
M.N. Sundararaman
Semen Bank, Department of Animal Genetics and Breeding, Madras Veterinary College, Tamilnadu Veterinary and Animal Sciences University, Chennai-600 007, India
J. Kalatharan
Semen Bank, Department of Animal Genetics and Breeding, Madras Veterinary College, Tamilnadu Veterinary and Animal Sciences University, Chennai-600 007, India
K. Thilak Pon Jawahar
Semen Bank, Department of Animal Genetics and Breeding, Madras Veterinary College, Tamilnadu Veterinary and Animal Sciences University, Chennai-600 007, India
Asian Journal of Animal and Veterinary Advances
Year: 2007 | Volume: 2 | Issue: 4 | Page No.: 196-204







ABSTRACT
To eliminate the subjectivity inherent in conventional methods, morphology evaluation of spermatozoa by morphometry using CASA technique was undertaken in bulls. In this study, 1219 properly digitized spermatozoa, 616 and 603 sperm for 2 clinically normal Jersey bulls identified by scanning the sperm slides prepared from 19 ejaculates were used for morphological and morphometric analysis by CASA technique. The total percentage of sperm abnormalities recorded was 24.4%, of which, the occurrence of tail abnormalities was more often than head and head and tail (in combination). The total and head abnormalities showed significant (p≤0.05) differences between bulls. Among the tail abnormalities the incidence of coiled tail was most frequent followed by bent tail and absent tail. The morphometric characteristics viz. major axis (μm), minor axis (μm), elongation (%), head area (μm2), perimeter (μm) and tail length (μm) were recorded as 9.53, 4.73, 49.5, 35.7, 23.7 and 61.14, respectively. The between bull variations for major axis, elongation, head area, perimeter and tail length were reflected in the morphological differences as well. The incidence of head abnormalities due to higher morphometric values was significantly (p≤0.05) more than abnormalities due to lower and/or higher and lower morphometric values. CASA technique facilitates identification of even small but significant variations in sperm head morphomtrey and certain types of morphological deviations namely total and head abnormalities between clinically normal bulls, which encourages further studies to explore the difference in fertility if any, between bulls.
PDF Abstract XML References Citation
How to cite this article
M.N. Sundararaman, J. Kalatharan and K. Thilak Pon Jawahar, 2007. Analyses of Morphological and Morphometrical Deviations of Bull Spermatozoa by Computer Assisted Semen Analysis Technique. Asian Journal of Animal and Veterinary Advances, 2: 196-204.
DOI: 10.3923/ajava.2007.196.204
URL: https://scialert.net/abstract/?doi=ajava.2007.196.204
DOI: 10.3923/ajava.2007.196.204
URL: https://scialert.net/abstract/?doi=ajava.2007.196.204
INTRODUCTION
The shape and size of spermatozoa are species-specific and differ between various wild animals such as deer (Soler et al., 2005; Esteso et al., 2006; Sundararaman et al., 2006), alpaca (Buendia et al., 2002) and farm animals like cattle (Gravance et al., 1996; Boersma et al., 2001; Beletti et al., 2005), sheep (Gravance et al., 1998; Sancho et al., 1998), goat (Gravance et al., 1995; Sundararaman and Edwin, 2004; Hidalgo et al., 2006), horse (Davis et al., 1993; Casey et al., 1997; Arruda et al., 2002) and companion animal, dog (Dahlbom et al., 1997; Rijsselaere et al., 2004). Existence of significant differences in sperm morphology between species suggests that dimensional characteristics of spermatozoa are genetically controlled.
The role of percentage of morphologically normal sperm of man and animals in fertility has been widely published. Normal sperm morphology may be an indicator of the fertility potential of a given male (Padrik and Jaakma, 2002; Esteso et al., 2006). The association of increased morphological abnormalities of spermatozoa with reduced reproductive efficiency has been reported in humans (Kruger et al., 1988; Davis et al., 1993), stallions (Bielanski and Kaczmarski, 1979; Jasko et al., 1990), goats (Skalet et al., 1988; Gravance et al., 1998) rams (Osinowo et al., 1988), pigs (Alm et al., 2006) and bulls (Saacke, 1970; Fitzpatrick et al., 2002; Walters et al., 2005).
