
M. Orunmuyi
Department of Animal Science, Ahmadu Bello University, Zaria, Nigeria
O.O. Oni
National Animal Production Research Institute, Ahmadu Bello University, Shika, Zaria, Nigeria
I.A. Adeyinka
National Animal Production Research Institute, Ahmadu Bello University, Shika, Zaria, Nigeria
O.E. Asiribo
Department of Statistics, University of Agriculture, Abeokuta, Nigeria
Asian Journal of Animal Sciences
Year: 2007 | Volume: 1 | Issue: 2 | Page No.: 76-81







ABSTRACT
A study was conducted to determine the relationship between Plasma Alkaline Phosphatase (PAP) activity and reproductive traits in two strains of Rhode Island chickens. Mean enzyme activity in the cocks was 133.07 I.U for strain A and 134.78 I.U for strain B. Values of percentage fertile (PCF), percentage hatch (PCH) and percentage hatchability (PCHBLT) were 80.47, 45.68 and 53.41 in strain A, corresponding values in strain B were 85.94, 51.79 and 56.67. Heritability estimate was high for PAP in both strains (0.57) but low to moderate values were obtained for reproductive traits except percent fertility in strain A (0.56). Genetic correlations between PAP and PCF, PCH and PCHBLT were high and negative in strain A but low and positive values except PCHBLT were obtained in strain B. Phenotypic correlations between PAP and PCF, PCH and PCHBLT were generally low in magnitude but follow the direction of genetic correlations. The enzyme activity can be useful in the selection of the cocks for improved fertility and or hatchability of fertile eggs.
PDF Abstract XML References Citation
How to cite this article
M. Orunmuyi, O.O. Oni, I.A. Adeyinka and O.E. Asiribo, 2007. Genetic Parameter Estimates for Plasma Alkaline Phosphatase Activity and Reproductive Traits in Two Strains of Rhode Island Chickens. Asian Journal of Animal Sciences, 1: 76-81.
DOI: 10.3923/ajas.2007.76.81
URL: https://scialert.net/abstract/?doi=ajas.2007.76.81
DOI: 10.3923/ajas.2007.76.81
URL: https://scialert.net/abstract/?doi=ajas.2007.76.81
INTRODUCTION
Fertility and hatchability are two important factors used to determine the reproductive performance of birds. Fertility is the proportion of number of eggs that are capable of developing into chicks over the total number incubated. Fertility is usually determined by candling at 18th day of incubation though it can be determined earlier at the 6th day of incubation.
Hatchability refers to the percentage of eggs hatched into viable chicks after 21 days of incubation. Hatchability is either determined on the basis of all eggs set (% hatch) or on the basis of fertile eggs set after candling (% hatchability). Both fertility and hatchability are always difficult to determine. Low fertility and a high degree of embryonic death are often confused. Most times when determining fertility in hatcheries, errors are usually made because fertility and hatchability of fertile eggs are frequently considered as hatchability of all eggs set.
Fertility and hatchability are sensitive to both environmental and genetic influences. Among the environmental factors affecting fertility and hatchability are season, age of hen, age at sexual maturity, nutrition, mating ratio, diseases, hatching egg condition and incubator conditions.
Both fertility and hatchability are also sensitive to genetic influence. North (1978) reported that fertility is inherited to some extent as securing good hatchability seems to depend more on the breeders than the incubator. However, most reported estimates of genetic parameters in literatures indicated that the heritability of fertility is low (Beaumont, 1992; Poivey et al., 2001). However, Chao and Lee (2001) reported a high heritability estimate for fertility in one of their two experiments (0.459±0.297 for experiment 1 and 0.234±0.227 for experiment 2).
Much genetic research is now directed towards the investigation of the relationship between physiological, biochemical and metabolic products/markers to the productive efficiency of farm animals. Biochemical traits, including blood groups, blood proteins and enzymes have been studied with a view to explaining the physiological basis of performance traits and the effect of heterosis.
