1989; Zhu, 1999). Gamecock and cockfighting pictures found on some archaeological assemblages can date to 45-23 B.C. (Xie, 1980, 1992; Zhu, 1999), e.g., the stone picture of a woman feeding two gamecock-like chicken found in the Dazu district of Chinese Sichuan Province and the unearthed copper and crockery vases of approximately 2000 years ago displaying cockfighting scenes (Zhu, 1999). On the top of a vase copper excavated in the grave of Jiangsu Province of China, a small sculpture as gamecock-like creature was discerned (Xie, 1980, 1992; Zhu, 1999). Popularity of cockfighting in history undoubtedly contributed to disperse these ancient chickens geographically. The number of gamecocks has increased during the past 20 years in China since the cockfighting game or culture returns to be popular accompanying the Chinese increasing economic circumstances. At present, there are five long-time formed breeds, i.e., Luxi gamecock, Henan gamecock, Banna gamecock, Tolufang gamecock and Zhangzhou gamecock. The former two breeds, whose breed numbers are presently estimated each at near 100000, made up the so-called Central Chinese gamecocks, while those of the latter three breeds are estimated at about 10000, 3000 and 20000, respectively.
However, the Chinese gamecock breeds are badly in need of conservation now. The fairly high artificial selection and reproductive isolation of Chinese gamecock populations suggested that each population should be restricted to a genetic pool and vulnerable to harmful effects of inbreeding. Tulufang gamecocks have been threatened with extinction and had a relatively reduced scale due to geographical isolation and inbreeding, while Banna gamecocks were considered as heterozygous and attribute to much mixture with foreign fighting cocks. Present analysis and collected data of long term gene flow showed that many of the Chinese native chicken colonies could no longer be viewed as separate entities, though five geographical distributions could be discerned. On the contrary, many chicken breeds are considered to be declined because of extensive introduction and admixture. Recently, the popularity of cockfighting was increasing in China and many breeders have introgressed gamecock breeds from other provinces and even nations in order to improve their gamecocks’ fighting traits. This recent admixture could account for the lack of monophyletic haplotype groups and a decline trend, as well as high genetic diversities.
ACKNOWLEDGMENTS
This study was supported by grants from the National Basic Research Program of China (2006CB102101).
APPENDIX
| Appendix 1: | Table of Fsts (below diagonal) and Nms (above diagonal) |
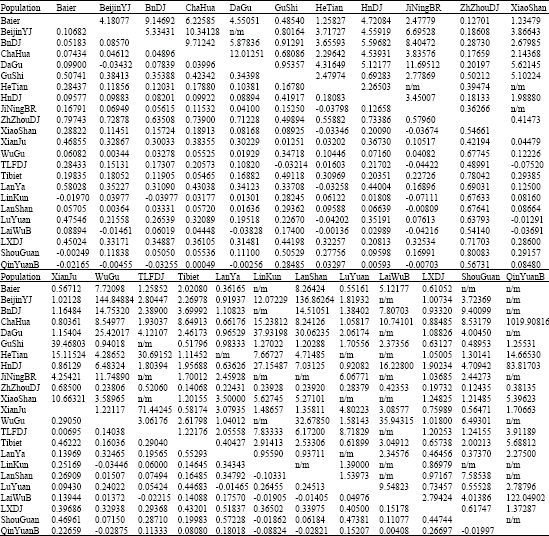 | |
| If Fst was less than or equal to 0, the estimator was considered undefined and not studied in the Nm analysis (noted as n/m) | |
| Appendix 2: | Table of markov simulated p-values (below diagonal) and significant differences (above diagonal) of non-differentiation exact tests |
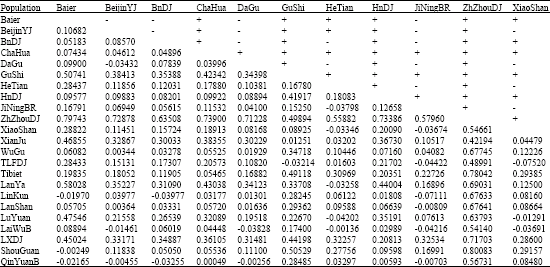 | |
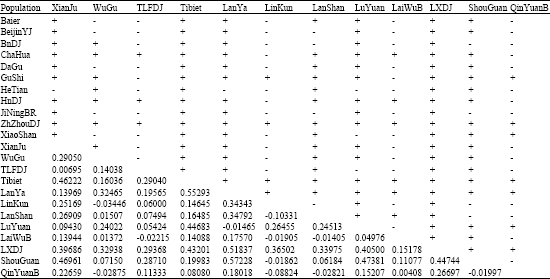 | |
| All pairs of samples were taken into a non-differentiation test with 10000 markov chain steps done and the exact p-value matrix of pairwise Fst estimates was produced with 6000 markov chain steps. The p-values with sign + showed differentiation and p-values with sign - showed non-differentiation (p<0.0500) | |
REFERENCES
- Bandelt, H.J., P. Forster and A. Rohl, 1999. Median-joining networks for inferring intraspecific phylogenies. Mol. Biol. Evol., 16: 37-48.
