
F.J. Al-Saffar
Department of Anatomy, Histology and Embryology, College of Veterinary Medicine, Baghdad University, Baghdad, Iraq
Eyhab R.M. Al-Samawy
Department of Anatomy, College of Medicine, Al-Muthana University, Iraq
Asian Journal of Animal Sciences
Year: 2015 | Volume: 9 | Issue: 6 | Page No.: 280-292







ABSTRACT
The study aimed to find out the histomorphological and histochemical structures of the walls of the stomach parts in mallard. To conduct this investigation, twelve healthy birds were collected from local suppliers at Baghdad province. Later on, birds were subdivided into three equal subdivisions, each of four for gross, histological and histochemical approaches. All birds were anesthetized and their abdomens were dissected, then their stomach were identified and photographed in situ. Organs were grossly described and specimens were immersed either in 10% neutral buffered formalin or in Bouin’s solution for histological fixation and then processed for histological and histochemical staining techniques, which were comprised Hematoxylin-eosin, Masson Trichrome, Periodic Acid Schiff (PAS) and PAS-Alcian Blue (AB) (pH 2.5). The gross findings showed that the stomach was consisted of two distinguishable chambers, that were proventriculus and ventriculus. The proventriculus was tubular-shaped organ. The ventriculus appeared fusiform in shape and covered by thin fatty tissue. There was well demarcation between proventriculus and ventriculus. Histologically, the proventriculus mucosa lining was simple columnar epithelium. The submucosal consists of numerous and voluminous deep submucosal glands. The covering of the ventriculus was simple cuboidal. Presence of thick cuticle layer with well-developed muscular stomach. Muscularis mucosa appeared characteristically thick layer of smooth muscle bundles arranged as, inner circular and outer longitudinal layers. The tunica muscularis appeared, as a very thick structure of smooth muscles fibers. Three layers of muscles were distinguished that were thin inner, outer longitudinal and very thick intermediate circular layers.
PDF Abstract XML References Citation
Received: July 11, 2015;
Accepted: August 20, 2015;
Published: September 18, 2015
How to cite this article
F.J. Al-Saffar and Eyhab R.M. Al-Samawy, 2015. Histomorphological and Histochemical Studies of the Stomach of the Mallard (Anas platyrhynchos). Asian Journal of Animal Sciences, 9: 280-292.
DOI: 10.3923/ajas.2015.280.292
URL: https://scialert.net/abstract/?doi=ajas.2015.280.292
DOI: 10.3923/ajas.2015.280.292
URL: https://scialert.net/abstract/?doi=ajas.2015.280.292
INTRODUCTION
The bird’s stomach shows distinct differentiation into two parts, the upper one, or the glandular stomach and the lower part or the gizzard. Both parts are characterized by a great morphological and functional variability, both between and within species. It is mostly influenced by accessibility amount and kind of food changing in time (Starck, 1999).
The glandular stomach (proventriculus) varies in size between species, being relatively small in graminivores but often quite large and distensible in carnivores that ingest large food items (Duke, 1997). In the black-winged kite (Elanus caeruleus), the stomach found large in size and possessed thin wall compared to seed eating avian species. The proventricular-ventricular isthmus was absent, so that the two organs form one large pear-shaped cavity (Hamdi et al., 2013).
The wall of the glandular part of the avian stomach, which produces gastric juice was constructed of the known usual layers of a typical tubular organ comprising mucosal, submucosal, muscular and serosal layers (Rossi et al., 2005; Hassouna, 2001). Generally, the mucosa was made of three constituents, that were epithelium, lamina propria and muscularis mucosa. Catroxo et al. (1997) established that the mucous membrane of the proventriculus in the red-capped cardinal consists of mucosal folds in its luminal surface, which covered by epithelium of simple prismatic cells. Its propria was structured by the connective tissue with blood vessels, infiltrated with lymphocytes. In its interior part present simple tubular glands lined by the same type of the epithelium. Incipient muscularis mucosa observed, as isolated muscular smooth fibers together with the lamina propria. The submucosa was constructed by connective tissue, containing blood vessels and the deep proventricular glands, which were conic or pear in shape occupied most thickness of the proventriculus wall. The muscular layer, presents an inner and outer longitudinal smooth muscle fibers with an intermediary of circularly disposed fibers. The serosa of the was constituted by connective tissue lined by the mesothelium.
In Japanese quail, the proventricular mucous membrane was simple epithelium of columnar cells with vesicular shaped nuclei basally located with acidophilic cytoplasm. These cells possessed supra-nuclear granules that were positively reacted with both PAS and AB stains. Underlined lamina propria contained infiltrated lymphocytes and lymphatic aggregations. The submucosa poorly developed and the muscular tunnic was made up by inner thick circular and outer thin longitudinal smooth muscular fibers (Ahmed et al., 2011).
