
N. Choudhary
Department of Genetics and Plant Breeding, CCS Haryana Agricultural University, Hisar, 125004, India
U. Ahuja
Department of Genetics and Plant Breeding, CCS Haryana Agricultural University, Hisar, 125004, India
V. Chawla
Department of Genetics and Plant Breeding, CCS Haryana Agricultural University, Hisar, 125004, India
R.K. Jain
Department of Genetics and Plant Breeding, CCS Haryana Agricultural University, Hisar, 125004, India
P. Kumari
Department of Genetics and Plant Breeding, CCS Haryana Agricultural University, Hisar, 125004, India
K.R. Batan
Department of Genetics and Plant Breeding, CCS Haryana Agricultural University, Hisar, 125004, India
Asian Journal of Agricultural Research
Year: 2011 | Volume: 5 | Issue: 5 | Page No.: 250-259







ABSTRACT
Weedy rices commonly known as ‘red rices’, infest rice fields worldwide. They compete with rice for nutrients, deteriorate quality and are difficult to eradicate. Understanding their genetic diversity and the origin may prove helpful in designing an effective way for managing them. A collection of weedy rices from Haryana were evaluated along with cultivated and wild rices for variability and diversity at morphological and molecular level using 35 ISSR primers. Significant variation was observed at morphological and genetic level in weedy rices of Haryana. Weedy rices were found to be distinct from white and red type cultivated rices and wild strains. Twenty one ISSR markers gave amplification of DNA and a total of 91 polymorphic bands were found. With 9 ISSR markers 20 unique bands were observed in 17 genotypes. Markers such as Pr814, Pr836 and Pr858 produced unique bands in weedy rices only. Alleles unique to weedy rices observed by ISSR markers could be converted into SCAR markers for identification. Weedy rices were placed alone as well as with other groups in cluster analysis. Both short and long grain weedy rices were found in collection though at present only long grain types are cultivated indicating that leftover progenies of local cultivars segregated into various plant types giving rise to the weedy rice.
PDF Abstract XML References Citation
Received: August 18, 2011;
Accepted: November 02, 2011;
Published: December 16, 2011
How to cite this article
N. Choudhary, U. Ahuja, V. Chawla, R.K. Jain, P. Kumari and K.R. Batan, 2011. Morphological and Molecular Variability in Weedy Rices of Haryana. Asian Journal of Agricultural Research, 5: 250-259.
DOI: 10.3923/ajar.2011.250.259
URL: https://scialert.net/abstract/?doi=ajar.2011.250.259
DOI: 10.3923/ajar.2011.250.259
URL: https://scialert.net/abstract/?doi=ajar.2011.250.259
INTRODUCTION
Rice is the most important cereal crop. The plant has wild, cultivated and weedy forms. Off type rice plants with red seeds are considered weedy (Oka, 1988) and color of bran layer of red rice’s varies from light to dark red. Weedy and cultivated rice are taxonomically classified as same species as (O. sativa L.). Weedy rice shares traits with both cultivated and wild types of rice. It is thought to have evolved either by hybridization between wild and cultivated rice, or as a transitional form changing from wild to cultivated types, a relic or primitive cultivar, or an off type derived from the present cultivars (Tang and Morishima, 1988). Earlier to advent of high yielding white varieties, red rices formed an important group in rice growing Asian countries because of their tolerance to adverse conditions and high Zn and iron content (Ahuja et al., 2007) but weedy rice competes with rice for space, nutrients and has higher N use efficiency for biomass production and poses a number of problems (Burgos et al., 2006).
The close similarity between weedy rice and cultivated rice has prevented use of selective herbicides (Hoagland and Paul, 1978). Also concern exists for the danger of development of more aggressive weed through crop allele escape (Chen et al., 2004). In field, U. S. growers spend much energy and money in fighting red rices termed as ‘Red Menace’. Costly Color sorters are installed to separate them from milled product (Rood, 2000). The Asians are somewhat tolerant towards red weedy rices as they eat and assign special names for them e.g., Sharei in Korea and Ludao in Japan (Ahuja et al., 2007).
