
E.V. Ikpeme
Department of Genetics and Biotechnology, University of Calabar, PMB 1115, Calabar, Nigeria
O.U. Udensi
Department of Genetics and Biotechnology, University of Calabar, PMB 1115, Calabar, Nigeria
U.B. Ekaluo
Department of Genetics and Biotechnology, University of Calabar, PMB 1115, Calabar, Nigeria
M.E. Kooffreh
Department of Genetics and Biotechnology, University of Calabar, PMB 1115, Calabar, Nigeria
C.M. Okolo
Department of Genetics and Biotechnology, University of Calabar, PMB 1115, Calabar, Nigeria
P.B. Ekpo
Department of Genetics and Biotechnology, University of Calabar, PMB 1115, Calabar, Nigeria
N.C. Ogbonna
Department of Genetics and Biotechnology, University of Calabar, PMB 1115, Calabar, Nigeria
Asian Journal of Animal Sciences
Year: 2015 | Volume: 9 | Issue: 5 | Page No.: 187-197







ABSTRACT
The utilization of molecular markers-assisted genetic diversity study is critical and imperative for a more robust selection and breeding programmes in aquaculture. Twenty C. gariepinus species were randomly selected, 5 each from four Rivers/locations-Okurikan (OKU), Lemna (LMN), UNICAL Fish Farm (UFF) and MCC cultured fish site, in Cross River State, Nigeria for genetic diversity study using RAPD technique. Genomic DNA extraction and purification from caudal fin tissue sample was performed using ZR Genomic DNA tissue extraction Kit, primed using OPA09, OPA11 and OPA13 RAPD primers and amplified using PCR. The amplified fragments were ran on gel electrophoresis and visualized under ethidium bromide dye. Thirty four bands were generated by the three RAPD primers from the four C. gariepinus populations of which 28 were polymorphic, accounting for 82.35%. Polymorphism revealed by each fish population ranged from 92.86-100% while, gene diversity estimates ranged from 0.246±0.16-0.385±0.09. Intra-population similarity index ranged from 42.49-78.4% while, the inter-population similarity index across all population combinations ranged from 52.32-71.69%. These result also revealed that as genetic distances increases, genetic identities narrows between the wild and the cultured species. UPGMA dendrogram generated two major clusters with C. gariepinus species from OKU, UFF and MCC in cluster-1 and C. gariepinus species from LMN in cluster-2. Implicitly though, our results suggest that there were several genetic diversity in the populations studied, especially between the cultured and the wild, which could encourage selection of C. gariepinus from OKU and LMN populations for the purpose of breeding and improvement.
PDF Abstract XML References Citation
Received: May 06, 2015;
Accepted: June 10, 2015;
Published: July 02, 2015
How to cite this article
E.V. Ikpeme, O.U. Udensi, U.B. Ekaluo, M.E. Kooffreh, C.M. Okolo, P.B. Ekpo and N.C. Ogbonna, 2015. Unveiling the Genetic Diversity in Clarias gariepinus (Burchell, 1822) Using Random Amplified Polymorphic DNA (RAPD) Fingerprinting Technique. Asian Journal of Animal Sciences, 9: 187-197.
DOI: 10.3923/ajas.2015.187.197
URL: https://scialert.net/abstract/?doi=ajas.2015.187.197
DOI: 10.3923/ajas.2015.187.197
URL: https://scialert.net/abstract/?doi=ajas.2015.187.197
INTRODUCTION
Clarias gariepinus also known as African sharp-tooth catfish, is a typical air-breathing catfish with a scaleless, bony elongated body, long dorsal and anal fins as well as helmet like head. According to Picker and Griffiths (2011), C. gariepinus native range covers most African countries though with exception of Maghreb, Upper and Lower Guinea as well as Cape provinces of South Africa. However, though presumably, it is believed to be most widely distributed fish species in Africa (Skelton, 2001). Clarias gariepinus is the 2nd most important freshwater fish, after tilapia, in Africa. This is with exception of Nigeria, where C. gariepinus production exceeds that of tilapia, accounting for 70-80% of the total freshwater fish production. This notwithstanding, the industry in Africa is not well established due to several limiting factors, which could include, development of catfish production in relation to environmental protection, genetic diversity and the future sustainability of the industry. Despite these surmountable problems facing fish industry, Danish et al. (2012) observed that C. gariepinus cultivation has the potential towards livelihood development, employment generation as well as ensuring nutritional enrichment in the regular diet among the populace.