Proventriculus of the Blue and Yellow macaws which were member of herbivores avian species, Rodrigues et al. (2012) stated that its wall was formed greatly by proventricular glands. The mucosa formed of simple cells of prismatic or cylindrical type. The muscularis mucosa sublayer appeared as, diffuse, longitudinally arranged fibers interspersed between the connective tissue located between the lobes of the glands. The submucosal layer was a thin band of loose connective tissue. The muscular tunic consisted only circularly arranged smooth muscle fibers.
Recently, it was found that the mucosa of the proventriculus in the quail (Coturnix coturnix) thrown into folds with varying heights that were lined by a simple columnar epithelium. Histochemically, the cells of the epithelial lining of the mucosal folds were reacted positively with PAS due to the presence of mucin granules in the supra nuclear area of these cells. The lining cells of the ducts of the proventricular glands showed PAS positive reaction in their apical ends. Whereas, the secretory cells were negatively stained with PAS stain (Zaher et al., 2012).
The wall of proventriculus in chicken found very thick and consisted of the following layers i.e., inner mucosal membrane, submucosa, tunica muscularis and outer serosa. The submucosal glands formed the greater part of the thickness of this organ. The proventriculus was consisted of macroscopic papillae with numerous microscopic folds. Simple single glands grouped to form lobules each of which converged into a common cavity near the surface. The cavities converged to form a common duct that lead to the surface through the apex of a small papilla. The surface epithelium on the folds was a simple columnar tissue and each fold had a core of lamina propria to support it (Nasrin et al., 2012).
Rossi et al. (2005) observed in the gizzard’s mucosa of the partridge (Rhynchotus rufescens) low folds lined by simple columnar to cuboidal epithelium. The mucosa revealed the presence of tubular glands, which were lined by low cuboidal at their bases, whereas higher at their upper portions. In fact, the luminal surface of the gizzard was lined with secretory product of the mucosal glands, which solidified at the surface to form a hard cuticle of koilin.
Selvan et al. (2008) recorded that the gizzard’s wall of the guinea fowl (Numida meleagris) constructed from the usually known four tunics in addition to an internal lining of koilin which was a secretary layer above the mucosa. The koilin showed positive reaction with the PAS. The surface epithelium was PAS positive too. They showed predominance of neutral mucin. The PAS positive material was present in the lumen of the glands and in the cells that were lined both the surface and crypts.
In the stomach of Japanese Quail, the gizzard’s wall was represented by mucosa revealed branched tubular glands, submucosa, muscular layer and serosa. The mucosa formed folds which were continuous with the tubular glands located in the underlining lamina propria. Absence of muscularis mucosa and the gizzard’s glands were lined by cuboidal cells. The lining epithelium was simple columnar or cuboidal cells characterized by basally located rounded or oval nuclei and basophilic cytoplasm. Apical portions of these lining cells were stained positively by PAS and AB stains. Keratinized laminated coat covered the mucosa which reacted positively toward acid fuchsin after using trichrome stain (Ahmed et al., 2011).
The gizzard in chicken found highly muscular adapted well for grinding and macerating the ingesta. The cuticle observed as pattern of wavy lines running parallel to the mucosal surface. The surface lined by simple columnar tissue but the mucosal glands were simple cuboidal branched tubular glands. The lamina propria and submucosa consisted of loose connective tissue. The tunica muscularis was strongly thick and constructed of parallel smooth muscle fibers covered by the outer serosa (Nasrin et al., 2012). There were paucity of work studied the histomorphological and histochemical structures of the mallard’s stomach and to our knowledge there were no local studies yet. Accordingly, the current study intended to do so that using mallards (Anas platyrhynchos).
MATERIALS AND METHODS
Birds collection and study design: Twelve Mallards (Anas platyrhynchos) were used to conduct the current study. These birds were bought from specific markets at Baghdad province from the local suppliers. Birds were housed at animal house of the Veterinary Medicine College/Baghdad university in suitable cages. They were fed as well and giving them water ad libitum before their euthanasia and dissection. The total number of birds were divided into three equal subdivisions each of four, for gross, histological and histochemical approaches.
Birds dissection and morphology study: Birds were euthanized prior to its dissection with an intravenous injection of sodium pentobarbitone (80 mg kg–1), then after, dissected by fixing them on a dissecting board. A mid-line incision was made in the abdominal wall of each bird used in this study to view the coelomic viscera. The proventriculus and ventriculus (gizzard) were identified and photographed in situ using digital camera (pupil cam., ken-a-vision). Locations and relationships of these organs was well illustrated in figures. The organs then after washed by normal saline solution to remove blood or other adhering debris. The contents of the stomach were removed gently by washing with normal saline.
