
Claver Ngaboyisonga
Department of Plant Science and Crop Protection, Faculty of Agriculture, University of Nairobi, P.O. Box 29053-00625, Nairobi, Kenya
Kiarie Njoroge
Department of Plant Science and Crop Protection, Faculty of Agriculture, University of Nairobi, P.O. Box 29053-00625, Nairobi, Kenya
Duncan Kirubi
Department of Plant Science and Crop Protection, Faculty of Agriculture, University of Nairobi, P.O. Box 29053-00625, Nairobi, Kenya
Sam M. Githiri
Department of Plant Science and Crop Protection, Faculty of Agriculture, University of Nairobi, P.O. Box 29053-00625, Nairobi, Kenya
Asian Journal of Agricultural Research
Year: 2009 | Volume: 3 | Issue: 1 | Page No.: 1-10







ABSTRACT
The aim of this study was to assess how low nitrogen and drought stresses affect genetic parameters of grain yield and endosperm hardness of QPM. Twelve inbred lines were acquired from CIMMYT and used to generate 36 single cross hybrids with North Carolina Design II procedures. The single crosses were evaluated at Kiboko in Kenya and Rubona in Rwanda in 2005 and 2006 cropping seasons under optimum, low nitrogen and drought conditions. Observations were performed on grain yield and endosperm hardness. The results showed that gene action on the grain yield was predominantly non-additive and maternal, whereas that of the endosperm hardness was predominantly additive and maternal. Low nitrogen and drought changed the proportions of different genetic effects. They changed as well the magnitudes and direction of General Combining Abilities (GCAs) of lines and Specific Combining Abilities (SCAs) of crosses. It was possible to have together, significant and positive SCAs for grain yield and significant and negative SCAs for endosperm hardness. Crosses with significant and positive SCAs for grain yield and significant and negative SCAs for endosperm hardness are candidates to use in QPM production, especially in drought and low nitrogen prone areas.
PDF Abstract XML References Citation
How to cite this article
Claver Ngaboyisonga, Kiarie Njoroge, Duncan Kirubi and Sam M. Githiri, 2009. Effects of Low Nitrogen and Drought on Genetic Parameters of Grain Yield and Endosperm Hardness of Quality Protein Maize. Asian Journal of Agricultural Research, 3: 1-10.
DOI: 10.3923/ajar.2009.1.10
URL: https://scialert.net/abstract/?doi=ajar.2009.1.10
DOI: 10.3923/ajar.2009.1.10
URL: https://scialert.net/abstract/?doi=ajar.2009.1.10
INTRODUCTION
Maize is an important source of protein in human lives (Anonymous, 1988). Nutritionally, maize is deficient in two essential amino acids: lysine and tryptophan (Krivanek et al., 2007). The hope that breeders could improve the protein quality of maize goes back to the discovery of high lysine and tryptophan endosperm mutants: opaque-2 (o2) and floury-2(fl2) in 1960s. The mutants could raise about two to three folds the amount of lysine and tryptophan compared to normal maize and hence represented a valuable tool to correct maize deficiency in protein quality (Krivanek et al., 2007; Prasanna et al., 2001; Vasal, 2001).
Breeders and other scientists transferred o2 gene to normal maize and distributed O2-maize to the farmers but very soon, the research on o2 was totally abandoned except at CIMMYT, because o2 maize had unacceptable kernels, was very susceptible to ear rot and had low yield (Vasal, 2000).
It was seen that genetic modifiers could alter the endosperm of o2-maize by suppressing bad effects created by the mutant and by converting soft, chalky and opaque endosperm into normal looking, hard and translucent endosperm (Krivanek et al., 2007; Prasanna et al., 2001; Vasal, 2001).
Therefore, CIMMYT continued to work on Quality Protein Maize (QPM), combining the genetic systems of o2 and endosperm modifiers and using a multidisciplinary team of scientists. After, 10 years, in early 1980s, CIMMYT produced new types of maize that combined nutritional excellence of o2 maize with kernel structure and the grain yield of conventional maize varieties. These types of genotypes were called QPM (Vasal, 2001; Prasanna et al., 2001).
