
C. d’Andre Hirwa
Department of Animal Genetics, Breeding and Reproduction, College of Animal Science, South China Agricultural University, Guangzhou 510642, China
W. Paul
Department of Animal Genetics, Breeding and Reproduction, College of Animal Science, South China Agricultural University, Guangzhou 510642, China
W. Yan
Department of Animal Genetics, Breeding and Reproduction, College of Animal Science, South China Agricultural University, Guangzhou 510642, China
C. Luo
Department of Animal Genetics, Breeding and Reproduction, College of Animal Science, South China Agricultural University, Guangzhou 510642, China
Q. Nie
Department of Animal Genetics, Breeding and Reproduction, College of Animal Science, South China Agricultural University, Guangzhou 510642, China
G. Yang
Department of Animal Genetics, Breeding and Reproduction, College of Animal Science, South China Agricultural University, Guangzhou 510642, China
X. Zhang
Department of Animal Genetics, Breeding and Reproduction, College of Animal Science, South China Agricultural University, Guangzhou 510642, China
Asian Journal of Animal Sciences
Year: 2009 | Volume: 3 | Issue: 3 | Page No.: 85-91







ABSTRACT
In this study, we describe the insertion/deletion (indel) polymorphism of the THRSP gene evaluated using 1% agarose gel and 8% polyacrylamide gel electrophoresis in a sample of (17 breeds of chicken including 3 foreign breeds and 14 chinese native breeds). For the THRSPα gene indel polymorphism result, allele frequencies varied, between chinese native chicken and a foreign chicken. A high frequency of allele B (0.91) and a low frequency of allele A (0.09) were observed in Leghorn chicken, which has been selected for laying performance. Based on variation in allele frequencies among populations, either the low frequency of allele A or the high frequency of allele B could be linked to fatness trait of the chicken. In conclusion, THRSPα gene might be used as a molecular marker in high quality broiler assistant selection breeding program.
PDF Abstract XML References Citation
How to cite this article
C. d’Andre Hirwa, W. Paul, W. Yan, C. Luo, Q. Nie, G. Yang and X. Zhang, 2009. Allelic Frequency in Chicken Thyroid Hormone Responsive Spot 14 Alfa Gene (THRSPα). Asian Journal of Animal Sciences, 3: 85-91.
DOI: 10.3923/ajas.2009.85.91
URL: https://scialert.net/abstract/?doi=ajas.2009.85.91
DOI: 10.3923/ajas.2009.85.91
URL: https://scialert.net/abstract/?doi=ajas.2009.85.91
INTRODUCTION
The domestic chicken continues to serve mankind as widely as using non-mammal biological model, an important global source of high quality protein from meat and eggs. The chicken has received less attention for comparative and functional genomic lately, mainly because of a low number of expressed sequence taqs and the lack of a completed genome sequence (Cogburn et al., 2004). Over the past 5 years, chicken genome has led to a tremendous accumulation of data, which has resulted in a detailed linkage map (Groenen et al., 2000) and fine mapping a large number of economically important quantitative traits locus. Furthermore, additional sources have been developed that made it possible to integrate genetic and physical mapping data and allowed the commencement of the development of physical maps as well (Crooijmans et al., 1996). Understanding the genetic causes of phenotypic variation still remains a major challenge for animal genetics. Allelic variation resulting can potentially contribute to phenotypic variation, for example, the complementation and interaction of different alleles in hybrids are hypothesized to be a component of the genetic basis for heterotic phenotypes (Birchler et al., 2006; Springer and Stupar, 2007). In hundreds of cases, a single DNA sequence polymorphism affecting a protein coding sequence has been linked to a clear simple Mendelian phenotype (Botstein and Risch, 2003) and, for a much smaller but increasing number of cases, to more complex phenotypes (Glazier et al., 2002). The selection of single nucleotide polymorphism for more intensive investigation can be performed by determining single nucleotide polymorphism allele frequencies for DNA pools of a population (Liu et al., 2008). There are several aspects regarding chicken that make it extremely well suited for genetics experiments such as short generation interval, the ability to generate large full sib pedigrees and the ease of obtaining large quantities of genome DNA from the nucleated red blood cells. Chicken is an intermediate evolutionary model organism because there are many conserved regions between the chicken and human genome. It is also known that in chicken many genes play different role in growth performance, meat and egg production.
