
M.K. Pazir
Department of Shrimp Diseases, Iranian Shrimp Research Center, Bushehr, Iran
M. Afsharnasab
Department of Aquatic Diseases, Iranian Fisheries Research Organization, Tehran, Iran
N. Niamaymandi
Department of Shrimp Diseases, Iranian Shrimp Research Center, Bushehr, Iran
H. Khadem
Department of Aquatic Diseases, Iranian Fisheries Research Organization, Tehran, Iran
E. Akbarpour
Department of Fishery, Faculty of Engineering, Islamic Azad University Bushehr Branch, Bushehr, Iran
A.A. Zendebudi
Department of Shrimp Diseases, Iranian Shrimp Research Center, Bushehr, Iran
Asian Journal of Animal Sciences
Year: 2012 | Volume: 6 | Issue: 5 | Page No.: 209-219







ABSTRACT
This study was conducted to identify White Spot Syndrome Virus (WSSV) and Infections Hypodermal and Hematopoietic Necrosis Virus (IHHNV) by using clinical and histopathological sings of cultured shrimp, Litopenaeus vannamei in Bushehr farms, from December 2009 to April 2010. Samples were collected from Bushehr shrimp farms and hatcheries. Based on histopathological and gross signs, two viral diseases, WSSV and IHHNV have been detected. The gross signs of WSSV in the moribund samples showed reduced feeding, lethargy, difference in size, reddish coloration of appendages and white plaque on the carapace, while the gross sign of IHHNV exhibited abdominal dorsal changes of the muscles, opaque or milky spot on the cuticles, rostrum deformity syndrome and abdominal segmental abnormalities. The histopathological observations of WSSV showed basophilic Cowdry type A inclusion bodies in all tissues such as gills, haematopoietic tissue, cuticle epithelium, lymphoid organ and connective tissue. However, histologically, the hepatopancreas tissue showed vacuolization of B cells, without inclusion bodies. The histopathological signs observed such as hypertrophy, cellular degeneration and eosinophilic Cowdry type A inclusion bodies in the cells nucleus hepatopancreas, gills, haematopoietic, cuticle epithelium, digestive epithelium, lymphoid organ and connective tissue. Also, histopathological change of the striated muscles of the affected abdominal segmental abnormality showed severe Zenker's necrosis.
PDF Abstract XML References Citation
Received: November 05, 2011;
Accepted: May 03, 2012;
Published: August 17, 2012
How to cite this article
M.K. Pazir, M. Afsharnasab, N. Niamaymandi, H. Khadem, E. Akbarpour and A.A. Zendebudi, 2012. Histopathological Observation of White Spot Syndrome Virus and Infectious Hypodermal and Hematopoietic Necrosis Virus in Shrimp Farms, Litopenaeus vannamei, in Bushehr Province, Iran. Asian Journal of Animal Sciences, 6: 209-219.
DOI: 10.3923/ajas.2012.209.219
URL: https://scialert.net/abstract/?doi=ajas.2012.209.219
DOI: 10.3923/ajas.2012.209.219
URL: https://scialert.net/abstract/?doi=ajas.2012.209.219
INTRODUCTION
Shrimp farms have been expanded rapidly over the last two decades. In 2008, shrimp farming production in the world was reported 3, 281, 253 metric tons and the most commonly cultured species is Pacific white shrimp (Litopenaeus vannamei) that included 90% of global cultured shrimp (Afsharnasab et al., 2009).
Over the last decades shrimp pathogens (especially virus) have been expanded throughout shrimp aquaculture industries (Lightner, 1999). It was made a significant economic loses for the shrimp farming industry. So that, annually viral diseases caused millions dollar lost for the shrimp global industry (Wyban et al., 1992; Lightner, 1999). Baculovirus penaei was the first virus that has been reported from Gulf of Mexico in Farfantepenaeus duorarum (Couch, 1974). Although, more than 20 viruses have been identified in Penaeidae shrimps (Lightner, 1999) but four viruses consisted of WSSV, Yellow Head Virus (YHV), Taura Syndrome Virus (TSV) and IHHNV have the major impacts on the shrimp aquaculture (Lightner, 1996; Flegel, 1997).