The genetic system of o2 gene is qualitative. However, because it is recessive, its effects are expressed in the endosperm when three alleles, two from female and one from male parent are present. It increases lysine and tryptophan in endosperm by acting on the four types of storage proteins in maize endosperm: albumins, globulins, zeins and glutelins. Zeins contain low lysine with 0.1 g/100 g while glutelins are considerably rich in lysine with 2 g/100 g or more (Lin et al., 1997). The o2 mutant increases the level of lysine and tryptophan by suppressing or reducing the synthesis of zeins and increasing that of glutelins (Ferreira et al., 2005; Azevedo et al., 2003).
The o2 gene adversely affects several important agronomic traits including kernel characteristics. It adversely affects the accumulation of dry matter resulting in lower yields due to increased endosperm size. The kernel phenotype is changed in a soft, chalk and dull appearance. Kernels dry slowly following physiological maturity of the grain and have a higher incidence of ear rots. Other changes include larger germ size and low kernel density (Ferreira et al., 2005; Lin et al., 1997).
Modifiers are genes capable of altering the expression of other genes at different loci in the genome (Thain and Hickman, 2003). The genetic modifiers alter the undesirable correlated effects of o2 gene. The parties of the endosperm modified are vitreous and hard instead of being opaque and soft. Several reports indicated that modified endosperm texture is polygenically controlled with additive type of genetic variation playing an important role (Prasanna et al., 2001; Vasal, 2000, 2001).
Current effort on QPM is to increase its cultivation in the regions, especially in Sub-Saharan Africa, experiencing problems of malnutrition and where maize is the staple crop. In these regions, however, maize is frequently produced under environmental stresses, among which, low soil nitrogen and drought are the most important. Impacts of low nitrogen and drought on grain yield of normal maize have been extensively studied (Edmeades et al., 2006). However, impacts on genetic variation of grain yield, particularly on endosperm hardness of QPM have not yet been studied at any extent. The objective of this study was to asses the impacts of low nitrogen and drought on genetic parameters of grain yield and endosperm hardness of Quality Protein Maize. Specifically, this study was to assess gene action governing grain yield and endosperm hardness under optimum and how it is changed under low nitrogen and drought. Furthermore, it was to assess how those stresses affect combining abilities of grain yield and endosperm hardness.
MATERIALS AND METHODS
Twelve CIMMYT-Kenya inbred lines (Table 1) were used to produce 36 single cross hybrids with North Carolina Design II (NCDII) procedures during the season: October 2004-February 2005. The 36 single crosses were evaluated at KARI-Kiboko station (2°25 S, 37°75 E, 975 masl) in Kenya in 2005 (March-August 2005) and in 2006 (October 2005- February 2006) under optimum, low nitrogen and drought environments. They were also evaluated at ISAR-Rubona station (2°29 S, 29°46 E, 1650 masl) in Rwanda in 2005 (March-August 2005) under optimum and low nitrogen environments, as drought conditions were not possible in that site.
| Table 1: | Pedigrees of limes crossed in North Carolina Design II |
 | |
The optimal environments at Kiboko received water by sprinkler irrigation throughout the cropping season. The irrigation was performed two times a week for a duration of three hours each time. Fertilizers were applied by supplying 64 kg N ha-1 and 46 P ha-1 at planting, 46 kg N ha-1 4 weeks after planting and 46 kg ha-1 seven weeks after planting. At Rubona, the optimal environments were achieved by applying 51 kg ha-1 N, 51 kg ha-1 P2O5 and 51 kg ha-1 K2O before planting and 6 weeks after planting, 46 kg ha-1 N. Water was supplied by rain, as Rubona site did not have irrigation facilities.
The low nitrogen environments were achieved at Kiboko by not top-dressing nitrogen fertilizers during the season. However, a starter nitrogen of 18 kg ha-1 was applied at planting to allow uniform germination, emergence and early seedling growth. Phosphorus was applied at 46 kg ha-1 at planting while irrigation was provided during the cropping season. Field was thoroughly cleaned during plowing and all plant residues removed. Depleting nitrogen in the field achieved low nitrogen condition at Rubona. The depletion process followed the procedures described by Banzinger et al. (2000). Water was supplied by natural rain rainfall. During planting little nitrogen at a rate of 9 kg ha-1 was supplied. Drought environments were obtained at Kiboko by stopping irrigation one week before flowering. The field received 64 kg N ha-1 and 46 P ha-1 at planting, 46 kg N ha-1 4 weeks after planting and 46 kg ha-1 seven weeks after planting like optimum environment.