The S14 gene referred to as thyroid hormone responsive Spot 14 protein (THRSP), encodes a small acidic protein that was discovered in earlier studies of thyroid hormone action on hepatocytes. THRSP is strongly implicated as a transcription factor that controls expression of major lipogenic tissue such as liver, fat and mammary gland. The previously reported chicken related proteins are labeled chicken Alfa and Beta (Wang et al., 2004). The THRSPα gene was found on chromosome 1 at location 197836316-197838040 in chicken (Gene bank http://www.ncbi.nlm.nih.gov and http://www.genome.ucsc.edu/cgi.bin/hgBlat), the chromosome domain where the genes exist is related with the adiposity and the gene is thought to possess important function for the production of fat. The chromosome domain contains sebum traits points and the ventral fat quantity traits points (Ikeobi et al., 2002). The Spot 14 gene is specifically expressed in tissue that produce lipids for use as a metabolic fuel including lactating mammary, white and brown adipose tissue and liver. In liver, its expression is rapidly induced by stimuli for increased long chain fatty acid synthesis including thyroid hormone, glucose and insulin and is inhibited by glucagon and other stimuli that reduce lipogenesis. These observations coupled with immunohistochemical localization of Spot 14 to the nucleus, prompted the hypothesis that Spot 14 functioned in the tissue-specific regulation of genes encoding the enzymes of lipid synthesis (Joel et al., 1998). The genomic structure of THRSPα, which includes a TATA box, exon 1, represents the short 5'-Untranslated region (UTR) and the protein coding region, while exon 2 represents the 3'-UTR. Wang et al. (2004) reported the identification of a synteny group in chicken genomic DNA that includes THRSP and two flanking genes (NADH dehydrogenase (NDUFC2) and glucosyltransferase (ALG8)). THRSPα reported to be associated with growth, as well as abdominal fat traits of chickens (Cao et al., 2007; Wang et al., 2004; Zhan et al., 2009). In this study, we described genetic differences among 17 chicken breeds using DNA marker together with the method of PCR products and primers genotyped by polyacrylamide gel electrophoresis and Single Strain Conformation Polymorphism (SSCP). The objectives was to compare genetic variability within and among chicken breeds and then to identify line specific and breed specific DNA as genetic markers for estimation of relationships among 17 breeds.
MATERIALS AND METHODS
Chicken Populations
Genomic polymorphism of THRSPα indel was studied in seventeen chicken breeds by randomly selection. The Chinese native chicken that were included Xinghua (18), Quingyan (26), Xiangji (23), Mayu (36), Yugan (29), Chahua (26), Wuding (26), Gushiu (30), Lingshan (29), Tianhe silkies (18), Red Jungle (29), Tibet (30), Yangshan (34), Beijing fatty (31) while the foreign breeds were Hailan (23), White recessive (37) and White Leghorn (25). These populations represent different breeds from different part of China in 2004. Some chickens were kept in South China Agricultural University farm. The blood samples were collected from different farms. Leghorn is a layer breed and has been bred as laying type for many years. White Recessive Rock is fast growing broiler line and has been bred as meat type for many years.
Blood Samples Collection and DNA Extraction
Blood samples of 1-1.5 mL were collected from the wing vein with 2.5 mL Syringes, containing EDTA as anti-coagulating agent. The blood cells from individual birds were obtained from blood by centrifugation and serum separation. The extraction of the DNA was from the fresh blood or the blood that has been stored at -20°C.
THRSPα Analysis
Genomic DNA sample (40 ng) was amplified by PCR using forward primer (Deletion F:5’-GCC TCC GTC ACC GAT CAG-3’) and reverse primer (Deletion R: 5’-CGG TCA GAA CCT GCT GCAA-3’) at 0.2U Taq DNA polymerase, 25 mM MgCl2, 10 mM dNTP and 10xPCR Buffer. The PCR was performed of 33 cycles of 30 sec at 94°C, 30 sec at 59.5°C and 1 min at 72°C after denaturation at 94°C for 2 min, final extension was carried out for 10 min. The forward and reverse primer produce a 127 or 136 bp. The 136 bp is representative of THRSPα AA genotype and 127 bp is representative of THRSPα BB genotype, which is indicated 9 bp deletion. The amplified DNA fragments were separated on 1% agarose gel and stained with ethidium bromide. The amplified pattern was visualized on a UV transilluminator and photographed. After PCR product, genotypes was determined by 30% polyacrylamide solution (Acrylamide and N’N”methylene bisacrylamide) and 6 mL of 50% glycerol; 3 mL of 10xTBE and 60 μL of 30% TEMED and 600 μL of 10% Ammonium per sulfate (AP) were also added to the mixture and carefully poured into the mold made up of 20x20 cm, double glass plates clamped together by iron-clips. After this, two plastic combs, each with 20 flat-teeth was gently inserted into the upper part of the glass. A wedge gel of 0.5 mm thick was used. Before loading of the amplified products, 8 μL of blue loading buffer dye were prepared into tubes and 2 μL of the product were added, gently swirl and loaded it to each lane through the glasses. The PCR products were separated on polyacrylamide gel electrophoresis that lasted for 5 h at 180 V, 8°C. The amplified pattern was visualized on an ultraviolet transilluminator and photographed. After the PCR products were obtained, genotypes were determined by polyacrylamide gel electrophoresis.