Undoubtedly, genetic variability in fishes has been reported to be very valuable for aquaculture and fisheries management, identification of stocks, selection for breeding programmes, ecology restoration and estimation of genetic contributions in stocks (Danish et al., 2012). In the word of Giri et al. (2012), germplasm characterization is a critical link between the conservation and efficient utilization of genetic resources. Simonsen et al. (2005) asserted that ecological erosion orchestrated by natural and human activities possesses a serious threat to fish biodiversity resulting to reduced population size with the implication of decreased genetic variation. This decreased in genetic variation could be blamed on genetic drift and inbreeding constraint. This informed (Danish et al., 2012) probably to report that genetic monitoring is very imperative for an effective management strategy as fish population can suffer severe genetic erosion such as genetic drift, bottleneck, inbreeding, founder effect, etc., without being detected by the traditional demographic monitoring approach.
Several marker technologies have been employed in fish diversity studies which include morphometric, isozymes, cytologic and recently molecular markers (DNA markers). The essence for the study was for stock analysis, taxonomy/systematic as well as selective breeding programmes (Waldman and Wirgin, 1993; Ward and Grewe, 1994; Carvalho and Hauser, 1994; Ferguson and Dangamam, 1998). Of the molecular markers used, Random Amplified Polymorphic DNA (RAPD) has been applied for the study of genetic diversity in several organisms including fish (Cagigas et al., 1999; Bartish et al., 2000; Lumaret et al., 2000; Mohd-Azmi et al., 2000; Hwang et al., 2001). This marker is a dominant marker and according to Theodorakis and Bickham (2004), it can sample a large number of loci and does not require the prior DNA sequence information to perform the assay.
Mickett et al. (2003) proposed that understanding the genetic variation within catfish population is a main requirement for maximizing the selective breeding of this species. The study becomes timely owing to the dire need of C. gariepinus farmers to stock superior genotypes on one hand and the need to conserve and domesticate C. gariepinus species, especially in Cross River State, Nigeria where it is the fastest growing aspect of aquaculture. Unfortunately, there is paucity of information on the genetic diversity of C. gariepinus, its importance notwithstanding. Unveiling the genetic diversity of C. gariepinus using RAPD technique would be helpful to this direction.
MATERIALS AND METHODS
Sample collection: A total of 20 fish samples weighing approximately 1 kg were obtained from different cultured and wild populations of C. gariepinus in Calabar for diversity assessment. Locations for the wild populations include Lemna River (LMN) and Okurikan Beach (OKU) while, the cultured populations are Molecular Laboratory, 127 MCC Rd. Calabar (MCC) and Unical Fish Farm (UFF). Five fish samples of C. gariepinus were sourced from each of these sample locations. DNA analysis was done at Nigerian Institute of Medical Research Laboratory (NIMR) Yaba, Lagos, Nigeria.
Molecular analysis using RAPD
Isolation of genomic DNA: Fin samples were clipped from the caudal fin of each catfish and preserved in 95% ethanol for further analysis. Genomic DNA extraction and purification from fin tissue sample was performed using ZR Genomic DNA tissue extraction Kit (Zymo Research, California, USA) according to the manufactures protocols for solid tissue. Approximately 25 mg of each fin tissue was cut into small sizes, macerated and put into separate eppendorf tubes. The 95 μL of water, 95 μL of digestion buffer and 10 μL of proteinase K was added, respectively mixed and incubated at 55°C for 1 h. Genomic lyses buffer of 700 μL was added to the tube, vortexed and centrifuged at 10,000×g for 1 min to remove insoluble debris. The supernatant was transferred to Zymo-spin column in a collection tube and centrifuged for another minute. The DNA pre-wash buffer of 200 μL was added to the spin column in a new collection tube followed by 400 μL of g-DNA wash buffer and centrifuged. Finally, 100 μL DNA elution buffer was added, incubated for 2 min and centrifuged to elute the DNA which was stored at 20°C for further analysis. The concentration of DNA was estimated using a spectrophotometer.