The experimental design was an incomplete block design (alpha-lattice) with three replications. The plot was made of two rows of 5 m length with the distance between rows and hills of 0.75 and 0.25 m, respectively. Sowing was performed by seeds per hill and a thinning 3 weeks after planting reduced the stand at one plant per hill. Thus, a planting density of 53000 plants ha-1 was achieved.
At Kiboko, before planting, Furadan® 5G (composition: 5% w/w carbonfuran, 10% inert) was applied in rows and covered with little soil to control soil, germination and seedling pests. Additionally, an insecticide called Buldock was applied two times: three weeks after planting and 6 weeks after planting to control stem borers that are the major biological constraint to maize at Kiboko. Weeding in both sites was performed, as it was required.
The traits observed were yield and endosperm modification. The plot grain yields were obtained by weighting the total ears harvested (fresh weight in kg, FW) and sampling kernels to obtain grain moisture GM (%) using a portable moisture meter. The ears were dried to a constant weight (dry weight in kg, DW) and then shelled to obtain grain weight GW (kg). Taking A as the distance (m) between rows and B the distance (m) between hills at planting, C the length (m) of harvested rows and D the number of rows harvested, grain yield (GYD) in t/ha at 15% of grain moisture was obtained by the formula:
Endosperm hardness scores were recorded following the methodology of Pixley and Bjarnason (2002). Ten best ears in each plot were identified and 10 kernels were taken in the middle of the ear to make 100 kernels for each plot. The 100 kernels were thereafter sorted and classified into five classes of endosperm hardness using a light table. The scores were based on appearance of kernel endosperm on the light table. The class 1 was made of kernels 100% modified kernels and looked like those of normal maize. Classes 2, 3 and 4 were defined as follow: class 2: 75% hard and translucent and 25% soft and opaque; class 3: 50 % hard and translucent and 50% soft and opaque; class 4: 25% hard and translucent and 75% soft and opaque. The class 5 comprised kernels that were 100% soft and opaque (Fig. 1). Taking A as the number of kernels in class 1, B in class 2, C in class 3, D in class 4 and E in class 5, the endosperm hardness score of a plot was obtained by the formula:
 | |
| Fig. 1: | The five classes of endosperm hardness revealed by a light table: Class 1 = 100% hard, Class 2 = 75% hard, Class 3 = 50% hard, Class 4 = 25%, Class 5 = 100% soft |
The genetic analysis was performed in each environment, across combined environments of the same kind (same condition) and across all eight environments following the model of a cross-classification design where the sources of variation of crosses were subdivided into those of female parents (female additive effects), male parents (male additive effects) and femalesxmales (non-additive).
RESULTS
Grain Yield
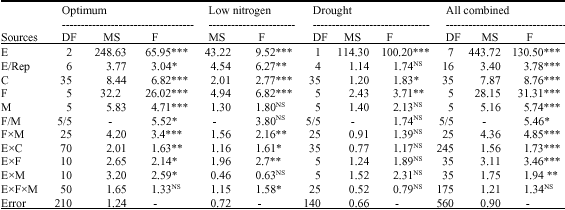
Combined Analysis of Variance (ANOVA) across optimum environments indicated highly significant differences (p<0.001) between crosses implying presence of highly significant genetic variation. Female and male parents were highly significantly different (p<0.001) implying that both female and male additive effects were highly significant. The interaction femalesxmales was highly significant (p<0.001) denoting that non-additive effects were highly significant. F = MSF/MSM showed the significance of maternal effects (p<0.05) (Table 3).