Statistical Analysis
The allelic frequencies and genotype frequencies were estimated by GENEPOP software package (Raymond and Rousset, 1995). Data for all breeds were subjected to Molkin v2.0 software (Gutierrez et al., 2005) for genetic analysis of populations and for quantifying the inbreeding effect of population substructure, Wright (1931) defined the fixation index (Fst). It equals the reduction in heterozygosity with random mating at any one level of a population hierarchy relative to another. The fixation index is a useful index of genetic differentiation (Daniel and Clark, 1997).
RESULTS
In this study, the mean genotypic frequencies THRSPα indel polymorphism revealed differences between native and foreign breeds, variation among breeds with respect to genotypic frequencies and relatedness of certain native breeds. On average, over all breeds of chicken, the B allele (0.65) was more frequent overall than the A allele. The most frequent genotype (Table 1) overall was AB (51%). The least frequent genotype was AA (9%).
| Table 1: | Genotypic and allelic frequencies THRSPα indel polymorphism in 17 chicken breeds |
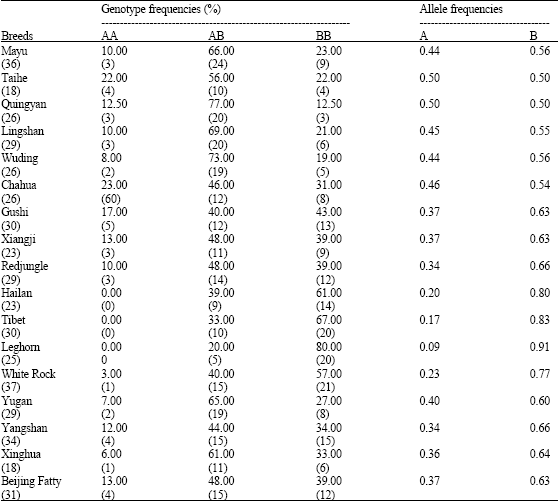 | |
| Table indicates genotype and allelic frequencies. Numbers in parenthesis represent the total individual and the numbers of each genotype detected in this experiment | |
Genotypic and allelic frequencies differed between native and foreign breeds. The A allele was more frequent in native breeds. The B allele was more frequent in foreign breeds. Thus, for Quingyan and Taihe chickens the frequency of the A allele was 0.5 but it was only 0.09 in Leghorn chickens. In native breeds, AB was the most frequent genotype and AA was in frequent. For example, for Quingyan chickens, frequencies of the AB and AA genotypes were 77 and 12.5%, respectively. In contrast, in foreign breeds, the most frequent genotype was BB. The AA genotype was rare or absent. In Leghorn chickens, the frequencies of the BB and AA genotypes were 80 and 0%, respectively. Variation in genotypic and allelic frequencies was greater in native breeds than in foreign breeds. For example, among foreign breeds the frequency of the A allele varied from 9% (Leghorn) to 23% (White Recessive Rock). The frequency of the BB genotype varied from 57% (White Recessive Rock) to 80% (Leghorn). Among native breeds, the frequency of the A allele varied from 34% (Yangshan) to 50% (Taihe and Quingyan) and the frequency of genotype AB varied from 40% (Gushiu) to 77% (Quingyan). Some native breeds appeared closely related. For example, allelic and genotypic frequencies were similar for the breeds Quingyan and Taihe. For these two breeds, the frequency of the A allele was 0.5 and the frequency of the AB genotype varied from 56% to 77%. Allelic and genotypic frequencies also were similar among Mayu, Lingshan and Wuding breeds. The frequency of the A allele varied from 8 to 10%. The frequency of the AB genotype varied from 66 to 73%.