Alkaline Phosphatase (AP) orthophosphoric acid monoester phosphohydrolase (EC 3.1.3.1) consists of a group of heterogenous isoenzymes that catalyze the hydrolysis of monophosphate esters at alkaline pH (Syakalima et al., 1998). The highest activity of the enzyme is found in the intestine mucous membrane, next in placenta, kidneys, bones, lungs and spleen (Szeniawska and Kurye, 1987). Its activity is low or does not exist in muscles, mature connective tissues, unossified cartilage and erythrocytes (Moss, 1974).
Very few literatures are available concerning the relationship between plasma alkaline phosphatase activity and reproductive traits in chickens.
Based on differences due to breed in fertility and hatchability of eggs and the paucity of information on the relationship between plasma alkaline phosphatase activity and reproductive traits, this study was designed with the following objectives.
| • | To determine variation in the reproductive traits in the two strains of Rhode Island chickens. |
| • | To estimate heritability of plasma alkaline phosphatase and reproductive traits. |
| • | To estimate genetic correlation between plasma alkaline phosphatase activity and reproductive traits. |
MATERIALS AND METHODS
Source of Experimental Animals
Two strains of Rhode Island chickens (Red and White) belonging to the breeding unit of Poultry Research Programme of National Animal Production Research Institute (NAPRI) Shika, Nigeria were used for the study. The study was conducted in the year 2005 at the Institute. The birds were obtained from the selected lines (male and female lines) and are denoted as strain A and strain B, respectively. Strain A has gold plumage and strain A has silver plumage. NAPRI is geographically located between latitude 11° and 12°N and longitude 7 and 8°E at an altitude of 640 m above sea level. This area is found in the Northern Guinea Savanna zone of Nigeria with an average annual precipitation of 1100 mm. Rainfall starts in late April or sometimes early May, reaches peak between June and September and lasts till October. The harmattan period which start from mid October through January is immediately followed by dry hot weather from February to May. Temperature ranges are harmattan 14 to 30°C and hot season 21 to 36°C. Relative humidity varies from approximately 21% during harmattan to 37% during the hot season. The average temperature and humidity during the wet season are 24.8°C and 77%, respectively.
Management Practices
On the day of hatching all chicks were wing-banded and pedigreed by Sire and Dam. Chicks were brooded in conventional floor pens and fed on chicks mash with 20% CP content. At 8 weeks, grower’s ration (16% CP) was fed until the birds reached point of lay at which time, layers mash was introduced (17% CP). Both feed and water were given ad-libitum. The birds were vaccinated against known and common poultry diseases such as Marek’s, New castle, Infectious Bursal and Fowl-pox. Other routine medication and management operations carried out include anti-coccidial medication, deworming and debeaking.
At the onset of lay, the hens were transferred to the laying cages to monitor their egg production up to 280 days of age.
Mating Procedures
The birds were selected from those monitored for egg production traits and were brought down from the cage and transferred to deep-litter floor pens at 280 days of age.
The birds were maintained on a sire family basis, each strain consisting of 28 sire families with a male to female ratio of 1:9. Trapnests were placed in each pen. On each egg laid, the strain, pen number and hen’s tag number were recorded. All birds received a layer breeder ration with feed and water ad libitum.
Collection of Blood Samples
Blood samples were drawn from each cock twice, one at the beginning of egg collection and one at the end of collection. Plasma alkaline phosphatase activity was determined according to Kind and King (1954).