The interaction crossesxenvironments was highly significant (p<0.01) revealing that genetic variation highly interacted with environments. Besides, interactions femalesxenvironments and malesx environments were significant (p<0.05) while the interaction femalexmalesxenvironment was not, signifying that female and male additive effects interacted with environments whereas non-additive effects did not (Table 2).
| Table 2: | Combined ANOVA for combining abilities across optimum, low nitrogen and drought for grain yield (t ha-1) |
 | |
| ***Significance at p<0.001, **Significance at p<0.01, *Significance at p<0.05, NS: No Significance (p>0.05), Rep: Replications, C: Crosses F: Female parents, M: Male parents, SS = MSxDF | |
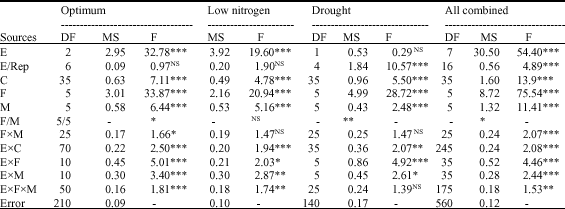
| Table 3: | Combined ANOVA for combining abilities across optimum, low nitrogen and drought for endosperm hardness scores (1-5) |
 | |
| ***Significance at p<0.001, **Significance at p<0.01, *Significance at p<0.05, NS: No Significance (p>0.05), Rep: Replications, C: Crosses F: Female parents, M: Male parents, SS = MSxDF | |
The proportion of female additive effects (SSF/SSC) was 54.5% and the fraction of male additive effects (SSM/SSC) was 9.9 %, thus, total additive effects [(SSF+SSM)/SSC] explained 64.4 % of genetic effects. Maternal effects, estimated as the difference between female and male additive effects [(SSF-SSM)/SSC] constituted 44.7% of genetic effects. Total additive effects comprised 44.7% of maternal effects and 19.8% of additive effects (2SSM/SSC). Non-additive effects formed 35.6% of genetic effects. Therefore, under optimum environments, genetic effects on grain yield comprised 19.8% of additive effects, 44.7% of maternal effects and 35.6% of non-additive effects demonstrating that additive, non-additive and maternal effects controlled grain, but maternal effects and non-additive effects were predominant over non-additive effects because they explained 80.3% of genetic variation. Combined ANOVA across low nitrogen environments showed highly significant differences (p<0.001) between crosses indicating significant genetic variation. Female additive effects were highly significant (p<0.001), but male additive effects were not significantly different (p>0.05) implying that additive effects were made essentially of maternal effects. The interaction femalesxmales was highly significant (p<0.01) denoting that non-additive effects were highly significant. The interaction Crossesx Environments being significant indicated that genetic variation interacted with environments. Female additive and non-additive effects interacted with environments while male additive effects did not (Table 2).
Female additive effects accounted for 35.2% of genetic effects and male additive effects constituted 9.2% both making total additive effects to constitute 44.4% of genetic variation. Non-additive effects accounted for 55.6% of genetic variation. Total additive effects comprised 25.9% of maternal effects and 18.5% of additive effects. Therefore, under low nitrogen environments, genetic effects on grain yield were made of 18.5% additive effects, 25.9% of maternal effects and 55.6% of non-additive effects indicating that additive, maternal and non-additive effects controlled grain yield, but non-additive effects were extremely predominant.
Combined ANOVA across drought environments showed significant differences (p<0.05) between crosses meaning that genetic variation was significant. However, this genetic variation was made essentially of female additive effects (maternal effects) because male additive (male parents) and non-additive (interaction femalesxmales) effects were not significant (p>0.05). Furthermore, genetic effects did not vary with environments because the interaction crossesxenvironments and its components were not significant (Table 2).
Globally, when all the eight environments were combined, there were highly significant differences between crosses female and male parents (p<0.001), indicating that genetic variation, female and male additive effects were significant. The interaction femalesxmales was also significant (p<0.05) signifying the significance of non-additive effects. Furthermore, F = MSF/MSM was significant (p<0.05) (Table 2).
Female additive effects explained 51.1% of genetic effects, male additive effects constituted 9.4% and thus maternal effects accounted for 41.7% whereas non-additive formed 39.6% of genetic effects. Total additive effects made 60.4% of genetic effects in which 41.7% were maternal effects and 18.7% additive effects. Therefore, when all environments were combined, genetic effects on grain yield comprised 18.7% of additive effects, 41.7% of maternal effects and 39.6% of non-additive effects meaning that grain yield in QPM was essentially under control of maternal and non-additive effects than additive effects.