Histological processes for the collected specimens: For the histological aspect of the study, the specimens (whole proventriculus, gizzard) from each dissected birds were fixed in 10% neutral buffered formalin. After well fixation the specimens were dehydrated by passing them through a series of ascending ethanol alcohol each for two hours (70, 80, 90 and 100%) and then specimens were cleared in xylene for one hour after that embedded in paraffin wax and then the blocks were sectioned at 6 μm thickness and stained with either one of the following stains: Mayer’s Hematoxylin and Eosin routine stain for general features identification, Masson Trichrome stain for the staining of the collagenous and smooth muscle fibers (Bancroft and Stevens, 2010).
Histochemical processes for the collected specimens: To conduct the histochemical study for the stomach, specimens were fixed in Bouin’s solution and then sections of 6 μm thickness were prepared and then after were stained with one of the bellow stains and subsequently examined and photographed by Olympus BH-2 microscope, using Dino-eye camera. For the determination of neutral and acidic mucin, PAS-Alcian Blue (AB) pH 2.5 was used. The PAS alone was used for the illustration of the basement membranes of the epithelial lining of the stomach and small intestine.
RESULTS AND DISCUSSION
Gross findings: The stomach in the studied bird appeared as an organ distinctly divided into two parts that were proventriculus and ventricular (Fig. 1) and such trait similarly observed in most avian species, such as; domestic fowl (Gallus gallus) (Akester, 1986), partridge (Rhynchotus rufescens) (Rossi et al., 2005) and Japanese quail (Ahmed et al., 2011). Conversely, Abumandour (2014) cited that stomach in some birds consists of three compartments; namely proventriculus, ventriculus and pyloric part.
Current observations revealed that the proventriculus was large, fusiform-shaped organ in mallard (Fig. 1). Similar findings were observed previously by Salem and Yousria (2000) in the proventriculus of the chicken, pigeon and duck. Whereas, the latter researchers differently described the proventriculus as lens-shaped organ in cattle agent avian species. On the same year, Whittow (2000) stated that the shape of the proventriculus is fusiform in the most avian species but with the differences in their sizes.
Macroscopic examination showed thick wall and distinct demarcation between the proventriculus and esophagus cranially and the gizzard caudally (Fig. 1). The well demarcation proventriculus in mallard was in accordance with previous findings observed by Pesek (1999) and Konish (2001) in seed eater birds such as Streaky and Paramo.
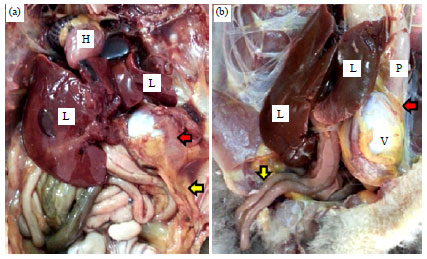 | |
| Fig. 1(a-b): | Viscera of mallard showed in situ the H: Heart , L: Liver, P: Proventriculus, V: Ventriculus, duodenum (yellow arrow) and demarcation (red arrow) |
Conversely, not well demarcated proventriculus from the esophagus and gizzard was described in meat eater birds, such as; Hawks and Eagles. Dissimilar to mallard, difficult demarcation of the esophagus-proventriculus junction was also found in case of most fowl (Baumel and King, 1993), in bustards (Bailey et al., 1997) and in partridge (Rossi et al., 2005).
The proventriculus in mallard was located in the left ventral part of the coelomic cavity lying between the esophagus cranially and the gizzard caudally (Fig. 1). The organ was partially covered by the left lobe of the liver. Likewise similar location of the proventriculus was found in other birds, such as; Ostrich (Tadjalli et al., 2011), common quail (Coturnix coturnix) (Zaher et al., 2012) and falcon (Abumandour, 2014).
The second parts of the stomach, which is the ventriculus appeared large in size and had pearl shape and covered by thin fatty tissue (Fig. 1). In the mallard which is one of the omnivorous species, ventriculus was markedly large in size with thick muscular wall because of its capability to eat fish, herb and bread but not with previous findings that the ventriculus is flattened-shaped organ and generally not covered with fatty tissue (Abumandour, 2014).
The constricted area between the two parts of the stomach, which was clearly visible in mallard (Fig. 1) was previously observed and mentioned in seed eater avian species such as brown-rumped (Serinus tristriatus) and white-collared (Sporophila torqueola) (Pesek, 1999; Konishi, 2001).
The ventriculus of the studied bird was located at the left side to the midline plane next to the proventriculus and covered partially by the left lobe of the liver (Fig. 1). Macroscopic examination of the organ revealed thick and well- developed muscular wall in mallard, because of the nature of their food, which is mostly the swallowed seeds. Such thickness gave rise to powerful muscular organ in these birds for grinding the seeds, which may mix with enzyme secreted by the proventriculus of such enzyme probably the pepsin (Schweizer et al., 2014). Grossly, the internal lining of the ventriculus in mallard was covered by a layer of cuticle which is probably established hard mucosa aid in physical digestion. These findings were in accordance with those of King and Mclelland (1984), whom observed well developed muscularity and cuticle in both granivorous and herbivorous species, such as; domestic fowl, pigeon, ducks and geese and differently they documented thin walled ventriculus in both owls and hawk.