Among the four viruses; WSSV is the major pathogenic infectious virus in Penaeidae family (Lightner, 1996). For the first time, this virus was reported in Marsupenaeus japonicus in Taiwan (Chen, 1995). It was also observed in other species, especially in Penaeus monodon in the most of Asia countries and the United States (Lo et al., 1999). In Iran the virus was identified in Fenneropenaeus indicus from Khozestan farms, that in some years created about 90% mortality within 3-10 days (Afsharnasab et al., 2007; Afsharnasab et al., 2009).
IHHNV is a parvovirus, which is the smallest (22 nm) among the mentioned viruses (Bonami et al., 1990). The first record of IHHNV was reported in Litopenaeus stylirostris from Hawaii farms in 1981 (Lightner et al., 1983a, b; Lightner, 1996; Flegel, 1997). This virus has been found in other areas through asymptomatic carriers (Motte et al., 2003). IHHNV was routinely found in wild and captured L. vannamei without any clinical signs (Mari et al., 1993). L. vannamei has partially resistant to the virus although lesions on the body and deformities such as Rostrum Deformity Syndrome (RDS) have been observed (Lightner, 1996). When RDS prevalence reached more than 30%, a great loss of 10 to 50% in marketing value has occurred (Wyban et al., 1992). Both WSSV and IHHNV have significant impact on shrimp production. WSSV is observed in many shrimp species and crabs, from various geographical areas and accompanying with 100% mortality (Rodriguez et al., 2003) that leading to a devastating economic impacts (Lo et al., 1999). IHHNV also is one of the most serious viral diseases of farmed penaeid shrimps (Lightner, 2003). It is widely distributed in many countries and has a large range of hosts in many species of cultured penaeid shrimps.
Different methods are used for identifying viral diseases including clinical signs, molecular methods, histopathology and Transmission Electron Microscopy (TEM). In this study WSSV and IHHNV were identified by using clinical signs and histopathology observation in L. vannamei farms of Bushehr province.
MATERIALS AND METHODS
The present study was performed in the six grow-out shrimp farms and five hatcheries which located in the Bushehr province, with previous history of WSSV mortality in contrary to IHHNV. Sampling was performed from December 2009 to April 2010. The 150 shrimp samples (juvenile and sub adult) accompanying with slow growth collected from grow-out shrimp farms along the Bushehr coastal area, Persian Gulf (Bandar Rig, Heleh, Shif, Mond and Delvar (I and II) sites) (Fig. 1). In addition, 200 shrimp larvae and post larvae producing from domesticated origin which were randomly selected from hatcheries. Moribund domesticated broodstock samples of L. vannamei (average weight 40-45 g) with opaque, whitish abdominal muscles and white, milky spot on carapace body cuticle were obtained from private sectors (87 samples). Individual shrimp were kept on ice and examined grossly for signs of WSSV, IHHNV and other viral diseases. The sub adults and broodstock shrimps were injected with cold Davidson's fixative solution (4°C) into the hepatopancreas, gills and 2, 4 and 6 abdominal segmental tissues.
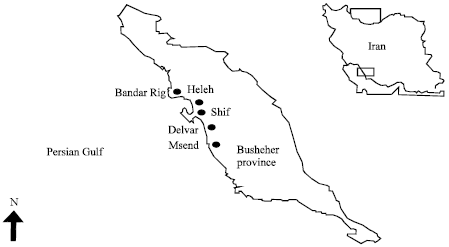 | |
| Fig. 1: | Location of shrimp farms in Bushehr province |
All the samples for histopathological examination were immersed with the same fixative for 24-48 h (ratio 1:10). The fixative was discarded and the samples preserved in 50% ethyl alcohol for subsequent histopathological preparation and analysis ( Bell and Lightner, 1988). Tissues (hepatopancreas, gills, lymphoid organ, connective tissue, hematopoietic, cuticular epidermis, digestive epithelium and striated muscles) were sectioned at 4-5 μm in thickness and stained with hematoxylin and eosin phloxine (H and E/ph) (Saberi et al., 2008; Afsharnasab et al., 2009; Fouzi et al., 2012). Subsequently gross signs and histopathological studies of WSSV and IHHNV were evaluated using light microscope (CETI; Triton II).
