
M.M. Khatun
Korean Zoonoses Research Institute, Chonbuk National University, Jeonju 561-756, Republic of Korea
M.A. Islam
Korean Zoonoses Research Institute, Chonbuk National University, Jeonju 561-756, Republic of Korea
B.K. Baek
Korean Zoonoses Research Institute, Chonbuk National University, Jeonju 561-756, Republic of Korea
S.I. Lee
Division of Model Animal, Institute of Biomedical Science, Kansai Medical University, Fumizono 10-15, Moriguchi, Osaka, 570-8506, Japan
Asian Journal of Animal and Veterinary Advances
Year: 2009 | Volume: 4 | Issue: 6 | Page No.: 267-277







ABSTRACT
The study was undertaken to investigate the cellular and humoral immune responses as well as antigen recognition in the acute and sub-acute stages of Brucella abortus biotype 1 infection in Sprague-Dawley (SD) rats. The SD rats were infected intraperitoneally with 1x1010 colony forming unit (cfu) of B. abortus biotype 1 Korean bovine isolate. The cellular and humoral immune responses were measured at 0, 3, 7, 14, 21, 28, 35, 42, 60, 90 and 120 days after infection against Crude Brucella Protein (CBP) by Lymphocyte Proliferation Assay (LPA) and Indirect Enzyme-linked Immunosorbent Assay (IELISA). The experimentally infected rats developed specific lymphoproliferative and humoral immune response within 1 week post infection. A significant increase in the proliferative response to CBP was recorded on day 28 post infection. Brucella abortus specific IgG responses were initiated in SD rats at 3 days after infection. The highest IgG antibody titers were recorded at 35 days after infection and then the titer gradually decreased until the end of the experiment. Recognition of immunodominant antigens in CBP of B. abortus was performed by Western Blot (WB) assay using infected rat sera collected at 0, 3, 7, 14, 21, 28, 35, 42, 60, 90 and 120 days after infection. Western blot assay of the sera using CBP antigens revealed a wide array of protein bands between molecular weight of 19 and 125 kDa. Proteins of 125, 105, 82, 66, 54, 46, 32, 24, 22, 21 and 19 kDa were frequently recognized by the sera of infected rats during the experiment. The 82, 46, 32, 24, 22, 21 and 19 kDa proteins were intensely recognized during the course of infection. These antigens should be considered useful for the diagnostic of B. abortus infection.
PDF Abstract XML References Citation
How to cite this article
M.M. Khatun, M.A. Islam, B.K. Baek and S.I. Lee, 2009. Cellular and Humoral Immune Responses and Antigen Recognition in Sprague-Dawley Rats Experimentally Infected with Brucella abortus Biotype 1. Asian Journal of Animal and Veterinary Advances, 4: 267-277.
DOI: 10.3923/ajava.2009.267.277
URL: https://scialert.net/abstract/?doi=ajava.2009.267.277
DOI: 10.3923/ajava.2009.267.277
URL: https://scialert.net/abstract/?doi=ajava.2009.267.277
INTRODUCTION
Brucella are Gram-negative, facultative intracellular bacteria which cause disease in humans, domesticated animals and wild mammals (Nicoletti, 1980; Young, 1983). Undulant fever, chills, sweating, anorexia, fatigue, weight loss, depression, arthralgia and myalgia are common clinical manifestations in human brucellosis patients. It causes abortion, infertility, stillbirth and retention of placenta in animals leading to huge economic losses (Radostits et al., 2007). Humans are generally infected through direct contact with infected animals or by the consumption of the contaminated food especially unpasteurized milk and milk products (Nicoletti, 1992; Pappas et al., 2006). Brucellosis mainly spread from one animal to another through contaminated materials during abortion, using infected bull or semen during natural breeding or artificial insemination (Lim et al., 2005).
Brucellosis remains endemic in many developing countries where it undermines animal health and productivity (Trujillo et al., 1994). At present, brucellosis is an emerging public and animal health problem in many countries as well as Korea despite animal brucellosis control program. The other most likely source of introduction of brucellosis in domesticated animals is from free ranging wildlife (Davis and Elzer, 2002). Rats are known to harbor Brucella in many parts of the world (Oliakova and Antoniuk, 1989) and found to be infected with B. abortus on farm where cattle are infected (Moore and Schnurrenberger, 1981). The control of brucellosis in animals as well as humans could not be achieved without eradicating the disease in primary reservoir hosts as well as free ranging wildlife (Davis and Elzer, 2002; Godfroid, 2002).