Primer screening and selection: Five decamer primers of random sequence were procured from Operon technologies, Alameda, USA. The primers were tested on randomly selected fish from each population in order to select primers of high intensity or resolution and repeatability of bands and polymorphism. Finally, three primers OPA09, OPA11 and OPA13 that exhibited comparatively high quality banding pattern and sufficient variability were used for analysis of all the samples (Table 1).
Polymerase Chain Reaction (PCR) amplification: Polymerase chain reaction was performed according to the method of Islam et al. (2012) with modifications. The DNA amplifications were carried out by using Ready to use AccuPower PCR PreMix bead (Bioneer Co., USA) containing 250 μL each dNTPs (dATP, dGTP, dCTP, dTTP), 10 mM Tris-HCl (pH 9.0), 30 mM KCl, 1.5 mM MgCl2, stabilizer and tracking dye. The 16 μL of sterilized ultra pure water, 1 μL of primer and 3 μL of DNA were added to the premix. The reaction mixture was amplified using thermal cycler (A and E Laboratories UK) under the following conditions, initial denaturation at 94°C for 2 min, followed by denaturation at 94°C for 1 min, annealing at 40°C for 1 min, extension at 72°C for 2 min for 40 cycles and final extension at 72°C for 5 min.
Electrophoresis of amplified products: The amplified products (10 μL) of each sample were separated by electrophoresis on 1.5% agarose gel containing 0.5 μg mL-1 ethidium bromide in TAE buffer (40 mM Tris-acetate, 2 mM EDTA [pH 8.3]) and run at 75 V for 2.5 h. A DNA ladder digest of 100 bp (Bioneer, South Korea) was electrophoresed alongside the RAPD reactions as molecular weight marker. Finally, the DNA bands were examined under Ultra-Violet (UV) transilluminator system (Clinix, Japan), image photographed and saved on computer. The RAPD patterns were analyzed and scored from the photomicrographs of the gel.
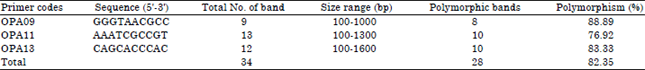
| Table 1: | Primer used to generate RAPD fingerprint with polymorphic bands obtained per primer from Clarias gariepinus DNA |
 | |
Statistical analysis: For the analysis and comparison of the patterns, only the well-separated clearly visible bands were scored. Band scoring from gel and band size calculation was done using the software Pyelph 1.3. The RAPD markers were scored by recording the presence (1) or absence (0) of these bands for each fish and each primer. A single data matrix was constructed pooling the binary data (1/0) for all the primers and used for estimating polymorphic loci, gene diversity, observed number of alleles (Na), effective number of alleles (Ne), gene flow (Nm) and genetic distance with the POPGENE (Version 1.31) (Yeh et al., 1999) computer program. The computer program was used to construct an Un-weighted Pair Group Method of Arithmetic Average (UPGMA) dendrogram among populations. The Similarity Index (SI) between the RAPD fingerprint of any two individuals on the same gel was calculated from RAPD band sharing according to the following equation:
![]()
where, NAB is the total number of RAPD bands shared by individuals A and B and Na and NB are the total number of bands produced by individuals A and B, respectively (Islam et al., 2012). Within population, similarity (Si) will be calculated as the average of S1 across all possible comparisons between individuals within a population. Between populations, similarity (Sij) was calculated as the average between randomly paired individuals from population i and j (Islam et al., 2012).