RESULTS
The gross signs of WSSV infections shrimps showed a reduced feeding, lethargy, difference in size, gathering around the ponds, reddish discoloration of body and appendages (i.e., the antenna, maxillipeds, pereiopods, pleopods, telson and uropods) with presence of white plaque on carapace cephalothorax or diffuse over body cuticle as characteristic feature of WSSV disease (Fig. 2a, b, g and h). These spots were abnormal deposits of calcium salts in cuticle. WSSV infection of some samples was confirmed by histopathological examination. Histopathological changes of hepatopancreas tissues such as vacuolization of B cells and increasing in the number of F and R cells, without intranuclear inclusion bodies Cowdry type A were observed (Fig. 3a). The epithelial pillar cells of secondary lamella of gill tissues showed basophilic intranuclear inclusion bodies Cowdry type A. In addition to fusion formations made between secondary lamella of infect tissue and the cells which exhibited the nucleus hypertrophy and watery and thin cytoplasm (Fig. 3b). Lymphoid organ showed three stages of WSSV infection. At the first stage, the nucleus of cells was hypertrophy and cytoplasm was thin. In the mediate stage, marginated chromatin and the space between nucleus and cell membrane was very thin. In the late stage, the final shape of intranuclear inclusion bodies was observed as similar spheroid cells (Fig. 3c). On the other hand, the number of hyaline cells of hematopoietic tissues in infectious broodstocks obtained from hatcheries accompanying with nucleus pyknosis and karyorrhexis was recorded higher compared to hematopoietic tissues of normal shrimps (Fig. 3d).
 | |
| Fig. 2(a-h): | Clinical sings in infection shrimps to WSSV and IHHNV, (a) White spots on carapace shrimps, (b) Reddish coloration of appendages (Telson area), (c) Rostrum deformity syndrome (RDS), (d) Milky moles on abdominal segmental, (e) Abdominal abnormalities, (f) Abdominal dorsal muscles opaque, (g) Accumulation of dead shrimps in the pool margin and (h) Difference in size |
In the prepared sections with external layers, the epithelial cells of cuticle in infected shrimps, which showed hypertrophy and basophilic intranuclear inclusion bodies, were detached from epidermis (Fig. 3e). In these specimens, characteristic WSSV basophilic intranuclear inclusion bodies and nucleus pyknosis of the cells were abundant in connective tissues of the general body; cuticle, appendages and hepatopancreas (Fig. 3f). Histopathological signs of striated muscles indicated hypertrophy and dense nucleus which were higher in infected broodstocks in compare with post larvae (Fig. 3g). The hypertrophied columnar cells and intranuclear inclusion bodies were observed in mid gut, where some same cells were ruptured and released inclusion bodies to the lumen of gut (Fig. 3h). Significantly, all these samples were showed WSSV infectious in histopathological study.
Clinical signs of IHHNV infected shrimps such as opaque, whitish abdominal muscles and white, milky moles on body cuticle (abdominal segmental junction) and shrimp with different size, slow growth and Rostrum Deformity Syndrome (RDS) were observed in samples. In addition cuticular deformities were found in the third to sixth abdominal segments and tail fan in sub adults grow-out shrimp farms and broodstocks hatcheries (Fig. 2c, d, e, f and h). Histopathological changes of the post larvae, sub adults and broodstocks infected tissues, especially in the hepatopancreas, cuticular epithelium, gills, connective tissues, hematopoietic tissues and digestive epithelium were characterized by widespread cellular degeneration, severe nuclear hypertrophy and margination chromatin (Fig. 4a-g).