In recent past, weedy rices have been used for various types of studies as variability, evolution of rice, gene flow, gene cloning, introgression and QTL analysis of low temperature tolerance, inheritance of seed dormancy, shedding and pollen sterility (Suh and Ha, 1987; Tang and Morishima, 1988; Vaughan et al., 1995; Suh et al., 1999; Ahn et al., 2002; Jing et al., 2003; Gealy et al., 2009). Understanding the origin of weedy rice may be helpful in designing an effective way for their manage (Pyon et al., 2000) and they have been utilized in rice varietal improvement programmes to impart resistance against many diseases and pests. Haryana state is known for cultivation of scented and fine grain rices. At present white, medium to long grain varieties are grown and no red cultivated or wild variety is known. Rice fields in Haryana are often contaminated by red colored weedy rice called choba. Color sorters are used to separate red grains from milled product in quality rices (Ahuja et al., 2007). A collection of weedy rices from Haryana were evaluated for variability at morphological and molecular level along with cultivated and wild rices to investigate its lineage.
MATERIALS AND METHODS
Plant material: As many as 15 weedy entries were collected from Kaul and Sonipat areas of Haryana. Weedy plants were distinct from cultivated rice by height, husk color and awns. Two cultivated white strains were obtained from Rice Research Station, Kaul whereas 11 red cultivated and 2 wild from P.A.U Ludhiana.
Morphological variability: Wild entries were grown in pots and the rest 28 rice genotypes were sown at Rice Research Station, Kaul in a dry bed nursery. Seedlings were transplanted after 30 days in randomized block design with three replications following standard package and practices. At maturity data were recorded for plant height, panicle length, grain yield per plant, 1000-grain weight, hull color, grain color, awning and grain dimensions (paddy length, breadth and thickness) on 10 plants per replication, selected randomly following Standard Evaluation System of International Rice Research Institute (IRRI, 1996).
Molecular variability: Genomic DNA was isolated from leaf samples of 60 day old rice plants using CTAB method as described by (Saghai-Maroof et al., 1984). The quantity and quality of DNA samples were checked by 0.8% agarose gel electrophoresis (Sambrook et al., 1989) using a standard containing 100 ng μL-1 genomic λ DNA.
A total of 35 ISSR primers (UBC primer set # 9, John Hobbs, NAPS Unit, University of British Columbia, Vancouver, V6T 1Z3 Canada) were used in present study. The PCR reaction was carried out using a single primer at a time in 10 μL reaction mix containing 12.5 ng of template DNA, 1xPCR buffer, 2.5 mM MgCl2, 500 μM dNTP mix, 0.3 μM primer and 1.5 unit Taq DNA polymerase. PCR amplifications were performed in 96-well plates on a PTC100TM 96V thermocycler (MJ Research, Watertown, MA, U.S.A.) with initial denaturation at 94°C for 4 min; followed by 45 amplification cycles of denaturing at 94°C for 1 min, annealing at 50°C for few primers and 55°C for rest for 1 min; extension at 72°C for 2 min and final extension at 72°C for 15 min. The PCR products were resolved by electrophoresis on 1.5% (w/v) agarose gel. They were viewed under UV light (350 nm) and photographed using Vilber Lourmat gel documentation system. Molecular weight of bands was estimated using a wide range ladder (100 bp) from Fermentas, India.
Statistical analysis: Based on morphological data mean performance and variance was analyzed among all genotypes and grouping was done using Ward’s minimum variance method using software Windostat version 7.0. Intra and inter-cluster Euclidean distances generated were used to describe the relationships among the rice genotypes. Euclidean distance [d (i,j)] between two individuals i and j, having observations on characters (p) denoted by x1, x2,…,xp and y1, y2…., yp for i and j, respectively, can be calculated as under:
d (i,j) = [(x1 – y1)2+(x2-y2)2+…(xp-yp)2]1/2
Based on presence/absence of band, genetic similarity was evaluated by calculating the Jaccard similarity coefficient to estimate all pair wise differences in the amplification products produced by the primers for all rice genotypes. The 0/1 matrix was used to calculate the similarity/genetic distance using ‘simqual’ sub-program of NTSYS-PC version 2.02 software package (Rohlf, 1990). The similarity matrix thus obtained was subjected to cluster tree and principal component analyses using the Un-weighted Pair-group Method with an Arithmetic Average (UPGMA) sub-program of NTSYS-PC.
RESULTS AND DISCUSSION
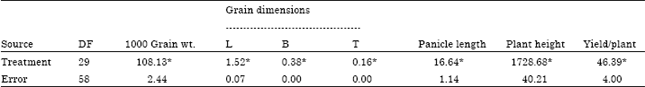
Variation for morphological traits: Analysis of variance showed significant differences among genotypes for all the characters under study (Table 1).