There are three clinical stages of brucellosis in humans according to the duration of the disease: acute, sub-acute and chronic. Acute brucellosis persists less than 8 weeks, duration of sub-acute stage is from 8 to 52 weeks and chronic brucellosis is more than 52 weeks (Gotuzzo and Cellillo, 1998). An acute phase of brucellosis can progress either to recovery or to a chronic form. The diagnosis of brucellosis in domesticated animals is based on the detection of antibodies against O polysaccharide of smooth Brucella sp., in the sera by the routine serological tests. Antibodies against the O polysaccharide of smooth Brucella are known to react with the closely related bacteria which hamper accurate diagnosis of Brucella (Kittelberger et al., 1998).
Antibody responses directed against proteins of Brucella is known to specific for the genus Brucella (Diaz and Moriyon, 1989). Test detecting antibodies to the Brucella proteins are considered specific (Cherwonogrodzky et al., 1990). Brucella proteins elicit cell-and antibody-mediated immune responses which have been investigated in animals and humans for diagnostic purpose. However, there is no study concerning diagnosis of brucellosis in free ranging wildlife such as rats using proteins of B. abortus. The aim of this study was to investigate the cellular and humoral immune responses against Crude Brucella Protein (CBP), during the acute and sub-acute stage of brucellosis in Sprague-Dawley (SD) rats and to investigate the specific antigen recognitions.
MATERIALS AND METHODS
Rats
Adult SD rats (n = 44) of 8 weeks old, were purchased from a commercial laboratory animal company (Koatech, Korea). The rats were housed in cages and provided with food and water ad libitum. The experimental protocol was approved by the local ethic committee of the Chonbuk National University, Korea. The animals were culture as well as seronegative for Brucella infection, prior to experimental infection, as ascertained by routine bacteriological examination of blood samples on brucella agar media (BBL, Becton, Dickinson and Company, Sparks, MD, USA) incubated at 37°C for 7 days under 5% CO2 and screening of sera by the Rose Bengal plate agglutination test (RBPT).
Bacterial Strain
Brucella abortus biotype 1 Korean cattle isolate was used for the experimental infection. Brucella abortus biotype 1 lyophilized stock culture was obtained from the laboratory repository. Stock culture was revived on brucella agar media (BBL, Becton, Dickinson and Company, Sparks, MD, USA) by incubating at 37°C for 7 days under 5% CO2. The grown bacteria were harvested in normal saline.
Experimental Inoculation
Forty rats were injected intraperitoneally with 0.1 mL sterile pyrogen free solution containing 1x1010 cfu mL-1 of B. abortus biotype 1. Four uninfected control rats were injected intraperitoneally with 0.1 mL sterile pyrogen free solution.
Clinical Examinations
Rectal temperature, food and water intake as well as other abnormal clinical signs of all infected and control rats were recorded daily for 2 weeks.
Preparation of Crude Brucella Protein Antigen
The CBP was prepared from B. abortus biotype 1 according to the previously described methods (Onate et al., 2000) with modifications. Briefly, B. abortus biotype 1 was inoculated onto brucella agar and incubated at 37°C for 7 days under 5% CO2. The culture was harvested in sterile Phosphate Buffered Saline (PBS) after 7 days of incubation. Bacteria were washed three times with the sterile PBS at 8000 rpm for 10 min at 4°C. Finally, it was resuspended in 50 mL of sterile PBS and inactivated by heating at 60°C for 1 h. The inactivated culture were sonicated at melting ice temperature, applying 6 cycles at 100 W, each cycle was of 1 min duration. The sonicated cell lysate was centrifuged at 1200 g for 20 min at 4°C. The supernatant was collected in 2 mL aliquots and stored at -20°C until tested. The protein concentration of the antigen was measured by using Bradford kit (Bio-Rad, Hercules, USA).