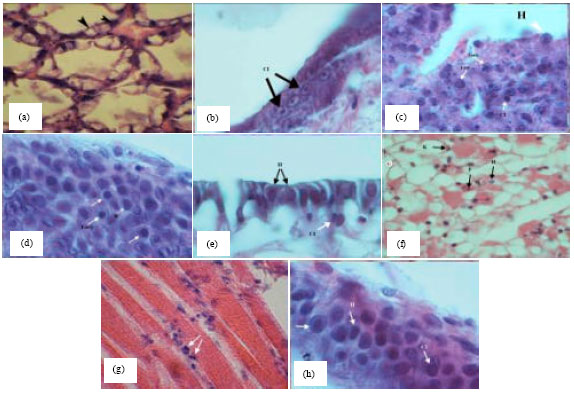 | |
| Fig. 3(a-h): | Histopathological observations of WSSV infection in tissue shrimps include nucleus hypertrophy (H), cellular degeneration (CD), intranuclear inclusion bodies Cowdry type A (CI) and nucleus pyknosis (P) and karyorrhexis (K) in L. vannamei (H and E\Ph 1000X), (a) Vacuolization of B cells in the hepatopancreas tissue and increase F cells (arrowheads), (b) Intranuclear inclusion bodies (arrowheads) in the gill tissue, (c) Nucleus hypertrophy, early, late and Cowdry type A inclusion bodies in the lymphoid organ (arrowheads), (d) Intranuclear inclusion bodies in hyaline cells of the hematopoietic tissue (arrowheads), (e) Nucleus hypertrophy and large basophilic intranuclear inclusion bodies (arrowheads) in the cuticle epithelium, (f) Nucleus hypertrophy, large basophilic intranuclear inclusion bodies and nucleus pyknosis and karyorrhexis (arrowheads) in the connective tissue, (g) Nucleus hypertrophy and dens nucleus (arrowheads) in the striated muscles and (h) Large basophilic intranuclear inclusion bodies (arrowheads) in the digestive epithelium |
Development of cells hypertrophy that observed in the necrotic tissues was different in the viral infection stages. Eosinophilic intranuclear inclusion bodies Cowdry type A exhibited in numerous hepatopancreas epithelial cells (Fig. 4a). The tissues section showed eosinophilic enlarged nuclei, often accompanying whit cloudy inclusion surrounded by marginated chromatin. Muscular atrophy, associated with reddish discoloration of the cuticles was prominent. Histologically, the striated muscles of the affected abdominal segmental abnormality showed severe Zenker's necrosis (Fig. 4h). All tissue exhibited nucleus pyknotic and karyorrhectic in advanced infectious stages A. The presence of IHHNV was confirmed by histopathological observation.
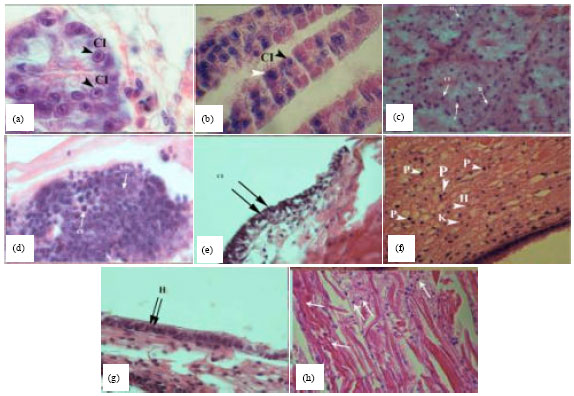 | |
| Fig. 4(a-h): | Histopathological observations of IHHNV infection in tissue shrimp include nucleus Hypertrophy (H), Cellular Degeneration (CD), Cowdry type A intranuclear Inclusion bodies (CI) and nucleus Pyknosis (P) and Karyorrhexis (K) in L. vannamei (H and E\Ph 1000X), (a) Large eosinophilic intranuclear inclusion bodies Cowdry type A in the hepatopancreas tissue (arrowheads), (b) Intranuclear inclusion bodies (arrowheads) in the gill tissue, (c) Nucleus hypertrophy in the lymphoid organ (arrowhead), (d) Intranuclear inclusion bodies and nucleus pyknosis and karyorrhexis (arrowheads) in the haematopoietic tissue, (e) Nucleus hypertrophy and intranuclear inclusion bodies (arrowheads) in the cuticle epithelium, (f) Nucleus hypertrophy, intranuclear inclusion bodies and nucleus pyknosis and karyorrhexis (arrowheads) in the connective tissue, (g) Intranuclear inclusion bodies (arrowheads) in the digestive epithelium and (h) Nucleus hypertrophy and Zenker's necrosis (arrowhead) of the striated muscles |
DISCUSSION
To detect viral diseases of Penaeidae shrimps, different methods are being used such as clinical signs, histopathology, molecular methods (Polymerase Change Reaction) and Transmission Electron Microscopy (TEM). Viral acute infections associated with mass death (100%) and without clinical symptoms (Lightner, 1996; Lightner, 1999). On the other hand, the genetic mutations due to geographic isolation may not be a suitable molecular method so by histopathological method, tissue damages caused by the pathogenic viruses were studied (Bell and Lightner, 1988; Lightner, 1996).