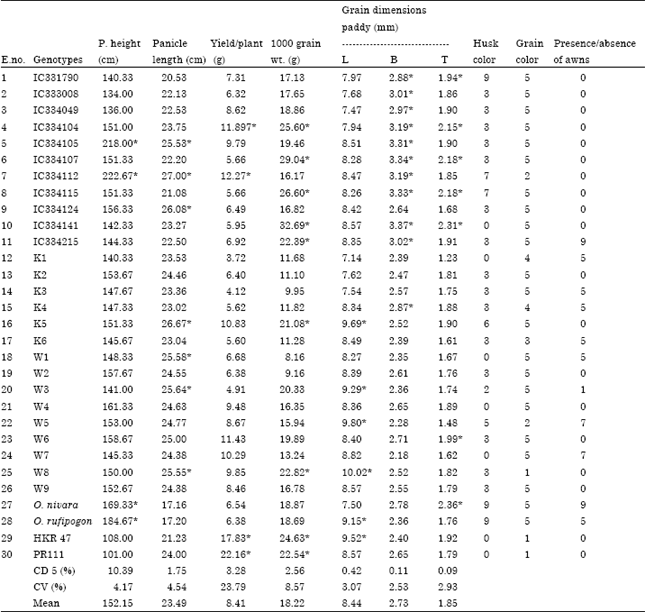
Agronomic traits: The plant height ranged from 101 cm (PR111) to 222.67 cm (IC334112). All wild, red cultivated and weedy rices were tall whereas both cultivated white entries were semi-dwarf in stature. The panicle length ranged from 27 cm (IC334112)-17.16 cm (O. nivara). The yield per plant ranged from 22.16 (HKR47)-3.72 g (K1). Yield per plant was highest in cultivated white followed by red cultivated, weedy rice and wild types. Grain weight ranged from 32.69 (IC334141)-8.16 g (W1) (Table 2).
Cultivated white and cultivated red genotypes were awn less. Both wild entries possessed awns however, awns of different length were found in weedy rice. Hull color varied from straw to black. Both the wild possessed black hulls and both cultivated white entries possessed straw hulls. However various shades of hull colors were observed in cultivated red and weedy rices. In weedy rice husk color ranged from straw, brown spots, brown furrows, reddish to light purple and purple spots (scale 0-9). Grain color ranged from white to red. Weedy rices showed different shades from white to light brown, brown, speckled red and red (scale 0-7). Interestingly in entry W3, red, pink and white grains coexisted in a single panicle (Table 2).
Cluster analysis for morphological data: Dendrogram was constructed based on data collected from all the rice genotypes for all characters mentioned above using Ward's Minimum Variance method.
| Table 1: | Analysis of variance for morphological traits of wild, red and weedy rices |
 | |
| *Mean values are significant at 5% level of significance | |
| Table 2: | Mean performance of different rice genotypes for different characters |
 | |
| Red cultivated 1-11, weedy Kaul, 12-17, Sonipat 18-26, Wild 27-28, White cultivated 29-30, *Mean values are significant at 5% level of significance | |
Five clusters were obtained. Number 1-30 correspond to serial number of rice genotypes as mentioned in (Table 3). Cluster I included 10 entries (8 cultivated red, 2 wild). Cluster II included 2 (both of cultivated white). Cluster IV included 7(weedy red). Cluster V included 9 (one cultivated red, 8 weedy red).
Euclidean intra-cluster distances showed a range of 1.14 to 3.01. Minimum intra-cluster distance (1.14) was within cluster III and maximum (3.01) was within cluster I. Euclidean inter-cluster distances showed a range of 2.99 to 6.18 (Fig. 1, 2).
ISSR markers based polymorphism among rice genotypes: Out of 35, only 21 ISSR markers gave amplification. A total of 95 bands were detected out of which 4 were monomorphic and 91 were polymorphic.