In bulls, higher proportion of abnormal spermatozoa could be an indicator of genetically heritable fertility (Hafez, 1987). Occurrence of a specific structural abnormality of sperm affecting the whole population or a high percentage of the populations might be associated with infertility (Perotti et al., 1981). Morphometric analysis of sperm heads has been shown to be an indicator of in vitro fertility (Kruger et al., 1993; Thompson et al., 1994).
Despite spermatozoa are terminal, highly differentiated cells, whose multitude of attributes that are of relevance for fertilization cannot be easily assayed by one single test (Rodriguez-Martinez, 2003), spermiogrammic analysis is currently considered as the most desirable approach to evaluate the reproductive ability of bulls (Hafez and Hafez, 2000). Comprehensive semen evaluation tests include morphology estimation by visual approximation of shape and size of spermatozoa, which is often subjective. However, CASA systems have been shown to correctly recognize and digitize 91% of the spermatozoa encountered (Davis et al., 1992) and to classify sperm head morphology with 95% accuracy (Morruzzi et al., 1988) compared to manual video-microscopic evaluation (Casey et al., 1997). Therefore, morphological assessment of spermatozoa by objective methods could add more value to semen evaluation protocols. With this background, a study was conducted to assess the morphology of bull spermatozoa by morphometry using CASA and to analyze the incidence of different types of abnormalities and deviations in morphomtric characteristics.
MATERIALS AND METHODS
Semen Collection, Evaluation and Processing
Semen samples were collected from 2 clinically normal Jersey bulls, aged 2 years, by artificial vagina. The bulls were in regular semen collection schedule of twice a week. Two consecutive ejaculates were harvested from each bull in every collection schedule. A total of 19 ejaculates, 9 and 10, respectively from 2 bulls were used for CASA analysis. This study was carried out during the July-August-September 2006.
The semen samples were evaluated for ejaculate volume, sperm concentration and sperm motility by conventional methods. After evaluation equal volume of Tris-egg yolk-glycerol based diluent was added to the semen samples. The sperm motility was assessed by phase contrast microscope at 37°C. Only samples containing progressive sperm motility of 70% and above were included for the study. The semen samples were further extended so as to fix the sperm number as 30 million per insemination dose.
Preparation Sperm Slides
After the final dilution, one aliquot (1 mL) of extended semen was further diluted with tris buffer so as to reduce the sperm concentration to approximately 20 million mL-1. Sperm slides were prepared by placing a 4 μL drop on a clean grease free glass slide and dragging the drop across the slide. The slides were air dried and stained by using, STAT III Andrology Stain (Mid-Atlantic Diagnostics Inc. NJ). Dried smears were fixed by immersing in methanol for 30 sec. After air-drying, the slides were immersed in Xanthene dye solution for 60 sec. Again after air-drying, the slides were immersed in Thiazine dye mixture for 60 sec. The excess stain on edges were blotted during the staining operation and finally washed with distilled water and air-dried. Several such slides were prepared from each sample for CASA analysis.
| Table 1: | Category gates used in HT-IVOS metrix software for evaluation of bull spermatozoa |
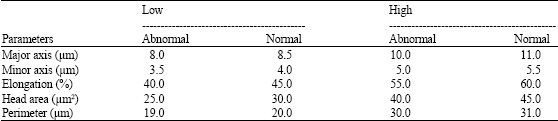 | |
CASA Analysis
Hamilton Thorne integrated visual optical system (HTM-IVOS) version 10.9 was the Computer Assisted Semen Analyzer (CASA) used in this experiment. The metrix software option for multi-species sperm morphology analysis was chosen for morphological and morphometrical analyses of bull sperm (Table 1). The sperm slide was loaded in the CASA system. The 60X objective was chosen (Gravance et al., 1996). After focusing, an image containing a spermatozoon was selected and the illumination was adjusted using adjust image option in order to obtain optimal illumination intensity (Rijsseaere et al., 2004). From each slide only properly digitized sperm heads were considered for morphometric measurement. Sperm heads were considered properly digitized when the computer-generated outline of the sperm head corresponded to the actual head outline. Using the reject option of the software, the non-sperm images scanned by the systems were eliminated from the analysis (Gravance et al., 1996). A total of 1219 properly digitized spermatozoa, 616 and 603, respectively for the 2 Jersey bulls identified by scanning from the sperm slides prepared from 19 ejaculates were used for morphometric analysis.