The highest genotype was recorded for AB genotype, followed by BB and AA. The differences between Chinese native chickens and foreign chicken as shown in Table 1 is reported the genotype frequencies; foreign chickens are low in AA and higher in BB. The genotype frequencies of allele A were almost the same in all Chinese native breeds, except 4 breeds (QY = 0.50, YS = 0.36, RJ = 0.34, TH = 0.50, BF = 0.37); for foreign chicken A allele was low (LG = 0.09, RW = 0.23). The Allele B was highest in foreign chicken (LG = 0.91; WR = 0.77; HL = 0.80), for Chinese chicken Tibet chicken was highest (0.83). However, distinctive differences in allele frequencies were observed between native Chinese chicken and a foreign chicken. A high frequency of allele B (0.91) and a low frequency of allele A (0.09) were observed in the Leghorn, which has been selected for laying performance. Based on the allele frequencies among different populations, either the absence of allele A or high allele frequency of allele B could be linked to fatness trait of the chicken.
Genetic analysis using molecular Coancestry among chicken breeds; base on wrist statistics Fst = 0.069, indicated that there is moderate genetic differentiation among chicken breeds.
DISCUSSION
In this study, the genotype and allele frequencies among 17 chicken breeds based on THRSPα indel polymorphism have been described. THRSP is reportedly involved in lipogenesis and energy metabolism (Cogburn et al., 2004). THRSP has two forms: alpha and beta paralogs in chickens and polymorphic alleles involving tandem repeats (of either 9 or 6 bp) were found in the putative protein coding region of the chicken THRSPα (a 9 bp indel) and THRSP beta (a 6 bp indel) genes (Wang et al., 2004). The study showed that the THRSPα locus is associated with abdominal fat traits in a broiler times. Leghorn resource population (Wang et al., 2004). These observations support a role of THRSP in control of lipogenesis and expression of abdominal fat traits in the domestic chicken. Chicken Spot 14 (THRSP) was first identified as a differentially-expressed sequence taq from microarray analysis of livers from chickens, divergently selected for fast or slow growth rate (Cogburn et al., 2000, 2003). THRSP involved in lipid metabolism and immunity (Wang et al., 2007). THRSP expressed in lipogenic tissues, is a transcription factor to regulate gene expression of rate-limiting enzymes in lipogenesis. Two THRSP isoforms, THRSPa and THRSPb, were detected at cDNA levels in chickens and ducks (Zhan et al., 2009). The relationships between the polymorphism of chicken (White Recessive Rock x Chinese Native Xinghua; THRSPα and growth, carcass trait) were reported to be significantly not different (Yan et al., 2004). The polymorphism involves either an insertion or deletion in a conserved region of the spot 14 gene implicated in homodimer formation (Cunningham et al., 1997) and result in decreased abdominal fat weight. In support of a role of this gene in obesity (Zhu et al., 2005) reported on a correlation between body mass index and the regulation of Spot 14 mRNA levels in Human white adipose tissues, found out that Spot 14 mRNA level were markedly down regulated by fasting in nonobese subjects and minimal down-regulated by fasting in obese subjects. THRSPα was reported to be associated with growth development and lipid metabolism because of significant correlation between the indels in the coding region of THRSP alpha and growth, as well as abdominal fat traits of chickens (Cao et al., 2007; Wang et al., 2004; Zhan et al., 2009). In the present study, THRSPα polymorphism in 17 chicken breeds by randomly selection was however observed. Chinese native chicken had high AB genotype frequency (Quingyan = 50%) than foreign breed (White Leghorn (20%) and (40%) for White recessive Rock). This study showed that Quingyan, Yangshanji, Lingshan, Taihe silkies and Wuding chickens were closer than others. They showed high genotype frequencies than Mayu, Yugan, Chahua, Gushiu, Xiangji. Beijing fatty, Red Jungle, Hailan and White Recessive Rock were closer than others. Tibet, White Leghorn and Xinghua chicken were closer. Wang and collegues have reported that in the broilerxLeghorn, THRSPα homozygotes present a higher fat content than heterozygotes (Wang et al., 2004). Yan et al. (2004) have also reported that THRSPα homozygotes (BB) have a high fat band width than THRSPα homozygotes (AA) p<0.05 and homozygotes (BB) and heterozygotes (AB) present a high fat content than homozygotes (AA). Regarding genotypes AA, AB and BB, there were slight differences. The highest among breeds was AB (51.4%), BB was (38.1%) and the least was AA (9.21%). The highest BB was in White Leghorn (80%) while the lowest was in Quingyan (12.5%). As we observe F-statistics level among chicken breeds, the data shows moderate genetic divergence (FST = 0.06). In conclusion, there are distinctive differences in THRSPα allele frequencies, which were observed between native Chinese chicken and a foreign chicken. A high frequency of allele B (0.91) and a low frequency of allele A (0.09) were observed in the Leghorn, which has been selected for laying performance. Based on the allele frequencies among different populations, either the low of allele A or high allele frequency of allele B could be linked to fatness trait of the chicken. For improvement of the breeding, having unique attributes like fitness with high meat quality efforts should be firstly made to strengthen the conservation programme, including constructing core population.