Hatching
A total of 1781 eggs from strain A and 1604 eggs from strain B were set. The eggs were set on a sire family and individual hen basis and candled on 18th day of incubation to determine fertility. Data on` fertility and hatchability were recorded for seven hatches on each strain. Percent fertility, percent hatch and percent hatchability were calculated as follows:
Genetic Parameter Estimates
Genetic parameters were estimated using the sire model whereby the variance component was partitioned into those due to sire or environment. In this design, the statistical model used was:
Where;
| yij | = | The record of the jth progeny of ith sire |
| μ | = | The common mean |
| ai | = | The effect of the ith sire |
| eij | = | The uncontrolled environmental and genetic deviations attributable to the individuals. All error terms were random, normal and independent with expectation equal to zero |
RESULTS AND DISCUSSION
Table 1 shows the Least Square Means for plasma alkaline phosphatase and reproductive performance of cocks belonging to the two strains of Rhode Island chickens. All the parameters studied were higher in the cocks of strain B than those of strain A. In both strains, coefficient of variations was very high in the traits studied except for the activity of the enzyme.
The values of percentage fertility obtained in this study were within the range reported in literature. Singh and Belsare (1991) reported 83.80% for fertility in White Leghorn population; Chao and Lee (2001) reported 85 and 87% fertility in two experiments in Taiwan Country chickens. Szwaczkwoski et al. (2003) also reported 87.1 and 89.4% fertility in White Leghorn and New-Hampshire respectively. The percentage of eggs hatched also compares with the 43.32% reported by Singh and Belsare (1991) but lower than 75.5% reported by Szwaczkwoski et al. (2003). Differences in hatchability could be due to breed effects and or incubation environment.
Table 2 and 3 show genetic parameter estimates for plasma alkaline phosphatase activity and reproductive traits in strain A and B respectively while Table 4 and 5 show the environmental correlations between plasma alkaline phosphatase and reproductive traits in strain A and B, respectively.
| Table 1: | Least square means (±Standard Error) and coefficient of variation for plasma alkaline phosphatase activity and reproductive traits in strain A and strain B |
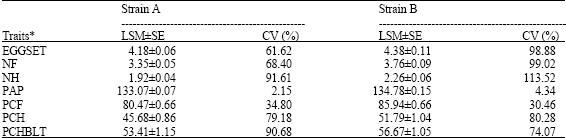 | |
| *: EGGSET = No. of egg set; NF = No. of fertile eggs; NH = No. of eggs hatched; PAP = Plasma Alkaline Phosphatase activity; PCF = % fertility eggs; PCH = % hatch; PCHBLT = % Hatchability of eggs | |
| Table 2: | Genetic parameter estimates for plasma alkaline phosphatase activity and reproductive traits in strain A** |
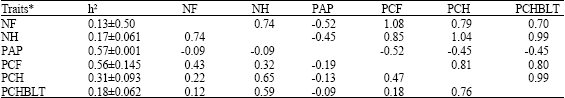 | |
| **: Above diagonal is genetic correlation and below the diagonal is phenotypic correlation. h2 = heritability | |
| Table 3: | Genetic parameter estimates for plasma alkaline phosphatase activity and reproductive traits in strain B** |
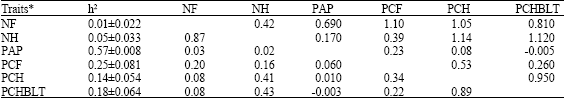 | |
| **: Above diagonal is genetic correlation and below the diagonal is phenotypic correlation. h2 = heritability | |
| Table 4: | Environmental correlations between reproductive traits in strain A |
 | |
| Table 5: | Environmental correlations between reproductive traits in strain B |
 | |
| *: EGGSET = No. of egg set; NF = No. of fertile eggs; NH = No. of eggs hatched; PAP = Plasma Alkaline Phosphatase activity; PCF = % fertility eggs; PCH = % hatch; PCHBLT = % Hatchability of eggs | |
High heritability estimates of enzyme activity obtained in this study agrees with earlier reports. Ferdoci et al. (1992) reported a high heritability estimate of (0.50±0.307). Obeidah et al. (1978) reported a high heritability estimate of 0.73 in Fayoumi layers while Zhon and Zhao (1991) reported a heritability value of 0.62 at 8 weeks of age in White Leghorn fowl. Matsumoto et al. (1960) also obtained an estimate as high as 0.85. High heritability of alkaline phosphatase activity suggests the possibility of changing the level of this enzyme at a rapid rate by selection (Ferdoci et al., 1992).