Major axis: the length of sperm head (μm); minor axis: the width of the sperm head (μm); elongation: the ratio of minor/major axis X 100 (%); head area: the total area of the sperm head (μm2); perimeter: the length of the sperm head perimeter (μm); tail length: the measured length of the tail (μm) were the morphometric characteristics measured.
Morphologically, a sperm cell was classified as normal or abnormal depending on the measured value for all the morphometric characteristics. If all the values were within the normal limits as indicated in the category gates of the CASA system, the spermatozoon was classified as normal provided the tail morphology of the sperm cell was also normal. If the morphometric values were in the abnormal range, the spermatozoon was designated as abnormal. The ranges of values used in metrix software to define bull spermatozoa as normal, abnormal and rejected are given in Table 1.
Classification of Data and Statistical Analyses
Based on the involvement of the part of the spermatozoa, the abnormalities were classified as head, tail and head and tail types. The abnormalities of tail as identified by the CASA system were classified as bent, coiled and absent tail. Further, the head abnormalities were sub-divided as high, low and high and low. If a sperm cell was identified as abnormal by the system because of higher head morphometric value(s), the head abnormality was classified as high and as low if the morphometric value was lower than the minimal value. If the abnormality was due to higher morphometric value in one parameter and lower in another in the same sperm cell, the head abnormality was classified as high and low. The data are presented as mean±SE. The values in percentages were subjected to analysis after arcsine transformation. The data were analyzed by one-way analysis of variance using microstat software (Ecosof Inc., 1984 Baltimore, USA). Significance of difference between means were determined at p≤0.05 and at p≤0.01 and the variations were designated as significant and highly significant, respectively.
RESULTS
The overall abnormality of spermatozoa was 24.4% (Table 2). The variations found between bulls for total sperm abnormalities in general and head abnormalities in particular were significant (p≤0.05). However, the variations between bulls for different types of tail abnormalities were not significant (Table 3).
Highly significant differences (p≤0.01) were noticed between bulls for major axis, elongation, head area, perimeter and tail length (Table 4).
Highly significant (p≤0.01) differences were observed between the incidences of types of sperm abnormalities in terms of location (Table 5). The proportion of abnormalities involving tail alone was the most, whereas the involvement of both head and tail in combination was the least. Table 6 shows the significant (p≤0.05) differences between the incidences of types of head abnormalities in terms of morphometric values. The head abnormalities due to higher morphometrics were more than for abnormalities because of other types of deviations.
| Table 2: | Mean percentages (±SE) for sperm abnormalities in Jersey bulls |
 | |
| Means bearing different superscripts in a column differ significantly (p≤0.05), n: No. of ejaculates | |
| Table 3: | Mean percentages (±SE) for types of tail abnormalities in spermatozoa of Jersey bulls |
 | |
| n: No. of ejaculates | |
| Table 4: | Means (±SE) for morphometric characteristics of bull spermatozoa of Jersey bulls |
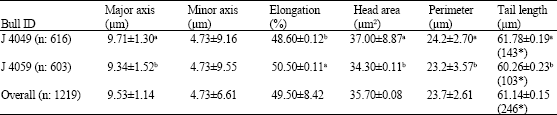 | |
| Means bearing different superscripts in a column differ significantly (p≤0.01), n: No. of spermatozoa analyzed for all morphometric characteristics except tail length, *: No. of spermatozoa analyzed for tail length | |
| Table 5: | Mean±SE for incidence of sperm abnormalities in terms of location, in Jersey bulls |
 | |
| Means bearing different superscripts differ significantly (p≤0.01), No. of ejaculates: 19 | |
| Table 6: | Means±SE for incidence of head abnormalities in terms of morphometric values of spermatozoa in Jersey bulls |
 | |
| Means bearing different superscripts differ significantly (p≤0.05), No. of ejaculates: 19 | |
| Table 7: | Means±SE for proportion of types tail abnormalities in spermatozoa of Jersey bulls |
 | |
| Means bearing different superscripts differ significantly (p≤0.01), No. of ejaculates: 19 | |
Regarding the abnormalities of tail (Table 7), the incidence of coiled tail was more than bent tail and absent tail. The variations between the occurrences of different tail abnormalities were highly significant (p≤0.01).