Lymphocyte Proliferation Assay
At 0, 3, 7, 14, 21, 28, 35, 42, 60, 90 and 120 days after infection, four rats from each group were euthanized and their spleens were removed under aseptic condition. Single-cell suspensions were prepared from these spleens according to a standard procedure (Onate et al., 2003). Briefly, single cell suspensions were prepared from rat spleens by dispersion of the tissue through a sterilized stainless steel. The spleen was meshed with Roswell Park Memorial Institute Medium (RPMI). The meshed spleen was centrifuged at 2000 rpm for 3 min. The supernatant was discarded. Lysis of the red blood cells in the sedimented spleenocytes was performed by Tris-buffered ammonium chloride (pH 7.2). After lysis of red blood cells the spleenocytes were centrifuged at 2000 rpm for 3 min and supernatant was discarded. Finally, 10 mL of complete RPMI-1640 media supplemented with 10% fetal calf serum, 2 mM L-glutamine, 100 IU mL-1 of penicillin, 100 μg mL-1 of streptomycin and 2 μg mL-1 of fungizone were added in the spleenocytes suspension. The concentration of the spleenocytes was adjusted in RPMI media at a concentration of 2x105 viable cells mL-1.
The lymphocyte proliferation response was determined by the colorimetric Thiazolyl Blue Tetrazolium Bromide (MTT) assay according to the method described by Mosmann (1983), with modifications. Briefly, 100 μL of splenic cell suspension was placed in triplicate (2x105 cells well-1) in 96-well tissue culture plates with 50 μL of medium alone or medium containing CBP (2 μg well-1) and Concanavalin A (ConA) 0.5 μg well-1. The cultures were incubated at 37°C in a humidified 5% CO2 atmosphere for 48 h, after which supernatant was removed by pipette. Then 50 μL of the MTT [3-(4, 5-dimethylthiazol-2-yl)-2, 5-diphenyl-tetrazolium bromide; Sigma] was added to each well. The cells were further incubated at 37°C for 4 h and then 100 μL of acid isopropanol (0.04 N HCl) was added to each well. The absorbance was measured at the wavelength of 570 nm with a microplate Enzyme-Linked Immunosorbant Assay (ELISA) reader (Tecan, Austria). Cellular proliferation was expressed as mean Stimulation Index (SI), calculated by dividing the mean Optical Density (OD) of the stimulated cultures by the mean OD of unstimulated control cultures.
Rose Bengal Plate Test
Sera collected from SD rats at 0, 3, 7, 14, 21, 28, 35, 42, 60, 90 and 120 days after infection were tested by RBPT using a commercial whole cell antigen of B. abortus 1119-3 (Deasung Micrbiological Labs, Korea) according to the previously described method (Alton et al., 1988).
Standard Tube Agglutination Test
Two-fold serial dilutions of sera collected at 0, 3, 7, 14, 21, 28, 35, 42, 60, 90 and 120 days after infection were tested by Tube Agglutination Test (TAT) using the whole cell antigen of B. abortus 1119-3 (National Veterinary Research and Quarantine Service, Anyang, Korea) according to the method of Alton et al. (1988). Reciprocal of the highest dilution of serum that showed clearing of the suspension and formation of a distinct agglutination mat at the bottom of the tube was expressed as the agglutination titer.
Enzyme-Linked Immunosorbent Assay
The presence of serum total immunoglobulin G (IgG) was determined by an indirect ELISA. The CBP was diluted to 10 μg mL-1 in 0.05 mM sodium bicarbonate buffer (pH 9.6) and used to coat the wells (100 μL well-1) of a flat-bottomed 96-well microtitre plate (Nunc, Denmark). Affinity purified Rat IgG (Bethyl laboratories, Inc., USA) were used to coat the 96-well plate starting from 500 to 7.8 ng well-1 for generation of standard curve. After overnight incubation at 4°C, plates were washed three times with wash solution (PBST: PBS pH 7.4) with 0.05% (V/V) Tween 20 and blocked with 1% Bovine Serum Albumin (BSA) (Sigma Aldrich Inc., St. Louis, MO, USA) in PBS for 30 min at 37°C. After three washes with PBST, 100 μL of control and test sera samples, diluted 1:500 in sample diluent (50 mM tris, 0.14 M NaCl ,1% BSA, 0.05% Tween 20, pH 8.0) were added to each well in duplicate. The plates were sealed and incubated at 37°C for 1 h. After five washing cycles with PBST, each well was incubated with 100 μL of 1: 100000 dilution of goat anti-rat IgG antibodies conjugated to horseradish peroxidase (Bethyl laboratories Inc., USA) diluted in conjugate diluent (50 mM tris, 0.14 M Nacl, 1% BSA, 0.05% Tween 20, pH 8.0). After 1 h incubation at 37°C, plates were washed 5 times as described above and the color reaction was developed by adding 200 μL well-1 of a solution containing 1.0 mg mL-1 of O-phenylenediamine dihydrochloride (OPD; Sigma, St. Louis, USA) in 0.05 M citrate buffer (pH 4.0) with 0.04% H2O2. After 30 min of incubation at room temperature, the enzyme reaction was stopped by addition of 50 μL of 3 M sulfuric acid/well and the absorbance of the developed color was measured at 492 nm, using an automatic ELISA plate reader (Tecan, Austria). The standard curve describing the relation between the concentration of standards and their absorbance value was generated and the concentration of antibody for each sample was expressed as ng mL-1.