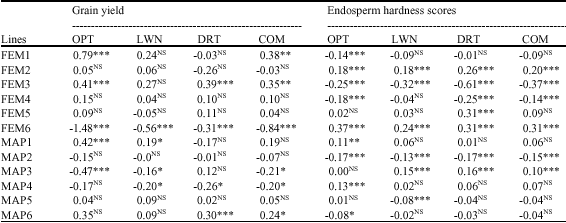
Parental lines FEP1, FEP3 and MAP6 had significant and positive GCAs while MAP 3 and MAP4 had negative and significant GCAs (Table 5). The highest SCA (1.01 t ha-1) across the eight environments was obtained by the combination S65. The second SCA in magnitude (0.56 t ha-1) across all environments was obtained by the combination S66. In general, best combinations for SCA were obtained in crosses between parental lines that had GCAs with opposite signs or between parental lines with one line having non-significant GCA and the other having either negative or positive, but highly significant GCA (Table 4).
| Table 4: | Estimation of GCA effects for grain yield (t ha-1) and endosperm hardness (1-5) |
 | |
| ***Significance at p<0.001, **Significance at p<0.01, *Significance at p<0.05, NS: No Significance (p>0.05), OPT: Optimum, LWN: Low nitrogen, DRT: Drought, COM: All combined | |
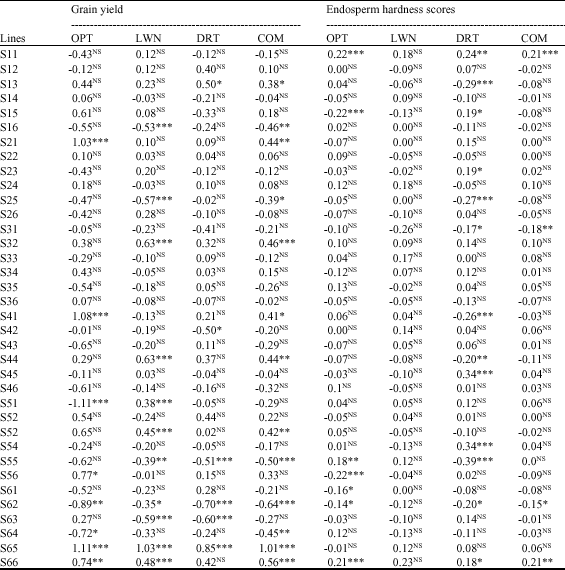
| Table 5: | Estimation of SCA effects for grain yield (t ha-1) and endosperm hardness (1-5) in 36 crosses produced with NCDII |
 | |
| ***Significance at p<0.001, **Significance at p<0.01, *Significance at p<0.05, NS: No Significance (p>0.05), OPT: Optimum, LWN: Low nitrogen, DRT: Drought, COM: All combined | |
Endosperm Hardness
Combined ANOVA across optimum environments revealed highly significant differences (p<0.001) between crosses and hence there was genetic variation. Female and male parents were highly significantly different (p<0.001) and F = MSF/MSM was significant (p<0.05) indicating that female and male additive and maternal effects were significant. Non-additive effects (femalesxmales) were also significant (Table 3).
The interaction crossesxenvironments was highly (p<0.001) significant indicating high interaction of genetic effects with environments. All genetic affects: female, male additive and non-additive effects interacted highly (p<0.001) with environments (Table 3).
Female additive effects accounted 67.8% of genetic effects and male additive effects formed 13.1%, thus total additive effects constituted 80.9% of genetic effects whereas non-additive effects formed 19.1%. Furthermore, total additive effects comprised 54.7% of genetic effects and 26.1% additive effects. Therefore genetic effects of endosperm hardness under optimum environments included 26.1% of additive effects, 54.7% of maternal effects and 19.1% of non-additive effects implying that it was under control of additive, non-additive and maternal effects but maternal effects and additive effects were predominant over non-additive forming more than 80% of genetic variation. Combined ANOVA across low nitrogen and drought environments showed highly significant differences (p<0.001) between crosses indicating significant genetic variation. Female and male parents were highly significant (p<0.001) whereas their interaction was not significant implying that female and male additive effects were highly significant while non-additive effects were not significant. F = MSF/MSM was not significant under low nitrogen condition, but was significant (p<0.05) under drought environments indicating the significance of maternal effects.
The interaction crossesxenvironments was highly significant (p<0.01) indicating that genetic variation interacted with environments. All genetic effects interacted with low nitrogen environments. Female and male additive effects interacted with drought environments, but non-additive effects did not interact with them (Table 3).