RESULTS
RAPD analysis
Rapid profiles and polymorphism generated by PCR: Among the 5 primers screened, OPA09, OPA11 and OPA13 yielded comparatively larger number of bands with good resolution though, OPA 13 (Fig. 1) showed more discriminating bands than OPA 09 and OPA 11. Each primer produced a unique fragment pattern of amplified DNA with varied number of bands. From Table 1, OPA09 though with the least number of bands resulted to 88.89% polymorphism, when compared with 83.33 and 76.92% of OPA11 and OPA13, respectively.
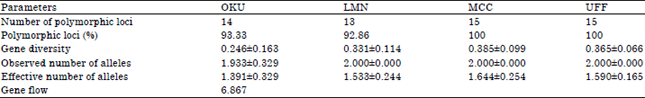
Genetic variation in the populations: The proportion of polymorphic loci was highest in the C. gariepinus species obtained from MCC and UFF with 100% polymorphism each. Clarias gariepinus species from LMN had the lowest polymorphism of 92.86% while, the C. gariepinus species from OKU had 93.33% (Table 2). The gene diversity of C. gariepinus species from MCC was highest (0.385±0.099) while, those of OKU (0.246±0.163) had the least. The effective number of alleles in the populations of C. gariepinus from OKU, LMN, MCC and UFF were 1.391±0.329, 1.533±0.244, 1.664±0.254 and 1.590±0.165, respectively. It was observed that the gene flow among the populations investigated was 6.867 (Table 2).
Intra and inter population similarity indices: For intra-population similarity indices, UFF C. gariepinus population had the highest similarity index of 78.4%, which was followed by the C. gariepinus population obtained from MCC. However, C. gariepinus population with the lowest similarity index was OKU (42.49%) (Fig. 2).
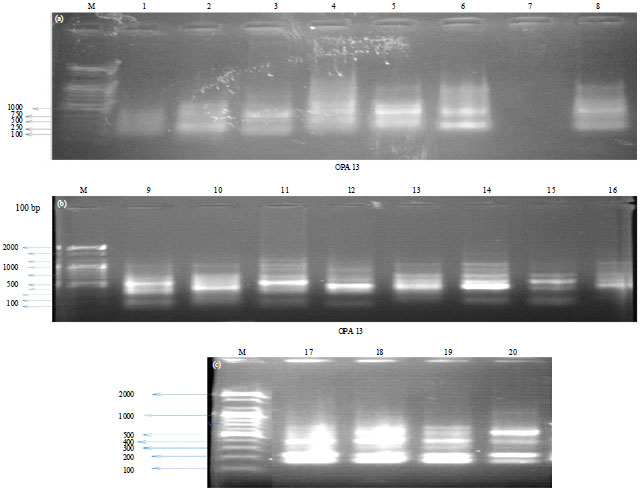 | |
| Fig. 1(a-c): | RAPD profile of Clarias gariepinus for primer OPA13. M: 100 bp DNA ladder |
| Table 2: | Estimates of genetic variation: number and proportion of polymorphic loci, gene diversity, number of alleles, effective number of allele and gene flow |
 | |
| Table 3: | Inter-population similarity indices of the Clarias gariepinus populations |
 | |
Clarias gariepinus population obtained from UFF and MCC has the highest inter-population similarity index of 71.69%, which was followed by the values obtained between Clarias population from LMN and UFF while, fish populations from OKU and LMN had the least similarity index of 52.32% (Table 3).