Western Blot Analysis
Sodium dodecyl sulfate-polyacrylamide gel electrophoresis (SDS-PAGE) was performed in a mini-gel system (Bio-Rad, USA) under reducing conditions as described by Laemmli (1970). Briefly, the antigen was solubilized in sample buffer and subjected to SDS-PAGE using 12% polyacrylamide gels and analysed with Coomassie blue (2.5% brilliant blue in 50% methanol, 10% acetic acid) staining. For Western blotting, electrophoresed antigen was transferred to nitrocellulose membranes (0.45 um pore size, Bio-Rad, USA) at 100 V for 1 h under conditions essentially similar to that described by Towbin et al. (1979). Unbound sites on the membrane were blocked with PBS containing 0.2% Tween 20 at 4°C overnight. The blocking solution was poured off and the membrane was washed with PBST (0.1% Tween 20 in PBS) three times. The membrane was then reacted with sample serum diluted in PBS containing 0.05% Tween 20 for 1 h at rt. After three times rinse with PBS containing 0.1% Tween 20 for 30 min, phosphatase-labeled affinity purified antibody rat IgG (KPL, Europe) diluted in PBS at 1:1000 were reacted for 1 h at room temperature. After 3 times rinse as described above, the membrane were immersed in BCIP/NBT substrate (KPL, Europe) and the reaction was developed in darkness at rt for 10-15 min or until desired color is achieved. A last washing step was performed once with distilled water. The membrane was allowed to dry in the air. The thoroughly dried membrane was scanned for taking the image.
Statistical Analysis
The data were analyzed for statistical significance using Student’s two-tailed t-test. The p-value of <0.05 was considered to be significant.
RESULTS AND DISCUSSION
Clinical Observations
All of the rats inoculated with B. abortus biotype 1, developed lethargic, anorectic and febrile conditions within 24 h. The uninfected control rats did not manifest any of the abnormal clinical signs. The highest rectal temperature was 38.30±0.152°C in the rats of inoculated group and 36.5±0.05°C in the uninfected control group.
Cellular Immune Response
Sequential monitoring of lymphocytes responses to CBP revealed a progressive increase of the SI in the infected rats. The antigen specific cellular response increased from 7 days after infection with a SI of 3.25±0.25. This was followed by a persistent rise in the lymphocyte response of infected rats, with a peak SI of 6.5±0.28 at 28 days after infection, which were significantly different from lymphocyte responses recorded in the control group. The proliferative response showed a decrease on 60 and 90 days after infection followed by a second one on 120 days after infection (p<0.05). The ConA mitogen as positive control was able to induce T-cell proliferation in all cases (data not shown). The results of cellular immune responses are presented in Fig. 1.
RBPT Screening
Sera collected from rats at 7, 14, 21, 28, 35, 42, 60, 90 and 120 days after infection were tested positive for B. abortus by the RBPT. On the other hand, RBPT was negative for sera collected from rats at 0 and 3 days after infection.