Female additive effects accounted 59.3% of genetic effects under low nitrogen environments and 74.8% under drought environments while male additive effects constituted 14.6 and 6.4%, respectively. Total additive effects represented 73.9 and 81.3% of genetic effects and non-additive effects 26.1 and 18.7%, but non-additive effects were not significant. Maternal effects accounted for 44.8% under low nitrogen, although they were not significant and 68.4% of genetic under drought while real additive effects constituted 29.1 and 12.9%, respectively. Hence, under low nitrogen and drought, additive and maternal effects controlled endosperm hardness, but maternal effects were predominant.
Combined ANOVA conducted across the eight environments on endosperm hardness revealed highly significant differences (p<0.001) between crosses and female and male parents. The interaction femalesxmales was highly significant. Hence, female and male additive and non-additive effects were highly significant. F = MSF/MSM was significant (p<0.05) and hence total additive effects contained significant proportion of maternal effects.
Total additive effects represented 89.3% of genetic effects whereas non-additive effects formed only 10.7%. Total additive effects were made of 65.8% of maternal effects and 23.5% of real additive effects. Therefore, overall endosperm hardness was predominantly controlled by maternal effects.
The interaction environmentsxcrosses and its components i.e., environmentsxfemales, environmentsxmales and environmentsxfemalesxmales were highly significant (p<0.01) indicating that genetic variation, female and male additive and non-additive effects interacted significantly with environments (Table 3).
Overall, parental lines FEP3, FEP4, MAP2 had negative and significant GCAs whereas FEP2 and MAP3 had positive and significant GCAs. Furthermore, FEP1, MAP5 and MAP6 had negative but non-significant GCAs whereas FEP5, MAP1 and MAP4 had positive, but non-significant GCAs (Table 4).
Overall, best combinations that had negative and significant SCAs were S31 and S62. The female parent of the combination S31 had negative and significant GCA and the male parent had positive and non-significant GCA while for the combination 62, the female parent had positive and significant GCA and the male parent had negative and significant GCA. Other combinations for endosperm hardness with negative SCAs, but not significant were S13, S15, S25, S44, S56, S61 and S63. The parents of the combinations S13 and S15 had negative GCAs and the parents of S25, S61 and S63 had both positive GCAs. One parent for the combination S44 and S56 had negative GCA while the other parent had positive GCA (Table 5).
By considering grain yield and endosperm hardness together, FEM1, FEM3 and MAP6 had best GCAs i.e., positive, significant for grain yield and negative for endosperm hardness. On contrary, FEP6, MAP3 and MAP 4 had high and negative GCAs for grain yield and positive GCAs for endosperm hardness (Table 5). Most of the best SCAs for either each trait or the two traits together had at least one parent from the two groups of lines. The best SCAs by considering together grain yield and endosperm hardness were: S13, S21, S41, S44, S52 because they were significant and positive SCAs for grain yield and negative SCAs for endosperm hardness (Table 5).
DISCUSSION
The results from the analysis of genetic variation revealed that under optimum conditions, grain yield was controlled by additive, non-additive and maternal effects of gene action. It has been frequently reported that grain yield is controlled by both additive and non-additive with a net predominance of additive effects in normal maize (Bhatnagar et al., 2004; Lee et al., 2005; Menkir and Ayodele, 2005). Contrary to these reports, non-additive and maternal effects together were considerably predominant than additive in QPM in the present study. Hence grain yield behaves like tryptophan concentration trait in QPM (Ngaboyisonga et al., 2008). Furthermore, maternal effects, with 42%, were the most predominant among the three genetic effects.
The results also showed that maternal and additive effects were more predominant than non-additive effects in control of genetic variation of endosperm hardness. In NCDII, maternal effects are included in total additive effects so that endosperm hardness in QPM under optimum environments depended almost exclusively on additive effects confirming several reports on control of genetic modification of o2-endosperm (Prasanna et al., 2001; Vasal, 2000, 2001). Thus, endosperm hardness behaved like several traits in normal maize especially physiological traits (Lee et al., 2005) and protein concentration in grain trait in QPM (Ngaboyisonga et al., 2008).