DISCUSSION
In this study a little more than 75% of spermatozoa analyzed were morphologically normal. These findings were obviously due to the fact that the bulls involved in the trail were selected and reared as Artificial Insemination (AI) sires and are in use for frozen semen production. Higher percentages of sperm with normal morphology recorded may suggest normal fertility as it appeared in an electron microscope study that a higher proportion of sperm bound to the zona pellucida were of normal morphology (Thundathil et al., 2001).
In fertile bulls the incidence of abnormal morphology of spermatozoa was found to be 10 to 18% (Lagerlof, 1934, 1936; Saacke et al., 1968; Rao et al., 1980). However, the higher sperm abnormalities (24.4%) observed in this trial may be because of different methods employed for morphological assessment. The computer automated system used in this trial, have been shown to classify sperm heads as normal or abnormal with 95% accuracy (Moruzzi et al., 1988) compared to manual video-microscopic evaluation (Casey et al., 1997).
The size of the sperm in terms of head length recorded in this trial for Jersey bulls is the largest (9.53 μm) when compared with the reports of Gravance et al. (1996), Boersma et al. (2001) and Beletti et al. (2005). The head area reported by Gravance et al. (1996) was smaller than that observed in this study and by other workers (Boersma et al., 2001; Beletti et al., 2005). The variations in the morphometric characteristics may be due to different breeds involved in the studies (Gravance et al., 1995). No report on bull sperm tail morphology could be traced to compare the results of this study. Nevertheless, the tail of bull sperm observed in this experiment is lengthier than the goat (Sundararaman and Edwin, 2004), dog (Rijsseleare et al., 2004) and deer (Sundararaman et al., 2006) spermatozoa.
Highly significant differences between bulls in major axis, elongation, head area, perimeter and tail length suggest that the morphometric characteristics varies between individual males. Within a species, estimates of the size of sperm heads vary greatly among stallions (Davis et al., 1993; Gravance et al., 1996; Casey et al., 1997) goats (Gravance et al., 1995; Sundararaman and Edwin, 2004) dogs (Dahlbom et al., 1997; Rijsseaere et al., 2004) rabbits (Gravance and Davis, 1995) alpaca (Buendia et al., 2002) and bulls (Gravance et al., 1996). The variation may be due to biological, such as inherited differences (Beatty, 1970) and stress factors influencing the condition of the male (Foote, 2003), DNA content associated with the sex chromosomes (Van Munster et al., 1999; Chandler et al., 2002) and incompletely condensed chromatin (Hingst et al., 1995). Despite the fact that overall head abnormalities in general was lower, it was more frequent in one of the bulls. The sperm head of the same bull was significantly larger. Further, most of the head abnormality in this study was due to larger sperm size in terms of morphometrics, which was significantly higher than the head abnormalities due to incidence of lower morphometrics and both higher and lower morphometrics in combination. Larger sperm heads are associated with subfertility (Boyle et al., 1992; Casey et al., 1997) since sperm with head abnormality have reduced capacity to bind ovum (Kot and Handel, 1987) and may lead to early embryonic loss, lowered fertility and embryo quality (Krzanowska and Lorenc, 1983; De Jarnette et al., 1992).