Histological findings: Proventriculus: Microscopic examination of the wall of proventriculus revealed the presence of four layers of the typical tubular organ, that were tunica mucosa, submucosa, muscularis and serosa (Fig. 2). The layers, which structured the wall of proventriculus was similarly documented in the proventriculus of most avian species, such as; Ostrich (Struthio camelus) (Cooper and Mahroze, 2004), Guinea fowl (Numida meleagris) (Selvan et al., 2008), quail (Attia, 2008), Japanese quail (Ahmed et al., 2011) and Coot bird (Batah et al., 2012). However, Rocha and De Lima (1998) cited only three tunicae in the wall of proventriculus in which only tunica mucosa, tunica muscularis and tunica serosa were detected.
In mallard the mucosa showed longitudinal branched folds that were lined by simple columnar epithelium (Fig. 2). The underlying lamina propria was constructed of loose connective tissue filled by blood vessels and infiltrated lymphatic cells (Fig. 2). The structure of the lamina extended inside the folds and possessed simple tubular mucous glands (Fig. 2). These glands were opened into the lumen of the proventriculus via their ducts. The glands were lined with simple cuboidal epithelium and were dispersed at the apical part of the lamina propria. The propria was separated from the underlying submucosa by fibers of smooth muscle called muscularis mucosa (Fig. 2 and 3). These fibers were surrounded the apical parts of the submucosal glands (Fig. 3).
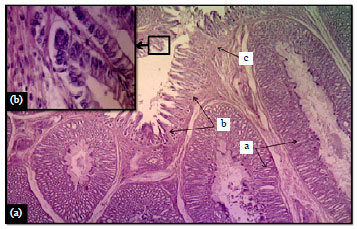 | |
| Fig. 2(a-b): | (a) Proventriculus wall of mallard showed a: T. mucosa, b: T. submucosa, c: Muscularis mucosa, H and E, X100 and (b) Higher magnification showed epithelial cells in mucosa, H and E, X400 |
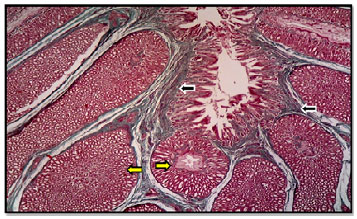 | |
| Fig. 3: | Proventriculus wall of mallard showed the green colored connective tissue in lamina propria (black arrows) and submucosal glands (yellow arrows), Masson’s Trichrome, X400 |
The mucosal glands which were observed lined with simple cuboidal epithelium in mallard were in accordance with those observed in the same organ of the Coot bird (Fulica atra) (Batah et al., 2012) and common quail (Coturnix coturnix) (Zaher et al., 2012). The presence of muscularis mucosa separating markedly the mucosa from the underlying submucosa in the mallard was in a good agreement with those observed in the same organ of the red-gartered coot (Fulica armillata) (Espinola and Galliussi, 1990) and the red jungle fowl (Kadhim et al., 2011).
Tunica submucosa, prominently found occupying most of the wall thickness of proventriculus. It consists of numerous deep glands called submucosal glands or proventriculus glands, which occupied the great part of the submucosa (Fig. 4). These findings were not parallel with King and Mclelland (1984), whom referred to the absence of proventriculus glands in the submucosa of proventriculus in chicken.
Submucosa was formed of loose connective tissue containing proventriculus glands which were rounded-shaped branched tubular glands surrounded by a fibrous capsule (Fig. 3) and such findings were not similar to those of Espinola and Galliussi (1990) and Rocha and De Lima (1998), whom mentioned that the glands were oval and lined by tall columnar epithelium.
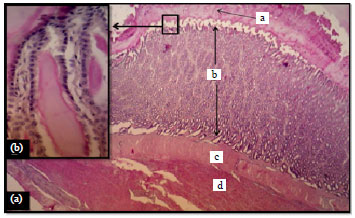 | |
| Fig. 4(a-b): | (a) Ventriculus wall of Mallard showed a: Cuticle b: T. mucosa c: T. submucosa and d: T. muscularis H and E, X100 and (b) Higher magnification showed epithelial cells in mucosa, H and E, X400 |
The glands consists numerous secretory tubules which were lined by cuboidal cells and each tubule continued by one duct opened into the main collecting duct which subsequently opened into luminal surface of the organ as observed similarly by Catroxo et al. (1997) in Red-Capped Cardinal (Paroaria gularis) and Batah et al. (2012) in Coot bird (Fulica atra) whom mentioned this glands located in submucosal layer.