In conclusion, this study suggests that THRSPα gene might be used as a molecular marker in high quality broiler assistant selection breeding program.
ACKNOWLEDGMENT
I would like to express my sincere thanks to UNESCO in Rwanda, Paris and China for supporting this study.
REFERENCES
- Botstein, D. and N. Risch, 2003. Discovering genotypes underlying human phenotypes: Past successes for mendelian disease, future approaches for complex disease. Nat. Genet., 33: 228-237.
PubMedDirect Link - Birchler, J.A.H. and S. Chudalayandi, 2006. Unraveling the genetic basis of hybrid vigor. Proc. Natl. Acad. Sci. USA., 103: 12957-12958.
Direct Link - Cao, Z.P., S.Z. Wang, Q.G. Wang, Y.X. Wang and H. Li, 2007. Association of Spot14α gene polymorphisms with body weight in chicken. Poult. Sci., 86: 1873-1880.
Direct Link - Cogburn, L.A., X. Wang, W. Carre, L. Rejto, S.E. Aggrey, M.J. Ducros, J. Simon and T. E. porter, 2004. Functional genomics in chickens: Development of integrated-system micro arrays for transcriptional profiling and discovery of regulatory pathways. Comp. Funct. Genomics, 5: 253-261.
CrossRef - Cunningham, B.A., M. Malony and W.B. Kinlaw, 1997. Spot 14 protein-protein interaction: Evidence for both Homo- and heterodimer formation in vivo. Endocrinology, 138: 5184-5188.
PubMedDirect Link - Crooijmans, R.P.M.A., F.A. Groen and A.J.A. Kampen, 1996. Microsatellites polymorphism in commercial broiler and layer lines estimated using pooled blood samples. Poult. Sci., 75: 904-909.
PubMed - Glazier, A.M., J.H. Nadeau and T.J. Aitman, 2002. Finding genes that underlie complex traits. Science, 298: 2345-2349.
PubMedDirect Link - Groenen, M.A., H.H. Cheng, N. Bumstead, B.F. Benkel and W.E. Briles et al., 2000. A consensus linkage map of the chicken genome. Genome Res., 10: 137-147.
PubMedDirect Link - Gutierrez, J.P., L.J. Royo, I. Lvarez and F. Goyache, 2005. MolKin v2.0: A computer program for genetic analysis of populations using molecular coancestry information. J. Hered., 96: 718-721.
CrossRefDirect Link - Ikeobi, C.O., J.A. Woolliams and D.R. Morrice, 2002. Quantitative trait loci affecting fatness in the chicken. Anim. Genet., 33: 428-435.
Direct Link - Liu, G.Q., X.P. Jiang, J.Y. Wang, Z.Y. Wang, G.Y. Liu and Y.J. Mao, 2008. Analysis of genetic diversity of yangzhou chicken by microsatellite markers. Int. J. Poult. Sci., 7: 1237-1241.
CrossRefDirect Link - Zhu, Q., G.W. Anderson, G.T. Mucha, E.J. Parks, J.K. Metkowski and C.N. Mariash, 2005. The Spot 14 protein is required for the novo lipid synthesis in the lactating mammary gland. Endocrinology, 146: 3343-3350.
PubMedDirect Link - Raymond, M. and F. Rousset, 1995. GENEPOP (version 1.2), population genetics software for exact tests and ecumenicism. J. Hered., 86: 248-249.
Direct Link - Springer, N.M. and R.M. Stupar, 2007. Allelic variation and heterosis in maize: How do two halves make more than a whole? Genome Res., 17: 264-275.
CrossRefPubMedDirect Link - Wang, X., W. Carre, H. Zhou, S.J. Lamont and L.A. Cogburn, 2004. Duplicated Spot 14 genes in the chicken: Characterization and identification of polymorphisms associated with abdominal fat traits. Genetics, 332: 79-88.
CrossRef - Wright, S., 1931. Evolution in Mendelian populations. Genetics, 16: 97-159.
CrossRefPubMedDirect Link - Zhan, K., G.Y. Xu, Y.Y. Xu and R.H. Zhao, 2009. Progresses on thyroid hormone responsive spot 14 (THRSP) genes in chickens and ducks. Yi Chuan., 31: 131-136.
PubMed