WSSV infection was created by Nimaviridae (Chen 1995). Other names for WSSV are Systemic Ectodermal and Mesodermal Baculovirus (SEMBV) (Wongteerasupaya et al., 1995), Rod-shaped Virus of Marsupenaeus japonicus (RV-PJ), Penaeid Rod-shaped DNA Virus (PRDV) (Inouye et al., 1996) Hypodermal and Hematopoietic Necrosis Baculo-like Virus (HHNBV) of Fenneropenaeus chinensis(Sreenivasa Rao et al., 2007). This virus is capable to infect at least 78 species, mainly decapods crustaceans including marine and fresh water shrimps, crabs, crayfish and lobster (Lightner, 1996; Flegel, 2006) and some arthropods can be the source of virus (Lo et al., 1999). It is rarely observed in non-penaeidae species, except Macrobrachium rosenbergii and Orconectes punctimanus (Peng et al., 1998).
The broodstocks L. vannamei collected from hatcheries of Bushehr province, showed typical symptoms of white spots on the inner surface of the carapace and shell similar to symptoms described by Afsharnasab et al. (2009). But the shrimps were normally feeding and no deaths were occurred, which may be due to accommodation of host with the virus (Afsharnasab et al., 2009). Juvenile and sub adult shrimps accompanied with severe mortality due to high virulence of virus but were not observed white spots on their carapace. Other signs were feeding reduction, as Saberi et al. (2008) reported the reddish body coloration on the moribund shrimp. Virulence studies show that L. vannamei has little resistance to WSSV, so when the virus has a high virulence, it could be associated with mass death (100%) (Wang et al., 2000).
Nuclear hypertrophy and cellular degeneration of the cells from ectoderm and mesoderm origin were commonly observed in moribund shrimp. Wang et al. (2005) reported that L. duorarum showed different tolerance with virus virulence in different geographical regions. Initial studies showed that clinical signs observed in M. japonicus and P. monodon infected to WSSV, after 3 days, made high mortality percentage (70-100%). Also there were differences in intensity virulent WSSV in Fenneropenaeus indicus compared to other species (especially L. vannamei) which it may be due to different sensitivity of the species, their defensive mechanism and environmental factors of the studied area. Afsharnasab et al. (2009) and Granja et al. (2003) showed that apoptotic cells reduce virus replication and control disease in L. vannamei. Apoptosis plays a critical role in development and maintenance of multicellular organisms. It has also been described as an anti-viral mechanism in both insects and vertebrates. In fact, to resistance against the immune system and to increase their outbreak, some viruses such as Baculovirus sp. produce anti-apoptotic molecules (Granja et al., 2003). Histopatholgical studies implies existence intranuclear inclusion bodies Cowdry type A in cells tissue target such as gills, lymphoid organ, hematopoietic tissue, cuticular epithelium, digestive epithelium, striated muscles and connective tissues which advanced stages were observed basophilic (H and E/Ph) (Perez et al., 2005; Afsharnasab et al., 2009). Lack of polyhydrogenic materials in WSSV structure caused basophilic color inclusion bodies but some of viruses (i.e., Parvovirus) existence polyhydrogenic material in the structure caused an eosinophilic color inclusion bodies (Afsharnasab et al., 2009). Cowdry type A, basophilic, intranuclear inclusion bodies surrounded by marginated chromatin in hypertrophied nuclei of cells in tissues of ectodermal (gills, cuticular epithelium and digestive epithelium) and mesodermal origin (hematopoietic organs, lymphoid organ, connective tissue and striated muscle) (Perez et al., 2005). However, in white-spot syndrome the Cowdry A inclusions represent an early stage of viral infection. Once infecting of these nuclei undergo further degeneration and finally develop into prominent eosinophilic and pale basophilic type inclusions.