PubMedDirect Link - Excoffier, L.L.G. and S. Schneider, 2005. Arlequin ver. 3.0: An integrated software package for population genetics data analysis. Evol. Bioinform. Online, 1: 47-50.
Direct Link - Falush, D., M. Stephens and J.K. Pritchard, 2003. Inference of population structure using multilocus genotype data: Linked loci and correlated allele frequencies. Genetics, 164: 1567-1587.
PubMedDirect Link - Fumihito, A., T. Miyake, S. Sumi, M. Takada, S. Ohno and N. Kondo, 1994. One subspecies of the red junglefowl (Gallus gallus gallus) suffices as the matriarchic ancestor of all domestic breeds. Proc. Natl. Acad. Sci. USA., 91: 12505-12509.
PubMedDirect Link - Fumihito, A., T. Miyake, M. Takada, R. Shingu and T. Endo et al., 1996. Monophyletic origin and unique dispersal patterns of domestic fowls. Proc. Natl. Acad. Sci. USA., 93: 6792-6795.
PubMedDirect Link - Goudet, J., M. Raymond, T. de Meeus and F. Rousset, 1996. Testing differentiation in diploid populations. Genetics, 144: 1933-1940.
PubMedDirect Link - Howard, R., 1984. A Compete Checklist of Birds of the World. Oxford University, New York, pp: 701.
Direct Link - Komiyama, T., K. Ikeo and T. Gojobori, 2003. Where is the origin of the Japanese gamecocks? Gene, 317: 195-202.
CrossRefPubMedDirect Link - Liu, Y.P., G.S. Wu, Y.G. Yao, Y.W. Miao and G. Luikart et al., 2006. Multiple maternal origins of chickens: Out of the Asian jungles. Mol. Phylogenet. Evol., 38: 12-19.
CrossRefPubMedDirect Link - Liu, Y.P., Q. Zhu and Y.G. Yao, 2006. Genetic relationship of Chinese and Japanese gamecocks revealed by mtDNA sequence variation. Biochem. Genet., 44: 19-29.
PubMed - Liu, Z.G., C.Z. Lei, J. Luo, C. Ding and G. H. Chen et al., 2004. Genetic variability of mtDNA sequences in Chinese native chicken breeds. Asian-Aust. J. Anim. Sci., 7: 903-909.
Direct Link - Niu, D., Y. Fu, J. Luo, H. Ruan, X.P. Yu, G. Chen and Y.P. Zhang, 2002. The origin and genetic diversity of chinese native chicken breeds. Biochem. Genet., 40: 163-174.
CrossRefPubMedDirect Link - Pritchard, J.K., M. Stephens and P. Donnelly, 2000. Inference of population structure using multilocus genotype data. Genetics, 155: 945-959.
PubMedDirect Link - Raymond, M. and F. Rousset, 1995. An exact test for population differentiation. Evolution, 49: 1280-1283.
Direct Link - Rozas, J., J.C. Sanchez-DelBarrio, X. Messeguer and R. Rozas, 2003. DnaSP, DNA polymorphism analyses by the coalescent and other methods. Bioinformatics, 19: 2496-2497.
CrossRefDirect Link - Tamura, K., J. Dudley, M. Nei and S. Kumar, 2007. MEGA4: Molecular Evolutionary Genetics Analysis (MEGA) software version 4.0. Mol. Biol. Evol., 24: 1596-1599.
CrossRefPubMedDirect Link - Weir, B.S. and C.C. Cockerham, 1984. Estimating F-statistics for the analysis of population structure. Evolution, 38: 1358-1370.
CrossRefDirect Link - West, B. and B.X. Zhou, 1989. Did chickens go North New evidence for domestication. Worlds Poult. Sci. J., 45: 205-218.
CrossRefDirect Link - Wright, S., 1949. The genetical structure of populations. Ann. Eugenics, 15: 323-354.
CrossRefPubMedDirect Link - Qu, L.J., X.Y. Li and N. Yang, 2009. Genetic relationships among different breeds of chinese gamecocks revealed by mtDNA variation. Asian-Aust. J. Anim. Sci., 8: 1085-1090.
Direct Link
Valter Reply
It's a relief to find someone who can explain tihgns so well