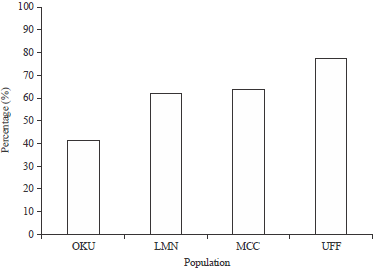 | |
| Fig. 2: | Intra-population similarity indices of the Clarias gariepinus populations sampled. OKU (42.49%), LMN (62.14%), MCC (64.97%) and UFF (78.4%) |
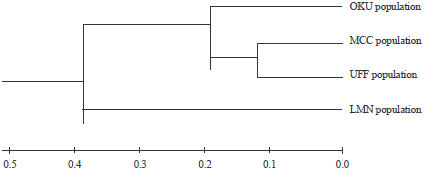 | |
| Fig. 3: | Unweighted pair-group method with average dendrograms based on Nei’s D value (Nei, 1972), original measures of genetic distance, summarizing the data on differentiation between Clarias gariepinus populations according to RAPD analysis |
| Table 4: | Genetic identity (above diagonal) and genetic distance (below diagonal) among the four populations of Clarias gariepinus (Nei’s original measure of genetic identity and genetic distance) |
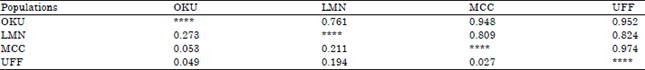 | |
Genetic identity and genetic distance: The value for inter-population genetic identity was highest for MCC and UFF populations (0.974) and Lowest for OKU and LMN populations (0.761). The genetic identity values 0.948, 0.92, 0.809 and 0.824 were found between OKU and LMN, OKU and UFF, LMN and MCC, LMN and UFF, respectively. The highest and lowest genetic distances were found between the OKU and LMN (0.273) and between the MCC and UFF (0.027) C. gariepinus populations, respectively (Table 4). The UPGMA dendrogram based on Nei’s genetic distance indicated the segregation of the four C. gariepinus populations sampled into two distinct clusters. OKU, MCC and UFF are in cluster 1 and LMN in cluster 2. The cultured populations of MCC and UFF produced one sub cluster (Fig. 3). However, when individual fish species were clustered, results showed several over lappings inter and intra-population wise (Fig. 4).
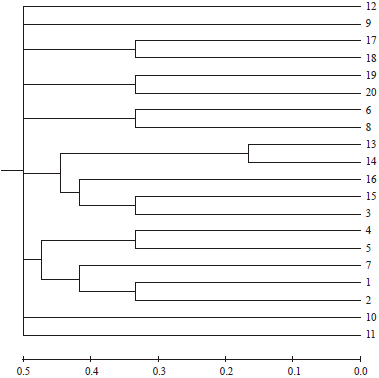 | |
| Fig. 4: | Unweighted pair group method with average dendrogram based on Nei’s (Nei, 1972) genetic distance, summarizing data on Clarias gariepinus individuals according to RAPD analysis. Individual (1-5 from OKU, 6-10 from LMN, 11-15 from MCC and 16-20 from UFF) |
DISCUSSION
One of the devastating challenges in the Sub-Saharan African countries (SSA) is poverty amidst scourging malnutrition. Ours is a continent, where refugee camps are on the increase with no hope of providing them with basic needs of life, including balanced diet. Danish et al. (2012) stated that while, the farming/cultivation of C. gariepinus has the potential towards livelihood development, employment generation; most importantly nutritional enrichment (protein) in the regular diet among populace, there is a remarkable reduction in the size of fish population due to environmental events-earthquakes, flood, drought or human activities (pollution). Additionally, inbreeding is a common problem in fish hatcheries. The former and the later are contributing factors to loss of genetic diversity in wild and cultured fish populations (Simonsen et al., 2005). Germplasm characterization is an important link between the conservation and efficient utilization of genetic resources (Giri et al., 2012; Mickett et al., 2003).
The RAPD markers were reported to be highly reproducible in a size ranging between 200 and 1500 bp (Liu et al., 1999). Good quality and reproducible PCR products of 100-1600 bp were found in our C. gariepinus study, which is similar to those found in Clarias batrachus (100-1200 bp) and of the same species (172-1677 bp) by Garg et al. (2010) in India. Present result showed that out of the 5 RAPD primers screened, OPA09, OPA11 and OPA13 revealed high polymorphism. OPA09 gave 88.89% polymorphism though with the least polymorphic bands (8) while, OPA11 and OPA13 produced 76.92 and 83.33% polymorphism with 10 polymorphic bands each (Table 1). Islam et al. (2012) observed that the number as well as the sizes of the bands generated, depend upon the nucleotide sequence of the primers used and the source of the DNA template (Clark and Lanigan, 1993).