Cells of the hepatopancreas has never shown to be infected with WSSV but enlargement and fragility of hepatopancreas tissue of contaminated shrimps, microscopic observations indicated vacuolization tissue which can be due to increased hemolymph from this organ to promote system of immunity cell (Lightner, 1996; Afsharnasab et al., 2009). Similar histopathological changes were observed among infected cultured Penaeid shrimps such as F. chinensis, M. japonicus, F. indicus, F. merguiensis and P. monodon. These observations coincided with the reports described by Wongteerasupaya et al. (1995) and Inouye et al. (1996). In addition, LOS (Lymphoid Organ Spheroids) was clearly observed in lymphoid organ that was infected by WSSV. LOS usually appears when shrimp was able to control or respond the infection to pathogenic agent in survival or chronic states (Lightner, 1996; Hasson et al., 1999). Nuclear pyknosis and karyorrhexis were observed in advance stages of WSSV and other viral severe infections, such as YHV, TSV, Lymphoid Organ Vacuolization Virus (LOVV) (Lightner, 1996). Existence of nuclear pyknosis and karyorrhexis in the hematopoietic tissue and lymphoid organ were accompanied with losses in tissue structure that implied attack viral infection (Rodriguez et al., 2003). However, in investigated samples, these lesions were associated with severe WSSV infection. Pantoja and Lightner (2003) have observed nuclear pyknosis and karyorrhexis in shrimps that were infected by WSSV experimentally. In addition, pyknosis and karyorrhexis were observed in the hematopoietic tissue of F. duorarum with exposure to WSSV experimentally (Wang et al., 2005).
There are milky moles on the surface of external cuticles (abdominal segmental) and opaque dorsal muscles in some sub adult shrimps accompanied with rostrum and abdominal segmental deformity. It was implicated IHHNV infection, but not observed death, because L. vannamei is resistant to the virus and act as a carrier (Lightner, 2003). L. vannamei led to transmission IHHNV to other parts has accompanied by economic losses (Lightner, 1999). IHHNV is one of the smallest viruses of penaeidae shrimps that identified in early 1980s and closely related to mosquito brevidensoviruses (Yang et al., 2007). Virus can infect shrimps during larval stages to adult and throughout vertical transmission or consume infected shrimps by healthy shrimps and potentially contact contaminated water (Motte et al., 2003). IHHNV usually infects ectodermal, mesodermal and rarely endodermal (Flegel, 1997). Some published reports describe that IHHNV has an affinity for various tissues and is rarely detected in endoderm derived tissues (Lightner, 1996; Tang and Lightner, 2006). However, if shrimps affected after post larval stage (sub adult), they could show symptoms such as rostrum deformity, curly antennae, carapace blistered, cuticle deformity and stunting (Tang and Lightner, 2006). RDS observed in L. vannamei by IHHNV infection experimental after 30 days (Singhapan et al., 2004). Usually RDS can affect in nursery period and growth stages of L. vannamei that associated with great economic damage, including irregular growth and difference in size (Lightner, 2003).
This study indicated that IHHNV had an affinity in the hepatopancreas of post larval of L. vannamei. Histopathological changes are formation of eosinophilic intranuclear inclusion bodies in the hypertrophied nuclei with marginated chromatin and cellular degeneration but in the present study affected sub adult showed growth retraction and deformities of the third to sixth abdominal segments and RDS, without eosinophilic intranuclear inclusion bodies formation. Eosinophilic intranuclear inclusion bodies Cowdry type A is due to existence of polyhydrogenic material in IHHNV structure (Rodriguez et al., 2003; Afsharnasab et al., 2009). These lesions were similar to those of RDS described in P. monodon, M. rosenbergii and L. stylirostris with IHHNV infection by Hsieh et al. (2006). So the presence of IHHNV from both hatcheries and grow-out shrimp farms (L. vannamei) indicated that this virus could slowly growth and severe difference in size which may be due to feeding reduction. Histopathological studies of gills, hematopoietic tissue, cuticular epidermis, digestive epithelium, lymphoid organ and connective tissue of infected shrimps were evidence of hypertrophy, cellular degeneration and formation intranuclear inclusion bodies that in advanced stages were together with nuclear pyknosis and karyorrhexis. Also, histopathological changes in the striated muscles of infected shrimp exhibited severe Zenker's necrosis. Muscular lysis was sometimes found in affected fiber accompanied with inclusion bodies while Hsieh et al. (2006) reported no observation inclusion bodies in striated muscles of M. rosenbergii.
Lightner (1996) reported HPV infections in L. vannamei by parvo or parvo-like viruses. He described that gross signs of HPV may not be specific, but in severe infections may include an atrophied heptopancreas, reduce growth rate and anorexia. The characteristic lesion of HPV infection is presence of prominent basophilic or eosinophilic intranuclear inclusion bodies associated with cap formation in the hypertrophied nuclei of hepatopancreatic tubular epithelial cells (Lightner, 1996). But the result of this study is quite different, where neither basophilic intranuclear inclusion bodies nor cap formation were observed.