The incidence of tail abnormality was most frequent in this experiment and did show similarity in both the bulls. The occurrence of bent tail was similar to the earlier findings of Zimjanis (1969). This abnormality is often due to translocating protoplasmic droplet (Saacke, 1970; Gosch et al., 1989). Bent tail with entrapped protoplasmic droplet was reported to be the most common abnormality in bulls (Wildeus and Entwistle, 1984; Chacon, 2001). However, in the present experiment coiled tail was the most frequently encountered tail abnormality. This variation may be due to sub-classification of different tail abnormalities on microscopic studies by different workers. A fraction over 18% of tail abnormalities was identified as absent tail. The tail may be detached following ejaculation (Jasko et al., 1990; Rao, 1998) or prior to ejaculation in certain individual males. If the detachment occurs prior to ejaculation it may cause infertility (Jasko et al., 1990).
To conclude, the proportion of sperm abnormalities in this study is slightly higher than the threshold value of 20% (Hafez, 1987). Nevertheless, it is well within the permissible levels recommended by Society for Theriogenology (Chenoweth, 1997). The variation in sperm head size between bulls is to be viewed with caution, as it could be a sensitive biomarker related to fertility (Gravance et al., 1996; Sailor et al., 1996). Furthermore, the morphometrical differences are also reflected in the significant morphological differences between bulls. Little is known on the impact of small morphological differences on male fertility (Wijchman et al., 2001; Suzuki et al., 2002). Since CASA allows identification of minute differences between spermatozoa in morphometric measurements which would otherwise be unnoticed by traditional manual evaluation, the results obtained in this study on head morphometry can be utilized to explore differences in fertility if any, among bulls. Strict objective guidelines for evaluation sperm head morphology have to be evolved for bulls as in humans. Further research on CASA technique can contribute significantly towards achieving this goal.
REFERENCES
- Alm, K., O.A.T. Peltoniemi, E. Koskinen and M. Andersson, 2006. Porcine field fertility with two different insemination doses and the effect of sperm morphology. Reprod. Dom. Anim., 41: 210-213.
Direct Link - Arruda, R.P., B.A. Ball, C.G. Gravance, A.R. Garcia and I.K.M. Liu, 2002. Effects of extenders and cryoprotectants on stallion sperm head morphometry. Theriogenology, 58: 253-256.
Direct Link - Beatty, R.A., 1970. The genetics of the mammalian gamete. Biol. Rev., 45: 73-117.
CrossRefDirect Link - Beletti, M.E., F.C. Luciano da and M.P. Viana, 2005. A comparison of morphometric characteristics of sperm from fertile Bos taurus and Bos indicus bulls in Brazil. Anim. Reprod. Sci., 85: 105-116.
Direct Link - Bielanski, W. and F. Kaczmarski, 1997. Morphology of spermatozoa in semen from stallions of normal fertility. J. Reprod. Fertil., 27: 39-45.
PubMed - Boersma, A., R. Rabhofer and R. Stolla, 2001. Influence of sample preparation staining procedure and analysis conditions of bull sperm head morphometry using morphology analyzer integrated visual optical system. Reprod. Dom. Anim., 36: 222-229.
Direct Link - Boyle, C.A., M.J. Khoury, D.F. Katz, J.L. Annest, M. Kresnow, F. De-Stefano and S.M. Schrader, 1992. The relation of computer-based measures of sperm morphology and motility to male infertility. Epidemiology, 3: 239-246.
Direct Link - Buendia, P., C. Sloer, F. Paolicchi, G. Gago, B. Urquieta, F. Perez-Sanchez and E. Bustos-obregon, 2002. Morphometric characterization and classification of alpaca sperm heads using the sperm-class analyser7 computer assisted system. Theriogenology, 57: 1207-1218.
Direct Link - Casey, P.J., C.G. Gravance, R.O. Davis, D.D. Chabot and I.K.M. Liu, 1997. Morphometric differences in sperm head dimensions of fertile and subfertile stallions. Theriogenology, 47: 575-589.