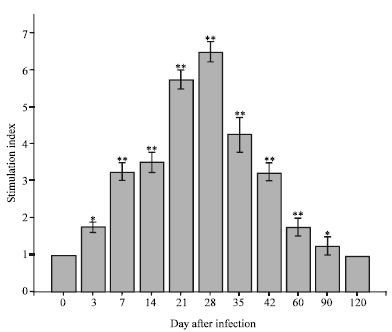 | |
| Fig. 1: | Lymphocyte proliferation assay in B. abotus biotype 1 infected SD rats at 0, 3, 7, 14, 21, 28, 35, 42, 60, 90 and 120 days after infection. Results are expressed as the Mean±SD. Statistically significant difference of stimuation index between uninfected control and infected rats at different time points of infection are indicated by asterisks (*p<0.05 and **p<0.001) |
TAT Antibody Titers
The data on TAT antibody responses of infected rats showed a remarkable increase in antibody titers after experimental infection. The TAT antibody titers at 7 days after infection was 125±25, but exhibited a five-fold increase to 625±50 on 35 days after infection. The antibody titers began declining until the end of the experiment. No antibody responses were noted in the control group. The mean reciprocal serum antibody titers measured by TAT are presented in Fig. 2.
ELISA Antibody Titers
In the infected rats, B. abortus specific IgG antibodies response started at 3 days after infection (150.9±14.67 ng mL-1). The IgG response rose significantly from 7 days after infection (509.26±26.07 ng mL-1). The highest serum IgG titer recorded at 35 days after infection (1213.27±9.06 ng mL-1) then the antibody titers decreased gradually until the end of the experiment. The results of IgG responses in the sera of infected rats are presented in Fig. 3.
Western Blot Analysis
The transfer of B. abortus antigens separated by electrophoresis onto a nitrocellulose gel revealed a large array of proteins with apparent molecular weight between 19 and 125 kDa. Pre-infection sera reacted weakly with the 24, 22 and 21 kDa proteins. Three days after infection 105, 54, 32 and 19 kDa proteins were recognized by the sera. At 7 days after infection, sera of the infected rats recognized 82, 32 and 19 kDa proteins. At 14 days after infection bands of 125, 46, 32 and 19 kDa protein were observed in the sera. Sera of 21 days after infection reacted with 82, 46, 32 and 19 kDa proteins. Protein bands of the 82, 46, 32 and 19 kDa reacted with sera collected after 28 days after infection. Sera collected at 35 days after infection reacted with 125, 105, 82, 46 and 32 kDa proteins. Five protein bands at the molecular weight of 125, 105, 82, 46 and 32 kDa reacted with sera collected at 42 days after infection.
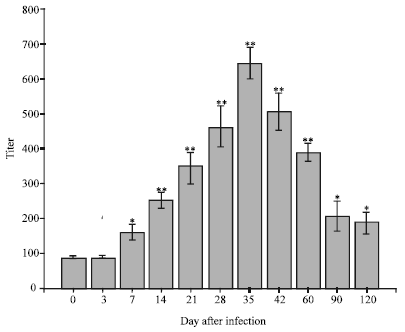 | |
| Fig. 2: | Serum antibody titers in B. abortus biotype 1 infected rats measured by TAT at 0, 3, 7, 14, 21, 28, 35, 42, 60, 90 and 120 days after infection. Results are expressed as the Mean±SD. Statistically significant difference of antibody titers between uninfected control and infected rats at different time points of infection are indicated by asterisks (*p<0.05 and **p<0.001) |
 | |
| Fig. 3: | Serum lgG antibody titers in the B. abortus biotype 1 infected rats mearsured by the indirect ELISA at 0, 3, 7, 14, 21, 28, 35, 42, 60, 90 and 120 days after infection. Results are expressed as the Mean±SD. Statistically significant difference of antibody titers between uninfected control and infected rats at different time points of infection are indicated by asterisks (*p<0.05 and **p<0.001) |
Sera at 60 days after infection reacted with 82 and 19 kDa proteins. Protein bands around 82, 66, 46 and 19 kDa molecular weight were observed in sera collected at 90 days after infection. Sera collected at 120 days after infection reacted with 46, 32 and 19 kDa proteins. The result of WB assay is shown in Fig. 4.
A wide range of wildlife is known to be the reservoir of B. abortus. Domesticated livestock animals might get infected with B. abortus by contact with the infected wildlife. Control of brucellosis in wildlife is necessary for eradication of bovine brucellosis. The cellular and humoral immunity in Brucella infection has always been a matter of interest for the researchers. The current understanding of immune response and antigen recognition against B. abortus has been arisen from the studies either in mouse or cattle. However, very little systemic information seems to be available on the measurement of immune response and antigen recognition in free ranging wildlife. We measured the cellular and humoral immune responses as well as antigen recognition induced by a virulent B. abortus biotype 1 in SD rats.