Tunica muscularis constructed of two layers, inner thin longitudinal and an outer thick circular layers. Between such layers, fine connective tissue was observed invested with blood vessels (Fig. 4). This finding differ with in owl (Rocha and De Lima, 1998), in partridge (Rossi et al., 2005) noted that a well-developed tunica muscularis is arranged as inner circular and outer longitudinal layers of smooth muscle. In parrots (Denbow, 2000) added that the outer longitudinal layer is absent.
Tunica serosa was constructed of loose connective tissue in which nerves, blood vessels, adipose cells were observed and such structures were covered by a layer of mesothelium. These findings were similarly observed by Ahmed et al. (2011) in Japanese quail and Batah et al. (2012) in Coot bird (Fulica atra).
Ventriculus (gizzard): Similarly to the proventriculus, the microscopic structure of this organ also showed the four known tunicae forming its wall (Fig. 4). Same findings regarding the wall structure were found in Red-Capped Cardinal (Paroaria gularis gularis) (Catroxo et al., 1997), yellow-necked mouse (Apodemus flavicollis) and in guinea fowl (Numida meleagris) (Selvan et al., 2008).
Tunica mucosa was covered by simple cuboidal epithelium characterized by basally located oval-shaped nuclei with lightly stained cytoplasm (Fig. 4). The simple cuboidal cells covering was dissimilarly observed columnar in other avian species, such as; partridge (Rhynchotus rufescens) (Rossi et al., 2005) in Red jungle fowl (Kadhim et al., 2011) and in the Blue and Yellow macaws (Rodrigues et al., 2012). Whereas, the simple cuboidal covering, which observed in the ventriculus of mallard was similarly observed in other birds, such as; Blue and Yellow macaws (Rodrigues et al., 2012), in Coot bird (Fulica atra) (Batah et al., 2012).
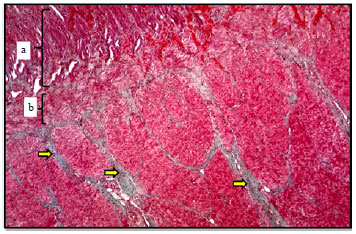 | |
| Fig. 5: | Ventriculus wall of mallard showed, (a): Mucosal layer and (b): Muscularis mucosa, green colored connective tissue between the muscular bundles of tunica muscularis (yellow arrows), Masson’s Trichrome, X400 |
The mucosal epithelium was thrown into folds (Fig. 4). The lamina propria showed numerous simple tubular glands lined by simple cuboidal cells. The examination of the ventriculus of mallard revealed the presence of eosinophilic secretion going away toward the epithelial surface as a strips forming the cuticle (Fig. 4 and 5). It spread all over the mucosal surface filling the lumen of the gastric pits as a pinkish thick material.
The color and the presence or absence of the cuticle was previously documented in avian species. The previous data indicated a relationship between it and the type of food consumed by the bird. Many researchers, such as; Bailey et al. (1997) and Gionfriddo and Best (1996) referred to the thickness of the cuticle, which is highly correlated with food consumed. They proposed thick cuticle in granivores and a thin in frugivores and nectarivores, which are often green, brown or yellow in color.
Actually, the cuticle serves as a grinding surface, enabling the contracted muscular ventriculus to mechanically digest the consumed food. So that in insectivorous, herbivorous and granivorous birds possess a well-developed muscular ventriculus with a thick abrasive cuticle layer. In contrast, birds that eat foods not requiring significant mechanical digestion, such as; carnivores and piscivores possessed an almost rudimentary ventriculus and soft cuticle (Akester, 1986; Lumeij, 1994; Denbow, 2000).
Muscularis mucosa appeared characteristically thick layer of smooth muscle bundles arranged as inner circular and outer longitudinal in mallard (Fig. 5). The presence of muscularis mucosa between the mucosa and submucosa in the studied birds appeared dissimilar to previous findings in other birds, such as; Blue and Yellow macaws (Ahmed et al., 2011; Rodrigues et al., 2012) and in Codorna nothura (Fieri, 1984) in which this layer was absent. In stomach of the Japanese quail (Ahmed et al., 2011) that stated the mucosal gland interferes with propria.
Tunica submucosa of the ventriculus composed of abundant connective tissue containing blood vessels and nerves (Fig. 5). Tunica muscularis appeared as a very thick structure of smooth muscles fibers. three layers of muscles were distinguished that were thin inner, outer longitudinal and very thick intermediate circular layers. The presence of three layers of muscles fibers was in accordance with the findings of Kadhim et al. (2011) in the ventriculus of the red jungle fowl. While differently, presence only two layers of muscles fibers in the ventriculus of other birds (Batah et al., 2012; Hamdi et al., 2013) in the same organ of Coot bird (Fulica atra) and most avian species, respectively. Tunica serosa examination revealed loose connective tissue rich in blood and covered by the mesothelium of simple squamous cells. The structure is commonly observed in many avian species, such as; Turkey (El-Zoghby, 2000).