Direct Link - Chacon, J., 2001. Assessment of sperm morphology in zebu bulls, under field conditions in tropics. Reprod. Dom. Anim., 35: 91-99.
Direct Link - Chandler, J.E., A.M. Canal, J.B. Paul and E.B. Moser, 2002. Collection frequency affects percent Y chromosome-bearing sperm head area and quality of bovine ejaculates. Theriogenology, 57: 1327-1346.
Direct Link - Dahlbom, M., M. Andersson, M. Vierula and M. Alanko, 1997. Morphometry of normal and teratozoospermic canine sperm heads using an image analyzer: Work in progress. Theriogenology, 48: 687-698.
Direct Link - Davis, R.O., D.E. Bain, R.J. Siemers, D.M. Thal, J.B. Andrew and C.G. Gravance, 1992. Accuracy and precision of the CellForm-Human automated sperm morphometry instrument. Fertil. Steril., 58: 763-769.
CrossRefDirect Link - Davis, R.O., C.G. Gravance and P.J. Casey, 1993. Automated morphometric analysis of stallion spermatozoa. Am. J. Vet. Res., 54: 1808-1811.
Direct Link - De-Jarnette, J.M., R.G. Saacke, J. Bame and C.J. Vogler, 1992. Accessory sperm: Their importance to fertility and embryo quality and attempts to alter their numbers in artificially inseminated cattle. J. Anim. Sci., 70: 484-491.
Direct Link - Esteso, M.C., M.R. Fernandez-Santes, A.J. Soler, V. Montoro, A. Quintero-Moreno and J.J. Garde, 2006. The effects of cryopreservation on the morphometric dimensions of Iberian Red Deer (Cervus elaphus hispancus) epididymal sperm heads. Reprod. Domes. Anim., 41: 241-246.
Direct Link - Fitzpatrick, L.A., G. Fordyce, M.R. McGowan, J.D. Bertram and V.J. Doogan et al., 2002. Bull selection and use in Northern Australia Part 2. Semen traita. Semen traits. Anim. Reprod. Sci., 71: 39-49.
Direct Link - Foote, R.H., 2003. Effect of processing and measuring procedures on estimated sizes of bull sperm heads. Theriogenology, 59: 1765-1773.
Direct Link - Gosch, B., T. Bartolomaeus and K. Fischer, 1989. Light and scanning electron microscopy of fallow deer (Dama dama) spermatozoa. J. Reprod. Fertil., 87: 187-192.
CrossRefDirect Link - Gravance, C.G. and R.O. Davis, 1995. Automated sperm morphometry analysis (ASMA) in the rabbit. J. Androl., 16: 88-93.
CrossRefDirect Link - Gravance, C.G., K.M. Lewis and P.J. Casey, 1995. Computer automated sperm head morphometry analysis (ASTHMA) of goat spermatozoa. Theriogenology, 44: 989-1002.
Direct Link - Gravance, C.G., R. Vishwanath, C. Pitt and P.J. Casey, 1996. Computer automated morphometric analysis of bull sperm heads. Theriogenology, 46: 1205-1215.
Direct Link - Gravance, C.G., Z.J. Champion and P.J. Casey, 1998. Computer-assisted sperm head morphometry analysis (ASTHMA) of cryopreserved ram spermatozoa. Theriogenology, 49: 1219-1230.
Direct Link - Hingst, O., S. Blottner and C. Franz, 1995. Chromatin condensation in cat spermatozoa during epididymal transit as studied by aniline and acridine orange staining. Andrologica, 27: 275-279.
Direct Link - Jasko, D.J., D.H. Lein and R.H. Foote, 1990. Determination of the relationship between sperm morphologic classifications and fertility in stallions: 66 cases (1987-1988). J. Am. Vet. Med. Assoc., 197: 389-393.
Direct Link - Kot, M.C. and M.A. Handel, 1987. Binding of morphologically abnormal sperm to mouse egg zonae pellucidae in vitro. Mol. Reprod. Dev., 18: 57-66.