In the present study, experimentally infected rats mounted a lymphoproliferative and humoral response within one week after infection. We noticed lymphocyte stimulation response before the development of antibodies. Kaneene et al. (1979) using an extract of autoclaved B. abortus cells as antigen found that experimentally infected cattle developed cellular responses by 1 to 2 weeks post exposure and several days earlier than seroagglutinins could be detected. A study in a naturally infected herd confirmed that lymphocyte proliferative response preceded development of antibodies (Kaneene et al., 1978).
This study demonstrates that intraperitoneal infection of rat with B. abortus biotype 1 generates a strong specific humoral as well as T-cell responses. The induction of T-cell immune responses following B. abortus infection was evaluated by measuring T-cell-proliferative responses after in vitro stimulation of splenic cells with CBP.
 | |
| Fig. 4: | Western blot analysis of the rats sera using lgG.M. protein marker (kDa), lanes 1-11, corresponding to the sera collected at 0, 3, 7, 14, 21, 28, 35, 42, 60, 90 and 120 days after infection with B. abortus biotype 1, respectively |
Data of the lymphocyte proliferation assay suggests an initial high T-cell-proliferative response followed by a gradual decreased up to the end of the study. Onate et al. (2000) used CBP for measuring cellular immune response in mice and observed a higher lymphoproliferative response following single injection of CBP in the footpads of mice. Cabrera et al. (2009) measured lymphocyte proliferation response in B. abortus strain RB51 vaccinated mice by in vitro stimulation with CBP.
Using ELISA, several studies have been conducted to analyze the antibody response against proteins of Brucella sp. (Letesson et al., 1997). ELISA targeting selected cytoplasmic (Hemmen et al., 1995), periplasmic (Rossetti et al., 1996), or membrane (Cloeckaert et al., 1992; Zygmunt et al., 1994) proteins has been described. In the present study, IgG antibodies mediated humoral immune response against B. abortus biotype 1 has been measured by an indirect ELISA using CBP during the course of infection. We detected IgG antibodies response in the sera of SD rats at 3 days after infection with B. abortus biotype 1. In cattle, Beh (1973) recorded IgG antibodies responses at 7 days after infection with B. abortus.
In acute brucellosis, serum IgG response initially becomes low but with the progress of the infection the IgG antibody titers increase. In this study, the serum IgG antibodies measured by ELISA showed maximal antibody titers at 35 days after infection. Then the antibody titers gradually decreased from 35 days of infection to the end of the experiment. Similar pattern of IgG antibody responses to B. abortus under experimental conditions have previously been documented in BALB/c mice (High et al., 2007). On the contrary, in cattle, the IgG antibodies responses against B. abortus reached the peak value at 28 to 42 days after infection, after which they declined (Macmillan, 1990). The patterns of antibody responses recorded by the TAT in the sera of infected rat in our experiment were similar to that of ELISA.
The Brucella cell envelope is a three-layered structure in which an inner or cytoplasmic membrane, a periplasmic space and an outer membrane can be differentiated (Cloeckaert et al., 1990). In this study, we disrupted the B. abortus by sonication and cell free crude extract was obtained by centrifugation. The crude extract contains outer membrane, periplasmic as well as cytoplasmic proteins. We performed WB analysis of sera collected at 0, 3, 7, 14, 21, 28, 35, 42, 60, 90 and 120 days after infection using CBP to identify the immunoreactive antigens. WB assay has been used to identify the immunoreactive proteins of Brucella sp., which could be used as diagnostic antigens for animal or human brucellosis (Letesson et al., 1997). Among the immunodominant Brucella antigens identified by WB, some belong to the cell envelope and correspond to both major Outer Membrane Proteins (OMPs) (25 to 27 kDa and 36 to 38 kDa) and minor OMPs (10 kDa, 16.5 kDa, 19 kDa and 89 kDa) (Letesson et al., 1997). In this study, IgG antibodies present in the sera were reacted with protein bands at a molecular weight ranging between 19 and 125 kDa in WB assay. Six protein bands of 82, 46, 24, 22, 21 and 19 kDa were intensively recognized by almost all sera of infected rats. Protein bands of 125, 105, 66, 54 and 32 kDa were weakly recognized by the sera of the infected rats. The pattern of recognition of immunodominant bands by the sera of infected rats was almost similar between acute and sub-acute infections.