Low heritability estimates of reproductive traits confirm earlier reports (Kinney, 1969; Crittenden et al., 1957; Singh and Belsare 1991; Szwaczkwoski et al., 2003).
Moderate to high heritability estimates obtained for percentage fertility have been reported elsewhere. Chao and Lee (2001) reported heritability of fertility percentage of 0.459±0.297 in experiment 1 and 0.234±0.227 in experiment 2. The low heritability estimates for most of the reproductive traits could be due to non-additive genetic effects which are usually important in reproductive traits (Falconer and Mackay, 1996).
Negative correlations (genetic and phenotypic) between enzyme activity in cocks and reproductive traits in strain A agree with the reports of Veronina and Kurbatov (1971). They reported that fertility of eggs set and hatchability of fertile eggs were significantly higher in cocks with low enzyme activity.
McClung et al. (1972) however, reported that the genotypic correlations between alkaline phosphates and fertility were positive while between alkaline phosphates and hatchability was not determined. The observations on strain B seemed to follow this trend.
Positive genetic correlations between reproductive traits also confirm literature reports (Beaumont, 1992; Poivey et al., 2001). This is an indication of pleiotropic effect. Positive phenotypic correlations were observed among reproductive traits in both strains and environmental correlations follow the same trend as obtained for genetic correlation for each of the strains.
In conclusion, a high heritability estimate of the enzyme activity suggests that the level of this enzyme can be changed at a rapid rate by selection. And the high genetic correlation of the enzyme activity with reproductive traits in the cocks of strain A is an indication that the enzyme activity can be useful in selecting the cocks to improve fertility and or hatchability of eggs.
REFERENCES
- Beaumont, C., 1992. Genetic parameters of the duration of fertility in hens. Can. J. Anim. Sci., 72: 193-201.
CrossRefDirect Link - Chao, C.H. and Y.P. Lee, 2001. Relationship between reproductive performance and immunity in Taiwan. Country chickens. Poult. Sci., 80: 535-540.
Direct Link - Crittenden, L.B., B.B. Bohren and U.L. Anderson, 1957. Genetic variance and covariance of the components of hatchability in new hampshires1,2. Poult. Sci., 36: 90-103.
CrossRefDirect Link - Falconer, D.S. and T.F.C. Mackay, 1996. Introduction to Quantitative Genetics. 4th Edn., Prentice Hall, Harlow, England, ISBN-13: 9780582243026, Pages: 464.
Direct Link - Kind, P.R.N. and E.J. King, 1954. Estimation of plasma phosphatase by determination of hydrolysed phenol with amino-antipyrine. J. Clin. Pathol., 7: 322-326.
CrossRefPubMedDirect Link - McClung, M.R., H.M. Hyre and W.G. Martin, 1972. Two-way selection for serum alkaline phosphatase in laying hen. Poult. Sci., 51: 1428-1437.
CrossRefDirect Link - Moss, D.W., 1974. Multiple forms of alkaline phosphatate: Some topics of current interest. Histochem. J., 6: 353-360.
CrossRef - Poivey, J.P., Y.S. Cheng, R. Rouveir, C. Tai, C.T. Wang and H.L. Liu, 2001. Genetic parameters of reproductive traits in brown Tsaiya ducks artificially inseminated with semen from Muscovi drakes. Poult. Sci., 80: 703-709.
Direct Link - Syakalima, M., M. Takiguchi, J. Yasuda and A. Hashimoto, 1998. The canine alkaline phosphatases: A review of the Isoenzymes in serum, analytical methods and their diagnostic application. Jap. J. Vet. Res., 46: 3-11.
PubMed - Szwaczkwoski, T., K. Cywa-Benko and S. Wezyk, 2003. A note on inbreeding effects on productive and reproductive traits in laying hens. Anim. Sci. Paper Rep., 21: 121-129.
Direct Link