Additionally, percentage polymorphism as revealed by RAPD is detected as presence and absence of bands and may cause either by failure to prime a site in some individuals. The implication is that the differential among the primers used could have arisen due to the nucleotide sequence differences since the quality of the DNA template sources were good. Saad et al. (2009) and Popoola et al. (2014) reported lower percentage polymorphic loci of 68.5 and 74.7%, respectively in cultured C. gariepinus species, which did not corroborate with our current report of 100% for the cultured C. gariepinus population. However, our report agrees with the observation of Agbebi et al. (2013). This difference in polymorphic loci might be multifaceted which could include variant types of RAPD markers used.
Gene diversity is a measure of the expected heterozygosity in a sample of gene copies collected at a single locus. It represents patterns of molecular diversity within a sample of gene copies. It was observed that the gene diversity in the cultured populations [MCC (0.385); UFF (0.365)] were higher than in the wild populations [LMN (0.331); OKU (0.246)]. The implication is that the C. gariepinus from the cultured populations might be showing variant pattern of molecular diversity that may be absent in those from the wild. Band sharing Similarity Index (SI) is usually employed to compare the similarity in DNA fingerprints of two individual samples as to make deductions concerning the extent of genetic variation within and between populations. The similarity index gives an indication of relative levels of population homogeneity. It does suggest that populations showing higher intra-population similarity are likely to have less genetic diversity. For instance, a domestic population may suffer from a bottleneck, when only a few mating pairs reproduce hundreds of offspring that constitute the entire subsequent generation (Danish et al., 2012). This is very peculiar to hatchery owners that sometimes maintain a polygamous mating system in which a few males monopolize breeding and mate with many females. This could have led to the high intra population similarity indices observed in the C. gariepinus in the cultured populations. The inter population similarity value observed in C. gariepinus obtained from OKU indicates a good level of genetic diversity.
Dendrogram generated after RAPD analysis revealed two important clusters. Cluster-1 is made of C. gariepinus populations from OKU, MCC and UFF while, cluster-2 is made up C. gariepinus collected from LMN. The inference therefore, is that C. gariepinus sampled from Lemna River (wild) showed wide genetic diversity when compared with C. gariepinus population from Odukpani river though wild. However, in cluster-1, C. gariepinus population from Odukpani river also exhibited slight differences from those from the cultured populations (MCC and UFF). It should be understood that genetic diversity in any population is the level of biodiversity which represents the total number of genetic characteristics in the genetic makeup of a species. What is means is that C. gariepinus from the wild (LMN and OKU) may have higher genetic features comparing with those in the cultured populations (MCC and UFF). When the genetic diversity in a population is high, there will be a higher adaptability to changing environmental conditions by the species. According to Asuqwo and Udoh (2002), the pollution levels in Calabar River are alarming which Ewa et al. (2013) observed to be as a result of agricultural wastes, crude oil exploration and exploitation (Lae et al., 2004). This has led to high fish mortalities causing a drastic decline genetic diversity and variability (Nwafili et al., 2012; Islam et al., 2012) as well as risk of extinction (Hilborn et al., 2003). The high genetic diversity in the wild populations may have contributed significantly to their adaptability in the Rivers. The clustering of C. gariepinus species into major populations notwithstanding, inter and intra-population similarity indices showed that these similarities though genetic, might have been expressed morphologically, nutritionally as well as biochemically.