White spots were observed on the cuticle surface of infected shrimps in both, WSSV and IHHNV. In WSSV infection, white spots were only observed on internal and external of surfaces carapace while in IHHNV infection, white plaque observed on the third to sixth abdominal segments. Cowdry type A inclusions were found in infected shrimp with WSSV similar to the appearance of intranuclear inclusions caused by IHHNV. Quere et al. (2002) also reported that inclusion bodies of WSSV could be easily mistaken as by of IHHNV but histopathological findings (H and E\Ph) showed that intranuclear inclusion bodies Cowdry type A were as basophilic in WSSV infection while in IHHNV infection observed as eosinophilic (Afsharnasab et al., 2009). Infected hepatopancreas tissues by WSSV showed severe vacuolization that was due to high activity it, but Cowdry type A inclusion intranuclear was not observed.
ACKNOWLEDGMENTS
We would like to express our gratitude to the assistance provided by the staff of Iranian Shrimp Research Center, Dr. Aienjamshid. Mr. Nazaei and Mr. Moaref for help us.
REFERENCES
- Afsharnasab, M., R. Mortezaei, V. Yegane and B. Kazemi, 2009. Gross sign, histopathology and polymerase chain reaction observations of white spot syndrome virus in shrimp specific pathogen free Litopeneaus vannamei in Iran. Asian J. Anim. Vet. Adv., 4: 297-305.
CrossRefDirect Link - Couch, J.A., 1974. Free and occluded virus, similar to baculovirus, in hepatopancreas of pink shrimp. Nature, 247: 229-231.
Direct Link - Flegel, T.W., 1997. Major viral diseases of the black tiger prawn (Penaeus monodon) in Thailand. World J. Microbiol. Biotechnol., 13: 433-442.
CrossRef - Flegel, T.W., 2006. Detection of major penaeid shrimp viruses in Asia, a historical perspective with emphasis on Thailand. Aquaculture, 258: 1-33.
CrossRefDirect Link - Granja, C.B., L.F. Aranguren, O.M. Vidal, L. Aragon and M. Salazar, 2003. Does hyperthermia increase apoptosis in White Spot Syndrome Virus (WSSV)-infected Litopenaeus vannamei? Dis. Aquat. Org., 54: 73-78.
CrossRefDirect Link - Hasson, K.W., D.V. Lightner, L.L. Mohney, R.M. Redman and B.M. White, 1999. Role of lymphoid organ spheroids in chronic Taura syndrome virus (TSV) infections in Penaeus vannamei. Dis. Aquat. Org., 38: 93-105.
Direct Link - Lightner, D.V., 1999. The penaeid shrimp viruses TSV, IHHNV, WSSV and YHV: Current status in the Americas, available diagnostic methods and management strategies. J. Applied Aquacult., 9: 27-52.
CrossRefDirect Link - Lightner, D.V., 2003. The Penaeid shrimp viral pandemics due to IHHNV,WSSV, TSV and YHV: History in the Americas and current status. Aquaculture and pathobiology of crustacean and other species. Proceedings of the 32nd UJNR Aquaculture Pane Symposium, November 17-18, 2003, Davis and Santa Barbara, California, USA., pp: 1-20.
- Lightner, D.V., R.M. Redman and T.A. Bell, 1983. Infectious hypodermal and hematopoietic necrosis: A newly recognized virus disease of penaeid shrimp. J. Invertebrate Pathol., 42: 62-70.
CrossRef - Lightner, D.V., R.M. Redman, T.A. Bell and J.A. Brock, 1983. Detection of IHHN virus in Penaeus stylirostris and P. vannamei imported into Hawaii. J. World Mariculture Soc., 14: 212-225.
CrossRefDirect Link - Lo, C.F., H.C. Hsu, M.F. Tsai, C.H. Ho, S.E. Peng, G.H. Kou and D.V. Lightner, 1999. Specific genomic DNA fragment analysis of different geographical clinical samples of shrimp white spot syndrome virus. Dis. Aquat. Org., 35: 175-185.