Histochemical findings: Microscopic examination of the proventriculus revealed cells in its surface lining of the mucosal folds strongly positive to PAS as the reaction gave rise dark purple coloration. The observed reaction was with the granules located at the supra-nuclear area of these cells which was an indication of the presence of neutral type of mucin (Fig. 6a). These findings were comparable to those observed by Hamdi et al. (2013) in the glandular stomach of the black-winged kite (Elanus caeruleus). Similarly, in the proventriculus of the quail, Zaher et al. (2012) observed PAS-positive mucin granules occupying the supra-nuclear area of the cells in the surface lining epithelium of the mucosal folds. The connective tissue of the lamina propria, which extended between the folds of mucosa showed moderate reaction toward PAS in mallard. The submucosal glands in glandular stomach were strongly reacted with such stain. The muscularis mucosa and the walls of the blood vessels in the submucosa showed mild to moderate reactions. Whereas, the tunica muscularis showed mild reaction but interspersed collagen fibers showed a moderate reaction.
On applying the combined PAS-AB (pH 2.5) stain, the mucous cells lining the surface epithelium and gastric pits were strongly reacted giving rise to blue and magenta staining with it. In fact, such reaction indicated the presence of high content of neutral and acidic polysaccharides, respectively. Whereas, the glandular secretion gave the magenta color only (Fig. 6b). Recently, Zaher et al. (2012) established similarly in the proventriculus of the quail, abundant neutral and acidic mucopolysaccharides in the gastric glands since they gave red and blue colors with PAS-AB procedure stain.
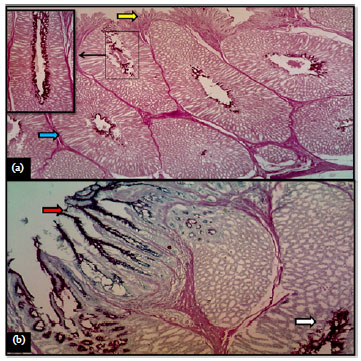 | |
| Fig. 6(a-b): | (a) Proventriculus wall of mallard showed mucosal layer (yellow arrow) and submucosal glands blue arrow), PAS, X 40, Rectangle, X 400 and (b) Proventriculus wall of mallard showed surface epithelia (red arrow) and submucosal, glands (white arrow), PAS-AB (pH 2.5), X 40 |
As same as to the above findings, neutral and acid mucopolysaccharides were detected in previous study in the surface lining of the mucosal folds in the proventriculus of the black-winged kite (Elanus caeruleus) too (Hamdi et al., 2013). Similarly, Selvan et al. (2008) recorded neutral and acid mucins in the mucosal surface of the proventriculus of the guinea fowl. In addition to that previous comments stated that the presence of neutral and acid mucins will aid to protects the mucosal surface and forms a resistant mucosal barrier (Hamdi et al., 2013).
The columnar epithelial cells were positively reacted with PAS-AB (pH 2.5) in the glandular part of the stomach (Fig. 6b). However, the lamina propria, muscularis mucosa, walls of the blood vessels in the submucosa and the tunica muscularis showed mild reaction. The mucosal glands were positively reacted to this combined stain (Fig. 6b). These findings coincided well with those observations in domestic fowl (Abumandour, 2014) and in the ducks (Shyla et al., 1992) in which they were reported PAS and Alcian blue positive mucin granules (neutral and acid mucin, respectively) in their proventriculus mucosa. Such results were in accordance also with recent findings of Selvan et al. (2008) whom recorded both acid and neutral mucins in the proventricular surface epithelial cells in the Guinea fowl (Numida meleagris).
The cuticle covering which was detected in the ventriculus of mallard showed positive reaction to PAS stain (pink color) as it present above their epithelial lining (Fig. 7). Cuticle positive reaction with this stain was similarly observed by Hamdi et al. (2013) in the ventriculus of the black-winged kite (Elanus caeruleus) and Selvan et al. (2008) in the Guinea fowl (Numida meleagris). The positive colored cuticle, which was recorded currently also observed in other birds such as the quail (Zaher et al., 2012) and in ducks and pigeons. The latter, described the cuticle layer as abrasion-resistant lining membrane present as a covering to the mucosa and extended deeply into the glandular lumina.
The epithelium, which lined the mucosal folds in the mucosal layer showed negative reaction with PAS. The secretory material within the lumina of the glandular tubules were negatively reacted with PAS stain (Fig. 7). Dissimilarly, in the black-winged kite (Elanus caeruleus) Hamdi et al. (2013) recorded strong positive reaction with this stain in the same glandular tubules of the ventriculus as the birds was classified as carnivorous avian species.