There seem to be an inverse relationship between genetic identity and genetic distance. Present result showed that C. gariepinus species obtained from OKU and LMN (wild) exhibited a genetic identity of 0.761 while, they were distant apart from each by 0.273. The implication explicitly affirms to the fact that as the genetic distance between one species and another widens, their genetic identity narrows giving a good opportunity for selection to be made in the population with wide genetic distance. The result also suggests that though C. gariepinus from OKU and LMN are wild species, there were so separated apart genetically as revealed by RAPD analysis. Since, RAPD analysis is devoid of the influence of the environment, it might probably be that C. gariepinus from these two rivers contain some superior economic important genes that may be absent in the other and present in another. This was true for C. gariepinus from MCC and UFF (cultured) with 0.027 as the genetic distance and 0.974 as genetic identity. Popoola et al. (2014) reported a high genetic identity between two cultured populations of C. gariepinus in the southwest, Nigeria.
Interestingly, it was observed that C. gariepinus sampled from OKU population had the least similarity index (42.49%), which was followed by species from LMN (62.14%). This genetically diverse nature of C. gariepinus from these populations could be attributed to the reduced inbreeding, bottleneck as well as founder effect experiences. This was not the case with C. gariepinus species from MCC (64.97%) snd UFF (78.40%). It is worthy of note that fish farmers do not breed rather the multiply for income generation. This activity may probably reduce the quality of fish in our markets, especially as it regards nutrition. Inter-population similarity indices revealed a thrilling trend. Clarias gariepinus species sampled from OKU and LMN showed low value (52.32%) which indicates more genetic diversity. However, C. gariepinus from MCC and UFF show a similarity index of 71.69%. When compared with that between OKU and LMN, C. gariepinus from MCC and UFF showed more genetic homogeneity. This is clearly implicated in the population dendrogram (Fig. 2). Clarias gariepinus obtained from OKU had the lowest percentage similarity index (42.49%) suggesting that C. gariepinus species from OKU River might carry important genes which could be exploited in hybridization, especially with C. gariepinus from LMN River for improvement purposes.
CONCLUSION
Selection of superior genotypes of C. gariepinus for breeding and improvement purposes is very cardinal revolutionizing the sector, which a key aspect of aquaculture in Calabar, Nigeria. Present results showed that there is wide genetic diversity in C. gariepinus comparing the wild and the cultured, thus encouraging selection of stocks from the cultured.
ACKNOWLEDGMENTS
This study was under the financial assistance of the Research Unit of the Department of Genetics and Biotechnology, University of Calabar, Nigeria. We want to sincerely thank the administration of the Nigerian Institute of Medical Research (NIMR) Yaba, Lagos, Nigeria for providing the laboratory space for this study. We also thank to Dr. Francesca Nwokorie for her technical assistance.
REFERENCES
- Agbebi, O.T., D.E. Ilaboya and A.O. Adebambo, 2013. Preliminary characterization of genetic strains in clariid species, Clarias gariepinus and Heterobranchus bidorsalis using microsatellite markers. Afr. J. Biotechnol., 12: 364-369.
CrossRefDirect Link - Asuqwo, F.E. and J.P. Udoh, 2002. Patterns of total hydrocarbon, copper and iron in some fish from Cross River Estuary, Nigeria. West Afr. J. Applied Ecol., 3: 91-97.
CrossRefDirect Link - Bartish, I.V., L.P. Garkava, K. Rumpunen and H. Nybom, 2000. Phylogenetic relationships and differentiation among and within populations of Chaenomeles Lindl. (Rosaceae) estimated with RAPDs and isozymes. Theor. Applied Genet., 101: 554-563.
CrossRefDirect Link - Cagigas, M.E., E. Vazquez, G. Blanco and J.A. Sanchez, 1999. Combined assessment of genetic variability in populations of brown trout (Salmo trutta L.) based on allozymes, microsatellites and RAPD markers. Mar. biotechnol., 1: 286-296.
CrossRefDirect Link - Carvalho, G.R. and L. Hauser, 1994. Molecular genetics and the stock concept in fisheries. Rev. Fish. Biol. Fish., 4: 326-350.
CrossRefDirect Link - Theodorakis, C.W. and J.W. Bickham, 2004. Molecular characterization of contaminant-indicative RAPD markers. Ecotoxicology, 13: 303-309.