| Table 3: | Cluster information using ward’s minimum variance |
 | |
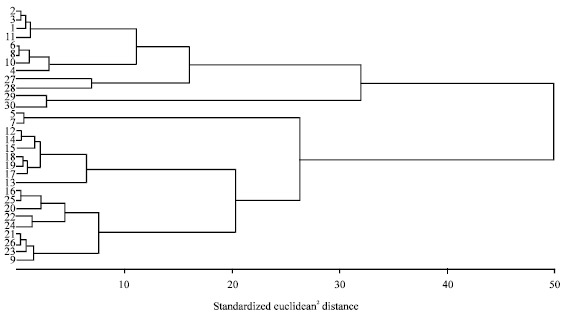 | |
| Fig. 1: | Ward's Minimum Variance Dendrogram based on agronomic data of all accessions |
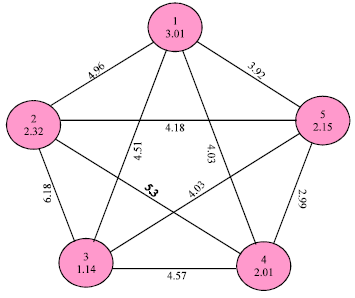 | |
| Fig. 2: | Inter and intra cluster distance among all accessions |
ISSR DNA bands varied between 1 (primer IS21) and 14 (UBC primer No. 845) with an average of 4.5 bands per primer. Maximum number of polymorphic bands (14) was obtained using UBC primer No. 845; the average number of polymorphic bands was 4.3 per primer. The polymorphism percentage ranged from 0-100% with an average polymorphism of 88.9% across all the varieties. The size of PCR amplified products ranged between 200 and 1300 bp. UBC No. 854, IS21, IS99 and IS121 gave monomorphic bands whereas UBC No. 809, 811 814, 822, 835, 836, 845, 852, 856, 858, 873 and IS29, IS43, IS69, IS88 IS90 and IS116 gave polymorphic bands.
Genetic diversity analysis: ISSR marker analysis generated high levels of molecular polymorphism within this selection of cultivated red and weedy rices (Fig. 3). Dendrogram obtained using the ISSR allelic diversity data shows that at 0.46 similarity coefficient the 30 rice genotypes could be divided into five groups (Table 4). All the cultivated and weedy rice selections were found to be genetically distinct from one another except W5 and W6 which has a high similarity coefficient of 0.77. Though number of indica rice varieties included in this study were low but the study do suggest huge diversity among the indica and cultivated red and weedy rice genotypes. Notably, indica rice variety, HKR47, was relatively closer to the wild ancestors, O. rufipogon and O. nivara, compared to cultivated red and weedy rice genotypes.
Unique alleles: A total of 19 unique alleles were detected in a total of 17 genotypes with 9 ISSR primers. These unique bands ranged from 1-4 with an average of 2.22.
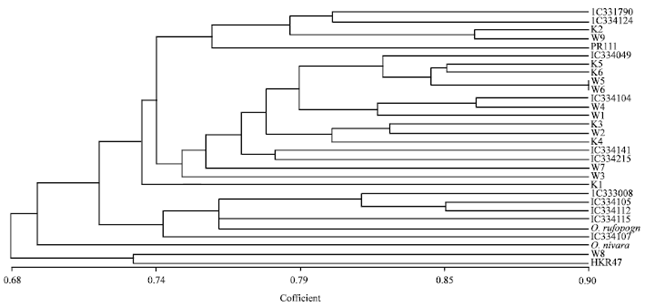 | |
| Fig. 3: | Dendrogram based on ISSR Polymorphisms of all 30 genotypes by UPGMA cluster analysis |
| Table 4: | UPGMA cluster analysis of weedy, wild and cultivated rice accessions |
 | |
| Table 5: | Unique alleles obtained using 9 ISSR primers in 17 rice genotypes |
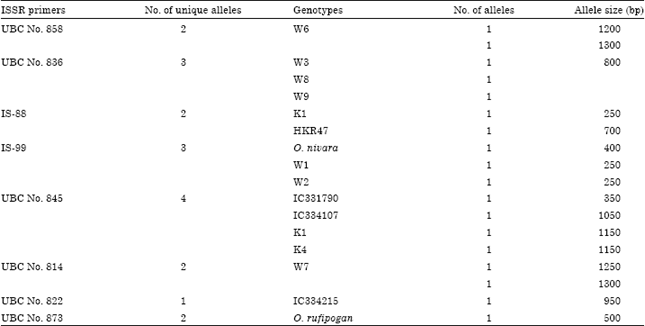 | |
The size of unique bands ranged from 250-1300 bp (Table 5). UBC primer No. 814, 836 and 858 produced unique bands in weedy rices only.