In this study, a few bands were weakly recognized by pre-infection sera collected at 0 day after infection. Cross reaction and background binding were reported especially when soluble antigens were used (Dubey et al., 1996; Harkins et al., 1998; Nishikawa et al., 2002; O’Handley et al., 2002). In the persent study, 105 kDa proteins were moderately recognized by sera collected at 3, 35 and 42 days after infection. Proteins band of 82 kDa was intensely recognized by sera collected at 7, 21, 28, 35, 42, 60 and 90 days after infection. The 46 kDa protein band was intensely recognized by the infected rat sera from 14 days after infection until the end of the experiment. Intense protein band of 19 kDa was identified by the sera collected at 21 days after infection and persisted up to 120 days after infection. Sera collected at 14 days after infection weakly recognized 125 kDa protein bands which were persisted until the end of the experiment. The protein band of 32 kDa was weakly recognized by sera collected at 7 days of infection and persisted until the end of the study.
Data of this study suggest that the anti-protein antibody responses were heterogeneous among infected animals and that only a combination of selected Brucella proteins could lead to a satisfactory diagnostic test (Limet et al., 1993; Tabatabai and Hennager, 1994; Hemmen et al., 1995). Based on frequency and intensity of recognition, the 82, 46, 32, 24, 22, 21 and 19 kDa proteins should be considered in rats as immuno-dominant B. abortus antigens which could be useful for the diagnosis of B. abortus infections.
REFERENCES
- Alton, G.G., L.M. Jones, R.D. Angus and J.M. Verger, 1988. Techniques for the Brucellosis Laboratory. 1st Edn., Institute Nationale de le Rech, France, Paris, Pages: 174.
Direct Link - Beh, K.J., 1973. Distribution of Brucella antibody among immunoglobulin classes and a low molecular weight fraction in serum and whey of cattle. Res. Vet. Sci., 14: 381-384.
PubMed - Cabrera, A., D. Saez, S. Cespedes, E. Andrews and A. Onate, 2009. Vaccination with recombinant Semliki Forest virus particles expressing translation initiation factor 3 of Brucella abortus induces protective immunity in BALB/c mice. Immunobiology, 214: 467-474.
CrossRef - Cloeckaert, A., P. Kerkhofs and J.N. Limet, 1992. Antibody response to Brucella outer membrane proteins in bovine brucellosis: immunoblot analysis and competitive enzyme-linked immunosorbent assay using monoclonal antibodies. J. Clin. Microbiol., 30: 3168-3174.
PubMed - Dubey, J.P., D.S. Lindsay, D.S. Adams, J.M. Gay, T.V. Baszler, B.L. Blagburn and P. Thulliez, 1996. Serologic responses of cattle and other animals infected with Neospora caninum. Am. J. Vet. Res., 57: 329-336.
PubMed - Harkins, D., D.N. Clements, J. Maley, J. Marks, S. Wright, I. Esteban, E.A. Innes and D. Buxton, 1998. Western blot analysis of the responses of ruminants infected with Neospora caninum and with Toxoplasma gondii. J. Comp. Pathol., 119: 45-55.
PubMed - Hemmen, F., V. Weynants, T. Scarcez, J.J. Letesson and E. Saman, 1995. Cloning and sequence analysis of a newly identified Brucella abortus gene and serological evaluation of the 17-kilodalton antigen that it encodes. Clin. Diagn. Lab. Immunol., 2: 263-267.
PubMed - High, K.P., R. Prasad, C.R. Marion, G.G. Schurig, S.M. Boyle and N. Sriranganathan, 2007. Outcome and immune responses after Brucella abortus infection in young adult and aged mice. Biogerontology, 8: 583-593.
PubMed - Kaneene, J.M.B., D.W. Johnson, R.K. Anderson and C.C. Muscoplat, 1978. Utilization of a specific in vitro lymphocytes immunostimulation assay as an aid in detection of Brucella-infected cattle not detected by serological tests. J. Clin. Microbiol., 8: 512-515.
Direct Link - Kaneene, J.M.B., R.D. Angus, D.W. Johnson, C.C. Muscoplat, R.K. Anderson and D.E. Pietz, 1979. Temporal cell-mediated immune responses of cattle following natural and experimental exposure to living Brucela abortus. Can. J. Comp. Med., 43: 132-141.