CrossRefDirect Link - Kruger, T.F., A.A. Acosta, K.F. Simmons, R.J. Swanson, J.F. Matta and S. Oehninger, 1988. Predictive value of abnormal sperm morphology in in vitro fertilization. Fertil Steril., 49: 112-117.
PubMed - Kruger, T.F., T.C. DuToit, D.R. Franken, A.A. Acosta, S.C. Oehninger, R. Menkveld and C.J. Lombard, 1993. A new computerized method of reading sperm morphology (strict criteria) is as efficient as technician reading. Fertil. Steril., 59: 202-209.
CrossRefDirect Link - Krzanowska, H. and E. Lorenc, 1983. Influence of egg investments on in-vitro penetration of mouse eggs by misshapen spermatozoa. J. Reprod. Fertil., 68: 57-62.
CrossRefDirect Link - Rijsselaere, T., A.V. Soom, G. Hoflaqck, D. Maes and A. De-Kriuf, 2004. Automated sperm morphometry and morphology analysis of canine semen by the Hamilton-Thorne analyzer. Theriogenology, 62: 1292-1306.
Direct Link - Rodriguez-Martinez, H., 2003. Laboratory semen assessment and prediction of fertility: Still Utopia? Reprod. Dom. Anim., 35: 312-318.
PubMedDirect Link - Sailor, B.L., L.K. Jost and D.P. Evenson, 1996. Bull sperm head morphometry related to abnormal chromatin structure and fertility. Cytometry, 24: 167-173.
Direct Link - Sancho, M., F. Perez-Sachez, L. Tablado, J.J. de Nmonserrat and C. Soler, 1998. Computer assisted morphometric analysis of ram sperm heads: Evaluation of different fixative techniques. Theriogenology, 50: 27-37.
Direct Link - Soler, C., B. Gadea, A.J. Soler, M.R. Fernandez-santos and M.C. Esteso et al., 2005. Comparison of three different staining methods for the assessment of epididymal red deer sperm morphometry by computerized analysis with ISAS7. Theriogenology, 64: 1236-1243.
Direct Link - Sundararaman, M.N. and M.J. Edwin, 2004. Morphometric evaluation of Caprine spermatozoa by HT-IVOS system at pre-freeze and post-thaw stages of cryopreservation. Indian J. Anim. Reprod., 25: 91-93.
Direct Link - Suzuki, T., H. Shibahara, H. Tsunoda, Y. Hirano, A. Taneichi, H. Obara, S. Takamizawa and I. Sato, 2002. Comparison of the sperm quality analyzer II C variables with the computer-aided sperm analysis estimates. Int. J. Androl., 25: 49-54.
Direct Link - Thompson, L.A., P.F. Brook, M.A. Warren, C.L.R. Barratt and I.D. Cooke, 1994. A morphometric comparison of the nuclear morphology of fresh and frozen-thawed human zona-bound and unbound sperm. J. Androl., 15: 337-342.
Direct Link - Thundathil, J., J. Palamino, A. Barth, R. Mapletoft and C. Barros, 2001. Fertilizing characteristics of bovine sperm with flattened or indented acrosomes. Anim. Reprod. Sci., 67: 231-243.
Direct Link - Van-Munster, E.B., J. Stap, R.A. Hoebe, G.J. te Meerman and J.A. Aten, 1999. Difference in volume of X and Y chromosome-bearing bovine sperm heads matches difference in DNA content. Cytometry, 35: 125-128.
Direct Link - Walters, A.H., W.E. Eyestone, R.G. Saacke, R.E. Pearson and F.C. Gwazdauskas, 2005. Bovine embryo development after IVF with spermatozoa having abnormall morphology. Theriogenology, 63: 1925-1937.
Direct Link - Wijchman, J.G., B.T. De-Wolf, R. Graafe and E.G. Arts, 2001. Variation in semen parameters derived from computer-aided semen analysis, within donors and between donors. J. Androl., 22: 773-780.
Direct Link