A collection of weedy rices of Haryana showed wide variation in morphological traits. Weedy rices were tall and poor yielded. Two weedy entries (K5 and W8) possessed grain weight near cultivated rices whereas, the seven entries possessed lowest 1000 grain weight (8-12 g) among all the entries under test. The lighter grain weight in weedy in comparison with cultivated rice is in accordance with earlier reports (Tang and Morishima, 1989). Weedy entry, W8 possessed grains longer (10.02 mm) than local Basmati rice (9.81 mm). Four weedy rices could be classified under short grain category. At present in Haryana only long grain types are cultivated while short grain types were grown some 50 years back (Ahuja et al., 1995). Therefore, short grain weedy rices seem to be progeny of volunteer plants of old cultivars (Zhang et al., 2008). Weedy rices possessed thin grains and the grain thickness was lower than that of cultivated rices. Paddy grain dimensions (length, breadth and thickness) in Indianrices ranges from 5.15-11.27, 1.97-3.73 and 1.61-2.57 mm (Hector et al., 1933; Ramaiah and Rao, 1953).
The husk color of weedy rices was found to be between cultivated rices and wild i.e., varied from straw color (4 out of 15) to various shades of brown. Eight out of 15 weedy rices possessed awns and 14 out of 15 red grains. Red, pink and white grains in a single panicle of W3 might be the result of crossing between weedy and cultivated or between weedy and weedy. Gealy et al. (2009) showed evidence of hybridization of weedy rice with cultivated rice by DNA studies. Different hull colors were reported by (Shivrain et al., 2010) in weedy rices of Arkansas. Federici et al. (2001) reported weedy rices with straw hull and no awn mimicking cultivated as well as wild types. It is observed that weedy rices are distinct from cultivated rices (both red and white) in awning, husk color, grain color, grain weight and thickness. Similar variation in grain traits was reported by Suh and Ha (1987) and Vaughan et al. (1995).
In the present study, both the cultivated white rice genotypes are placed in extremes groups by molecular marker analysis though were placed in same cluster by Euclidean square analysis.
Similar is the case with wild rice and other groups. Similar results i.e., variability at morphological level did not coincided with diversity at molecular level were reported by Vanaja et al. (2007) in rice, Bandyopadhyay et al. (2007) in finger millets and Devanshi et al. (2007) in Ber. In the present study Oryza nivara and rufipogon are placed in different cluster. Similar results were reported by Sujatha et al. (2004). Ryza rufipogon is clustered along with f our cultivated red rices in cluster 4. It is thought to be the progenitor species of the cultivated rice, O. sativa L. (Oka, 1988).
The genetic clustering of weedy rice accessions from Haryana was found to be random with respect to location as samples from one location did not belong to the same genotypic cluster. Most of the weedy rices were placed with cultivated white, red and wild rices except K1 which was placed in separate cluster. UBC primers No. 814, 836 and 858 produced unique bands in weedy rices which indicate that these ISSR primers may be used to distinguish cultivated and weedy rice varieties at molecular level. However, this will require converting them into SCAR markers. Gealy et al. (2009) reported SSR marker loci which could distinguish between U.S. weedy red rice and white cultivars. Chen et al. (2004) reported frequent occurrence of gene flow from cultivated rice to weedy rices and according to Yu et al. (2005) unique alleles in weedy rices are through gene flow.
Possible origin of weedy rices have been proposed as segregating progenies of cultivated rice (Suh et al., 1999), intervarietal hybridization or mutation (Cao et al., 2006), hybridization of discrete types of rice cultivars generating partially sterile plants resulting into weedy strains (Ishikawa et al., 2005) or parallel evolution, hybridization, gene flow etc (Kane and Baack, 2007). Watanabe et al. (2000) reported that different rice growing locations show different patterns of genetic diversity, depending on specific combinations of germplasm from which weedy rices originated. Present material comprises weedy strains genetically close to red and white cultivated strains, wild strains and distinct from all three groups indicates that the weedy rices of Haryana might have originated by local cultivars leaving volunteer plants through the succeeding cropping seasons, hybridization of discrete types, segregating progenies of cultivated rice and gene flow as proposed by various authors.
CONCLUSION
From the above study it is concluded that weedy rices possess genes useful under adverse environmental conditions are genetically more similar to cultivated rice than wild rices but with the help of some ISSR primers weedy rices are isolated from cultivated and wild rice and they are easily eliminated from the rice field. This was a small sample collected from two districts of Haryana and variability obtained shows that there is need to collect and evaluate more weedy rices having important genes.
REFERENCES
- Burgos, N.R., R.J. Norman, D.R. Gealy and H. Black, 2006. Competitive N uptake between rice and weedy rice. Field Crops Res., 99: 96-105.