PubMed - Kittelberger, R., P.G. Bundesen, A. Cloeckaert, I. Greiser-Wilke and J.J. Letesson, 1998. Serological cross-reactivity between Brucella abortus and Yersinia enterocolitica 0:9: IV. Evaluation of the M- and C-epitope antibody response for the specific detection of B. abortus infections. Vet. Microbiol., 60: 45-57.
Direct Link - Laemmli, U.K., 1970. Cleavage of structural proteins during the assembly of the head of bacteriophage T4. Nature, 227: 680-685.
CrossRefDirect Link - Letesson, J.J., A. Tibor, G.V. Eynde, V. Wansard, V. Weynants, P. Denoel and E. Saman, 1997. Humoral immune responses of Brucella-infected cattle, sheep and goats to eight purified recombinant Brucella proteins in an indirect enzyme-linked immunosorbent assay. Clin. Diagn. Lab. Immunol., 4: 556-564.
PubMed - Lim, H.S., Y.S. Min and H.S. Lee, 2005. Investigation of a series of brucellosis cases in Gyeongsangbuk-do during 2003-2004. J. Prev. Med. Public Health, 38: 482-488.
PubMed - Limet, J.N., A. Cloeckaert, G. Dezard, J.V. Broeck and G. Dubray, 1993. Antibody response to the 89 kDa outer membrane protein of Brucella in bovine brucellosis. J. Med. Microbiol., 39: 403-407.
PubMed - Moore, C.G., and P.R. Schnurrenberger, 1981. A review of naturally occurring Brucella abortus infections in wild mammals. J. Am. Vet. Med. Assoc., 179: 1105-1112.
PubMed - Mosmann, T., 1983. Rapid colorimetric assay for cellular growth and survival: Application to proliferation and cytotoxicity assays. J. Immunol. Methods, 65: 55-63.
CrossRefPubMedDirect Link - Nicoletti, P., 1980. The epidemiology of bovine brucellosis. Adv. Vet. Sci. Comp. Med., 24: 69-98.
PubMed - Nishikawa, Y., F.G. Claveria, K. Fujisaki and H. Nagasawa, 2002. Studies on serological cross-reaction of Neospora caninum with Toxoplasma gondii and Hammondia heydorni. J. Vet. Med. Sci., 64: 161-164.
PubMed - O'Handley, R., S. Liddell, C. Parker, M.C. Jenkins and J.P. Dubey, 2002. Experimental infection of sheep with Neospora caninum oocysts. J. Parasitol., 88: 1120-1123.
Direct Link - Oliakova, N.V. and V.I. Antoniuk, 1989. The gray rat as a carrier of infectious agents in Siberia and the Far East. Med. Parasitol., 3: 73-77.
PubMed - Onate, A.A., S. Cespedes, A. Cabrera, R. Rivers and A. Gonzalez et al., 2003. A DNA Vaccine encoding Cu, Zn superoxide dismutase of Brucella abortus induces protective immunity in BALB/c mice. Infect. Immun., 71: 4857-4861.
Direct Link - Pappas, G., P. Papadimitriou, N. Akritidis, L. Christou and E.V. Tsianos, 2006. The new global map of human brucellosis. Lancet Infect. Dis., 6: 91-99.
CrossRefPubMedDirect Link - Rossetti, O.L., A.I. Arese, M.L. Boschiroli and S.L. Cravero, 1996. Cloning of Brucella abortus gene and characterization of expressed 26-kilodalton periplasmic protein: Potential use for diagnosis. J. Clin. Microbiol., 34: 165-169.
PubMed - Tabatabai, L.B. and S.G. Hennager, 1994. Cattle serologically positive for Brucella abortus have antibodies to B. abortus Cu-Zn superoxide dismutase. Clin. Diagn. Lab. Immunol., 1: 506-510.
PubMed - Towbin, H., T. Staehelin and J. Gordon, 1979. Electrophoretic transfer of proteins from polyacrylamide gels to nitrocellulose sheets: Procedure and some applications. Proc. Natl. Acad. Sci. USA., 76: 4350-4354.
PubMedDirect Link - Trujillo, I.Z., A.N. Zavala, J.G. Caceres and C.Q. Miranda, 1994. Brucellosis. Infect. Dis. Clin. North Am., 8: 225-241.
PubMed