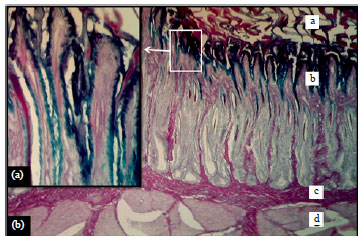 | |
| Fig. 7(a-b): | (a) Ventriculus wall of mallard showed a: Cuticle, b: Mucosa, c: Submucosa and d: Muscularis, X 40 and (b) magnification of mucosal epithelial cells, PAS-AB (pH 2.5), X 400 |
Dissimilar observations to those of mallard were previously recorded by Selvan et al. (2008) in the surface epithelium of ventriculus of the Guinea fowl (Numida meleagris) in which it revealed PAS positive materials. In addition to that, Aitken (1958) stated that the columnar cells covering the surface and the lining of the upper part of the mucosal tubular glands which contain granules were stained positively with the PAS in the ventriculus of chicken.
The connective tissue in the lamina propria, submucosa and in tunica muscularis showed PAS positive reaction in ventriculus, while the smooth muscles fibers which constructed the tunica muscularis of the organ gave rise mild reaction (Fig. 7).
When the combined PAS-AB (pH 2.5) used, the cuticle layer showed pink-colored positive reaction for PAS and negative with Alcian blue mallard ventriculus. The PAS-positive cuticle layer was similarly, observed in other birds such as Guinea fowl (Numida meleagris) (Selvan et al., 2008). The mucosal simple cuboidal epithelium lining of the surface and gastric pits of the ventriculus tubular glands were stained strongly positive with both parts of the combined PAS-AB (pH 2.5) stain (Fig. 7). It indicated the presence of both neutral and acidic mucin, respectively. Such observations were similar to those demonstrated by Pastor et al. (1988) and Imai et al. (1991) in the propria glandular cells in the ventriculus of chicken and fowls, respectively. The presence of neutral and acid mucin may protect the mucosal surface and forms a resistant mucosal barrier in the ventriculus of the birds (Hamdi et al., 2013). The latter reference found out in the ventriculus of black-winged kite which is considered one of the meat eater birds, similar positive reactions with this combined stain in the mucosa and the gastric crypts, ventriculus tubular glands and the secretory material within the lumina of these glands due to the presence of both neutral and acid mucin. The connective tissue gave the positive reaction with PAS and negative with AB. Additionally, the smooth muscle bundles of the tunica muscularis reacted weakly with PAS and negatively with AB (Fig. 7).
CONCLUSIONS
The stomach in the mallard appeared as an organ distinctly divided into two parts that were proventriculus and ventriculus, distinctly possessed well developed demarcation between them. The ventriculus appeared fusiform in shape and characteristically large in size. The cuticle present white in color in the ventriculus. The proventriculus was lined by simple columnar cells, whereas, the ventriculus was lined by simple cuboidal cells. The muscularis mucosa was very thick structure of smooth muscles fibers. The proventriculus reacted negative with the PAS in surface mucous cells and the mucous glands. The cuticle showed positive reactions to PAS stain. The connective tissue reacted positively with PAS and negative with the AB stain. In fact, the latter stain is an indicator for the acidic mucin substances.
REFERENCES
- Abumandour, M.M.A., 2014. Histomophological studies on the stomach of Eurasian hobby (Falconinae: Falco subbuteo, Linnaeus 1758) and its relation with its feeding habits. Life Sci. J., 11: 809-819.
Direct Link - Ahmed, Y.A.E.G., G. Kmel and A.A.E.M. Ahmad, 2011. Histomorphological studies on the stomach of the japanese quail. Asian J. Poult. Sci., 5: 56-67.
CrossRefDirect Link - Aitken, R.N.C., 1958. A histochemical study of the stomach and intestine of the chicken. J. Anat., 92: 453-466.
PubMedDirect Link - Akester, A.R., 1986. Structure of the glandular layer and koilin membrane in the gizzard of the adult domestic fowl (Gallus gallus domesticus). J. Anat., 147: 1-25.
PubMedDirect Link - Attia, H.F., 2008. Some histological studies on the proventriculus of the quail during pre and post hatching periods. Minufiya Vet. J., 5: 441-453.
Direct Link - Bailey, T.A., E.P. Mensah-Brown, J.H. Samour, J. Naldo, P. Lawrence and A. Garner, 1997. Comparative morphology of the alimentary tract and its glandular derivatives of captive bustards. J. Anat., 191: 387-398.
CrossRefDirect Link - Batah, A.L., H.A. Selman and M. Saddam, 2012. Histological study for stomach (proventriculus and gizzard) of coot bird Fulica atra. Diyala Agric. Sci. J., 4: 9-16.