Direct Link - Cao, Q.J., B.R. Lu, H. Xia, J. Rong, F. Sala, A. Spada and F. Grassi, 2006. Genetic Diversity and origin of weedy rice (Oryza sativa f. spontanea) populations found in North-Eastern China revealed by Simple Sequence Repeat (SSR) markers. Ann. Bot., 98: 1241-1252.
CrossRef - Chen, L.J., D.S. Lee, Z.P. Song, H.S. Suh and B.R. Lu, 2004. Gene flow from cultivated rice (Oryza sativa) to its weedy and wild relatives. Ann. Bot., 93: 67-73.
Direct Link - Gealy, D.R., H.A. Agrama and G.C. Eizenga, 2009. Exploring genetic and spatial structure of U.S. weedy red rice (Oryza sativa) in relation to rice relatives worldwide. Weed Sci., 57: 627-643.
Direct Link - Hoagland, R.E. and R.N. Paul, 1978. A comparative SEM study of red rice and several commercial rice (Oryza sativa) varieties. Weed Sci., 26: 619-625.
Direct Link - Ishikawa, R., N. Toki, K. Imai, Y.I. Sato and H. Yamagishi et al., 2005. Origin of weedy rice grown in Bhutan and the force of genetic diversity. Genet. Resour. Crop Evol., 52: 395-403.
CrossRefDirect Link - Jing, W., L.L. Liu, L. Jiang and J.M. Wan, 2003. Mapping of S33(t) and S34(t) for pollen sterility in hybrids between a weedy strain and a japonica variety in rice. Rice Genet. Newslett., 23: 16-19.
Direct Link - Saghai-Maroof, M.A., K.M. Soliman, R.A. Jorgensen and R.W. Allard, 1984. Ribosomal DNA spacer-length polymorphisms in barly: Mendelian inheritance, chromosomal location, and population dynamics. Proc. Natl. Acad. Sci., 81: 8014-8018.
CrossRefDirect Link - Suh, J.P., S.N. Ahn, H.P. Moon and H.S. Suh, 1999. QTL analysis of low temperature germinability in a Korean weedy rice (Oryza sativa L.). Rice Genet. Newslett., 16: 53-55.
Direct Link - Tang, L.H. and H. Morishima, 1989. Variation and inheritance of seed shedding in weedy rice. Rice Genet. Newslett., 6: 72-72.
Direct Link - Vaughan, D.A., H. Watanabe, A. Zain and K. Okuno, 1995. Genetic diversity of weedy rice in Malaysia. Rice Genet. Newslett., 12: 176-176.
Direct Link - Watanabe, H., D.A. Vaughan and N. Tomooka, 2000. Weedy Rice Complexes: Case Studies from Malaysia, Vietnam and Surinam. In: Wild and Weedy Rice in Rice Ecosystems in Asia: A Review, Chin, D.V., B. Baker and M. Mortimar (Eds.). International Rice Research Institute, Los Banos, Philippines, pp: 25-34.
- Yu, G.Q., Y. Bao, C.H. Shi, C.Q. Dong and S. Ge, 2005. Genetic diversity and population differentiation of Liaoning weedy rice detected by RAPD and SSR markers. Biochem. Genet., 43: 261-270.
CrossRefPubMedDirect Link - Zhang, J., N.R. Burgos, M.A. Kun, Y.J. Zhou, R.M. Geng and L.Q. Yu, 2008. Genetic diversity and relationship of weedy rice in Taizhou City, Jiangsu Province, China. Rice Sci., 15: 295-302.
CrossRef - Vanaja, T., G.J. Randhawa, R. Singh and T. Mohapatra, 2007. Analysis of molecular diversity and differentiation of photoperiod sensitive and insensitive rice varieties. Indian J. Genet. Plant Breed., 67: 128-134.
Direct Link - Sujatha, K., R. Upadhyay, K. Kaladhar, N.S. Rani and N. Sarla, 2004. Genetic relationship among aromatic short grain and Basmati rice based on ISSR and SSR markers. Rice Genet. Newslett., 21: 24-25.
Direct Link - Federici, M.T., D. Vaughan, N. Tomooka, A. Kaga and X.W. Wang et al., 2001. Analysis of uruguayan weedy rice genetic diversity using AFLP molecular markers. Analysis of Uruguayan weedy rice genetic diversity using AFLP molecular markers. Electron. J. Biotechnol.
Direct Link