CrossRefDirect Link - Clark, A.G. and C.M. Lanigan, 1993. Prospects for estimating nucleotide divergence with RAPDs. Mol. Biol. Evol., 10: 1096-1111.
Direct Link - Danish, M., I.J. Singh, P. Giri and C.P. Singh, 2012. Molecular characterization of two populations of catfish Clarias batrachus L. using random amplified polymorphic DNA (RAPD) markers. Afr. J. Biotechnol., 11: 14217-14226.
Direct Link - Ewa, E.E., A.I. Iwara, J.A. Adeyemi and G.N. Njar, 2013. Assessment of water quality of the Calabar River using multivariate statistical techniques. J. Applied Sci. Res., 9: 3354-3363.
Direct Link - Ferguson, M.M. and R.G. Danzmann, 1998. Role of genetic markers in fisheries and aquaculture: Useful tools or stamp collecting? Can. J. Fish. Aquat. Sci., 55: 1553-1563.
Direct Link - Garg, R.K., P. Sairkar, N. Silawat, N. Vijay, N. Batav and N.N. Mehrotra, 2010. Assessment of genetic diversity of Clarias batrachus using RAPD markers in three water bodies of Bhopal. J. Environ. Biol., 31: 749-753.
Direct Link - Giri, P., G. Taj and H.S. Ginwal, 2012. Molecular characterization of six populations of Acorus calamus L. using randomly amplified polymorphic DNA (RAPD) markers. Afr. J. Biotechnol., 11: 9522-9526.
Direct Link - Islam, M.N., A. Basak and M.S. Alam, 2012. Genetic diversity in wild and hatchery populations of stinging catfish (Heteropneustes fossilis Bloch) revealed by RAPD analysis. J. Bio-Sci., 19: 81-87.
Direct Link - Lae, R., S. William and A.M. Masosou, 2004. Review of the Present State of the Environment: Fish Stocks and Fisheries of River Niger, West Africa. In: Proceedings of the Second International Symposium on the Management of Large Rivers for Fisheries: Sustaining Livelihoods and Biodiversity in the New Millennium, Welcome, R.L. and T. Petr (Eds.). Vol. 1, Food and Agriculture Organization of the United Nations, Bangkok, Thailand, pp: 199-227.
- Lumaret, R., M. Amane, N. Ouazzani, L. Baldoni and C. Debain, 2000. Chloroplast DNA variation in the cultivated and wild olive taxa of the genus Olea L. Theo. Applied Gen., 101: 101-553.
CrossRefDirect Link - Mickett, K., C. Morton, J. Feng, P. Li and M. Simmons et al., 2003. Assessing genetic diversity of domestic populations of channel catfish (Ictalurus punctatus) in Alabama using AFLP markers. Aquaculture, Aquaculture,: 91-105.
CrossRefDirect Link - Mohd-Azmi, M.L., A.S. Ali and W.K. Kheng, 2000. DNA fingerprinting of red Jungle Fowl, Village chicken and broilers. Asian-Aust. J. Anim. Sci., 13: 1040-1043.
CrossRefDirect Link - Nwafili, S.A., O.O. Soyinka and T.X. Gao, 2012. Levels and patterns of genetic diversity in wild Chrysichthys nigrodigitatusin Lagos lagoon complex. Afr. J. Biotechnol., 11: 15748-15754.
CrossRefDirect Link - Saad, Y.M., M.S. Hanafi, M.A. Essa, A.A. Guerges and S.F. Ali, 2009. Genetic signatures of some Egyptian Clarias gariepinus populations. Global Veterinaria, 3: 503-508.
Direct Link - Simonsen, V., M.M. Hansen, K.L.D. Mensberg, R.I. Sarder and S. Alam, 2005. Widespread hybridization among species of Indian major carps in hatcheries, but not in the wild. J. Fish Biol., 67: 794-808.
CrossRef - Ward, R.D. and P.M. Grewe, 1994. Appraisal of molecular genetic techniques in fisheries. Rev. Fish Biol. Fish., 4: 300-325.
CrossRefDirect Link