Direct Link - Baumel, J.J., A.S. King, J.E. Breazile, H.E. Evans and J.C. Vanden Berge, 1993. Handbook of Avian Anatomy: Nomina Anatomica Avium. 2nd Edn., Nuttall Ornithological Club, Cambridge, MA., USA., ISBN-13: 9781877973345, Pages: 779.
Direct Link - Catroxo, M.H.B., M.A.I. Lima and C.E.M.P.D.M. Cappellaro, 1997. Histological aspects of the stomach (Proventriculus and gizzard) of the red-capped cardinal (Paroaria gularis gularis, Linnaeus, 1766). Revista Chilena Anatomia, 15: 19-27.
CrossRefDirect Link - Cooper, R.G. and K.M. Mahroze, 2004. Anatomy and physiology of the gastro-intestinal tract and growth curves of the ostrich (Struthio camelus). Anim. Sci. J., 75: 491-498.
CrossRefDirect Link - Duke, G.E., 1997. Gastrointestinal physiology and nutrition in wild birds. Proc. Nutr. Soc., 56: 1049-1056.
CrossRefPubMedDirect Link - Gionfriddo, J.P. and L.B. Best, 1996. Grit-use patterns in North American birds: The influence of diet, body size and gender. Wilson Bull., 108: 685-696.
Direct Link - Hamdi, H., A.W. El-Ghareeb, M. Zaher and F. AbuAmod, 2013. Anatomical, histological and histochemical adaptations of the avian alimentary canal to their food habits: II-Elanus caeruleus. Int. J. Scient. Eng. Res., 4: 1355-1364.
Direct Link - Imai, M., T. Shibata, K. Moriguchi, M. Yamamoto and H. Hayama, 1991. Proventricular glands in fowl. Okajimas Folia Anatomica Japonica, 68: 155-160.
CrossRefPubMedDirect Link - Kadhim, K.K., A.B.Z. Zuki, M.M. Noordin, S.M.A. Babjee and M. Zamri-Saad, 2011. Activities of amylase, trypsin and chymotrypsin of pancreas and small intestinal contents in the red jungle fowl and broiler breed. Afr. J. Biotechnol., 10: 108-115.
Direct Link - Nasrin, S., M.A. Islam, M. Khatun, L. Akhter and S. Sultana, 2012. Characterization of bacteria associated with omphalitis in chicks. Bangladesh Veterinarian, 29: 63-68.
CrossRefDirect Link - Pastor, L.M., J. Ballesta, J.F. Madrid, R. Perez-Tomas and F. Hernandez, 1988. A histochemical study of the mucins in the digestive tract of the chicken. Acta Histochemica, 83: 91-97.
CrossRefPubMedDirect Link - Rodrigues, M.N., J.A.P. Abreu, C. Tivane, P.G. Wanger and D.B. Campos et al., 2012. Microscopical Study of the Digestive Tract of the Blue and Yellow Macaws. In: Current Microscopy Contributions to Advances in Science and Technology, Mendez-Vilas, A. (Ed.). Formatex Research Center, Spain, ISBN-13: 9788493984359, pp: 414-421.
Direct Link - Rocha, S.D.O. and M.A.I. De Lima, 1998. Histological aspects of the stomach of burrowing owl (Speotyto cunicularia, Molina, 1782). Revista Chilena Anatomia, 16: 191-197.
CrossRefDirect Link - Rossi, J.R., S.M. Baraldi-Artoni, D. Oliveira, C. da Cruz, V.S. Franzo and A. Sagula, 2005. Morphology of glandular stomach (Ventriculus glandularis) and muscular stomach (Ventriculus muscularis) of the partrigde Rhynchotus rufescens. Ciecia Rural, 35: 1319-1324.
CrossRefDirect Link - Schweizer, M., M. Guntert, O. Seehausen, C. Leuenberger and S.T. Hertwig, 2014. Parallel adaptations to nectarivory in parrots, key innovations and the diversification of the Loriinae. Ecol. Evol., 4: 2867-2883.
CrossRefDirect Link - Selvan, P.S., S. Ushakumary and G. Ramesh, 2008. Studies on the histochemistry of the proventriculus and gizzard of post-hatch guinea fowl (Numida meleagris). Int. J. Poult. Sci., 7: 1112-1116.
CrossRefDirect Link - Tadjalli, M., P. Parto and A.F. Shahraki, 2011. Histological study of proventriculus of male adult ostrich. Global Veterinaria, 7: 108-112.
Direct Link - Zaher, M., A.W. EL-Ghareeb, H. Hamdi and F.A. Amod, 2012. Anatomical, histological and histochemical adaptations of the avian alimentary canal to their food habits: I-Coturnix coturnix. Life Sci. J., 9: 253-275.
Direct Link