The proportions of different genetic effects on endosperm hardness under low nitrogen (additive: 29.1%, non-additive: 26.1% and maternal: 44.8%) were closer to those of optimum. However, additive and non-additive effects slightly increased while maternal effects slightly decreased. Therefore, the effects of low nitrogen on genetic variation of endosperm hardness consisted of reducing all genetic effects, but maternal effects were the most importantly affected making additive and non-additive to look as if they were elevated.
Almost exclusively female additive effects and therefore maternal effects controlled grain yield and endosperm hardness, under drought. The effects of drought on genetic variation of grain yield and endosperm hardness consisted of reducing all genetic effects, but additive and non-additive effects were reduced at non-significance levels.
The effects of low nitrogen and drought on mechanisms of gene action had consequences on GCAs of the lines and SCAs of the crosses. The lines GCAs changed in direction and in magnitude with the change of environment. In general, it appeared that, best SCAs for both grain yield and endosperm hardness were obtained with lines of opposite directions. It followed therefore, that lines should be judged according their potential in crosses not according to their GCAs for grain yield and endosperm hardness. Furthermore, crosses with significant and positive SCAs for grain yield and significant and negative SCAs for endosperm hardness are good candidates to use in QPM production.
CONCLUSION
The genetic variation of grain yield and endosperm hardness in QPM is under control of additive, non-additive and maternal effects of gene action, but maternal and non-additive effects for grain yield and additive and maternal effects for endosperm hardness are more predominant. Low nitrogen and drought affect and change the proportions of different genetic effects by acting on some while others are unaffected. They change as well the magnitudes and direction of GCAs of lines and SCAs of crosses. However, it is possible to have together, significant and positive SCAs for grain yield and significant and negative SCAs for endosperm hardness. Crosses with significant and positive SCAs for grain yield and significant and negative SCAs for endosperm hardness are good candidates to use in QPM production.
REFERENCES
- Azevedo, R.A., C. Demerval, J. Landry, P.J. Lea and C.M. Bellato et al., 2003. Regulation of maize lysine metabolism and endosperm protein synthesis by opaque and floury mutations. Eur. J. Biochem., 270: 4898-4908.
CrossRefPubMedDirect Link - Banziger, M., G.O. Edmeades, D.L. Beck and M.R. Bellon, 2000. Breeding for Drought and Nitrogen Stress Tolerance in Maize: From Theory to Practice. CIMMYT, Mexico, ISBN: 970-648-46-3, Pages: 68.
Direct Link - Bhatnagar, S., F.J. Betran and L.W. Rooney, 2004. Combining abilities of quality protein maize inbreds. Crop Sci., 44: 1997-2005.
CrossRefDirect Link - Edmeades, G.O., M. Banzinger, H. Campos and J. Schussler, 2006. Improving Tolerance to Abiotic Stresses in Staple Crops: A Random or Planned Process? In: Plant Breeding: The Arnel R. Hallauer International Symposium, Lamkey, K.R. and M. Lee (Eds.). Wiley Inc., New York, ISBN: 9780813828244, pp: 293-309.
- Ferreira, R.R., V.A. Varisi, L.W. Meinhardt, P.J. Lea and R.A. Azevedo, 2005. Are high-lysine cereal crops still a challenge?. Braz. J. Med. Biol. Res., 38: 985-994.
CrossRefPubMedDirect Link - Krivanek, A.F., H. De Groote, N.S. Gunaratna, A.O. Diallo and D. Friesen, 2007. Breeding and disseminating Quality Protein Maize (QPM) for Africa. Afr. J. Biotechnol., 6: 312-324.
Direct Link - Lee, E.A., A. Ahmadzadeh and M. Tollenaar, 2005. Quantitative genetic analysis of the physiological processes underlying maize grain yield. Crop Sci., 45: 981-987.
Direct Link - Menkir, A. and M. Ayodele, 2005. Genetic analysis of resistance to gray leaf spot of mid-altitude maize inbred lines. Crop Sci., 45: 163-170.
Direct Link - Pixley, K.V. and M.S. Bjarnason, 2002. Stability of grain yield, endosperm modification and Protein quality of hybrid and open-pollinated Quality Protein Maize (QPM) cultivars. Crop Sci., 42: 1882-1890.
Direct Link - Prasanna, B.M., S.K. Vasal, B. Kassahun and N.N. Singh, 2001. Quality protein maize. Curr. Sci. Assoc., 81: 1308-1319.
Direct Link