Direct Link - Mari, J., J.R. Bonami and D.V. Lightner, 1993. Partial cloning of the genome of infectious hypodermal and hematopoietic necrosis virus, an unusual parvovirus pathogenic for Penaeid shrimps: Diagnosis of the disease using a specific probe. J. General Virol., 74: 2637-2643.
PubMed - Fouzi, M.N.M., M. Shariff and F.M. Yusoff, 2012. Stress quantification in Penaeus monodon exposed to different levels of ammonia and subsequent infection to WSSV. Res. J. Vet. Sci., 5: 12-24.
CrossRefDirect Link - Motte, E., E. Yugcha, J. Luzardo, F. Castro and G. Leclercq et al., 2003. Prevention of IHHNV vertical transmission in the white shrimp Litopenaeus vannamei. Aquaculture, 219: 57-70.
CrossRef - Pantoja, C.R. and D.V. Lightner, 2003. Similarity between the histopathology of white spot syndrome virus and yellow head syndrome virus and its relevance to diagnosis of YHV disease in the Americas. Aquaculture, 218: 47-54.
CrossRef - Peng, S.E., C.F. Lo, C.H. Ho, C.F. Chang and G.H. Kou, 1998. Detection of white spot baculovirus (WSBV) in giant freshwater prawn, Macrobrachium rosenbergii, using polymerase chain reaction. Aquaculture, 164: 253-262.
CrossRef - Perez, F., A.M. Volckaert-Filip and J. Calderon, 2005. Pathogenicity of white spot syndrome virus on postlarvae and juveniles of Penaeus (Litopenaeus) vannamei. Aquaculture, 250: 586-591.
CrossRef - Quere, R., T. Commes, J. Marti, J.R. Bonami and D. Piquemal, 2002. White spot syndrome virus and infectious hypodermal and hematopoietic necrosis virus simultaneous diagnosis by miniarray system with colorimetry detection. J. Virol. Methods, 105: 189-196.
CrossRefPubMedDirect Link - Rodriguez, J., B. Bayot, Y. Amano, F. Panchana, I. de Blas, V. Alday and J. Caleron, 2003. White spot syndrome virus infection in cultured Penaeus vannamei (Boone) in Ecuador with emphasis on histopathology and ultrastructure. J. Fish Dis., 26: 439-450.
PubMed - Saberi, A.M., M. Bandehpour, M. Afsharnasab, E. Ghayour, S.A. Yousefi Namin and B. Kazemi, 2008. Designing and introduce a diagnostic kit for detection of white spot syndrome virus in cultured Penaeus indicus in Iran. Pak. J. Biol. Sci., 11: 2660-2664.
CrossRefDirect Link - Sreenivasa Rao, M., B. Rajitha, E. Pavitra and N. Anjaneyulu, 2007. Identification and changes of copper profile in different tissues during vitellogenesis of white shrimp Litopenaeus vannamei. J. Biol. Sci., 7: 989-992.
Direct Link - Singhapan, J., C. Limsuwan and N. Chuchird, 2004. Effect of infectious hypodermal and hematopoietic necrosis virus (IHHNV) on growth, survival rate and histopathological changes of Pacific white Shrimp (Litopenaeus vannamei). Kasetsart J. Nat. Sci., 38: 236-240.
Direct Link - Tang, K.F.J. and D.V. Lightner, 2006. Infectious hypodermal and hematopoietic necrosis virus (IHHNV) related sequences in the genome of the black tiger prawn Penaeus monodon from Africa and Australia. Virus Res., 118: 185-191.
PubMed - Wang, Q., B.T. Poulos and D.V. Lightner, 2000. Protein analysis of geographic isolates of shrimp white spot syndrome virus. Arch. Virol., 145: 263-274.
Direct Link - Wongteerasupaya, C., J.E. Vickers, S. Sriurairatana, G.L. Nash and A. Akarajamorn et al., 1995. A non-occluded, systemic baculovirus that occurs in cells of ectodermal and mesodermal origin and causes high mortality in the black tiger prawn Penaeus monodon. Dis. Aquat. Org., 21: 69-77.
CrossRefDirect Link - Yang, B., X.L. Song, J. Huang, C.Y. Shi and L. Liu, 2007. Evidence of existence of infectious hypodermal and hematopoietic necrosis virus in Penaeid shrimp cultured in China. Vet. Microbiol., 120: 63-70.
CrossRef