
Ram C. Sihag
Department of Zoology and Aquaculture, CCS Haryana Agricultural University, Hisar-125004, India
Parvati Sharma
Department of Zoology and Aquaculture, CCS Haryana Agricultural University, Hisar-125004, India
Journal of Fisheries and Aquatic Science
Year: 2012 | Volume: 7 | Issue: 2 | Page No.: 72-103







ABSTRACT
Diseases are considered to be the major constraint in aquaculture production. They cause mortality in shrimp larviculture and fish hatcheries. They are also a constraint on consistent production of fish and shell fish. Traditionally, the control of diseases in aquaculture has relied on the use of chemical compounds. More recently probiotic microorganisms and vaccination or other forms of immunostimulation have also been employed. The abuse of antimicrobials can result in the development of resistant strains of bacteria. Such resistance can be readily transferred to other strains, either following alterations to the existing genome or by transfer of genetic material between cells through plasmids or bacteriophages. The massive use of antibiotics for the control of diseases has been questioned by acquisition of antibiotic resistance in disease causing agents and the need of alternative measures to control these diseases is of prime importance. In recent years, probiotics have a center stage and are used as alternative measures to control the fish diseases. Probiotics have been used by man for millennia since the time humans first consumed fermented milk products. Probiotics can be essential for the normal digestive, endocrine and immunological functions of the bowel. They inhibit pathogenic microorganisms and have been used therapeutically to treat a variety of gastrointestinal and even systemic disorders. Probiotics transiently colonize the bowel and except when used to treat an acute disorder, must be regularly consumed to maintain benefit. Use of microbial probiotics to promote health maintenance and disease prevention and control is now widely accepted as the new ecofriendly alternative measures for sustainable aquaculture.
PDF Abstract XML References Citation
How to cite this article
Ram C. Sihag and Parvati Sharma, 2012. Probiotics: The New Ecofriendly Alternative Measures of Disease Control for Sustainable Aquaculture. Journal of Fisheries and Aquatic Science, 7: 72-103.
DOI: 10.3923/jfas.2012.72.103
URL: https://scialert.net/abstract/?doi=jfas.2012.72.103
DOI: 10.3923/jfas.2012.72.103
URL: https://scialert.net/abstract/?doi=jfas.2012.72.103
INTRODUCTION
Aquaculture provides opportunities for the production of wide variety of aquatic foods including fish and shell fish. Fish is a vital source of food for man. It is very important protein source particularly in regions where livestock is relatively scarce. It is the most important single source of high-quality protein, providing approximately 16% of the animal protein consumed by the world’s, population. The total animal protein intake to human diet, fish contribute 26.2% in Asia, 17.4% in Africa, 9.2% in Europe, 7.4% in North and Central America and 7.2% in South America (FAO, 2006). About one billion people world-wide depend upon fish as their primary source of animal protein. The value of fish traded internationally comes out to be US$ 51 billion per annum. Over 36 million people are employed directly through fishing and aquaculture (FAO, 2000) and as many as 200 million people derive direct and indirect income from fish (FAO, 2006). Consumption of food fish is increasing, having risen from 40 million tones in 1970 to 86 million tones in 1998 and is expected to reach 110 million tones by 2010. Aquaculture has evolved as the fastest growing food-producing sector and has been developed as an important component in the global food security (FAO, 2000).
Aquatic diseases make the biggest constraint in aquaculture production. This is because; water makes a very suitable medium for the proliferation of several diseases in aquatic animals. Bacterial fish diseases like dropsy, hemorrhagic septicaemia, edwardsiellosis/edwardsiella septicemia, bacterial kidney disease, bacterial gill disease, pop eye, vibriosis, Epizootic Ulcerative Syndrome (EUS), ulcerative disease and fin and tail rot were reported by different workers from different parts of the globe in fresh water fishes (Gahlawat et al., 2006). Bacterial pathogens cause heavy mortality in both cultured and wild fish/shell fish species over the world (Bader et al., 2003; Ampofo and Clerk, 2002; Jakhar et al., 2010a).
In the past, several methods have been tried to control the diseases of aquatic animals. These include hygiene maintenance, use of chemicals and use of antibiotics. High stocking densities and over feeding become the cause of aquatic pollution. Entry of wild fish or restocking of ponds with unhealthy or sub quality eggs is another source of ponds becoming unhygienic. Hygien maintenance is, therefore, the most important part of prevention of aquacultural diseases (FAO, 2008). Several chemicals have been recommended for the prevention and control of aquacultural diseases. Simple disinfectants like permanganate, benzalkonium chloride solution or calcium cyanamide have been used as prophylactic measures. Besides these, there are other chemicals in use e.g., formalin (Klontz, 1979), organophosphates (Burridge and Haya, 1995), malachite green and copper sulphate (Gomez-Gil et al., 2000), quaternary ammonium compounds (Snieszkd, 1981), furazolidane (Samuelsen et al., 1998), sulphadiazine (Capone et al., 1996), epson salt and di-n-butyl tin oxide (Burridge and Haya, 1995). Some of these chemicals are highly disease specific whereas others are highly non-specific. Their over doses can pollute the water bodies too.
As a replacement to chemicals, later on, the control of aquacultural diseases was focused on the use of chemotherapeutants, such as erythromycin and deoxycycline (Munday, 1994). At present many antibiotics used in veterinary therapy are commonly used for the control of aquacultural diseases (Smith, 1996). The indiscriminate worldwide use of antibiotics in aquaculture has, however, led to the development of drug-resistant bacteria which are becoming increasingly difficult to control and eradicate (Aoki et al., 1980; Aoki and Watanabe, 1993; Hayashi et al., 1993; Aoki et al., 1995; De Paola, 1995; Sahul and Balasubramanian, 2000; Van der Waaij and Nord, 2000; Bruun et al., 2000; Miranda and Zemelman, 2002). Besides development of drug resistant bacteria and pathogens, the adverse effect of antibiotics is caused by their influence on the aquatic microflora and the retention of harmful residues in aquatic animals (Fuller, 1989; Esiobu et al., 2004; Sartar et al., 2007). Therefore, to keep a sustainable aquacultural growth, health management strategies must go beyond antibiotics and chemotherapeutics. Although, now focus is increasingly being shifted to vaccination (Eldar et al., 1997; Klesius et al., 2000; Evans et al., 2004), yet the dangers of ill effects of antibiotics cannot be ruled out. Of late, the method that is gaining recognition for controlling pathogens within the aquaculture industry is the use of beneficial or probiotic bacteria (Ringo and Gatesoupe, 1998; Gatesoupe, 1999; Ringo and Birkbeck, 1999; Verschuere et al., 2000; Irianto and Austin, 2002; Dahiya et al., 2010; Sihag, 2010). These probiotic bacteria prove harm less to the host as well as human being and result in improved resistance in the host against infectious diseases. Therefore, the objective of prevention and control of disease can be achieved by the use of probiotics. The use of probiotics will prove a new ecofriendly alternative measure for sustainable aquaculture.
DISEASES IN FISH
Several bacteria have been reported as pathogenic to fish and shell fish. These are either obligate or facultative pathogens. The facultative bacterial pathogens prove a potential threat when fish are under environmental and physiological stress (Wedemeyer, 1970). Six gram negative rods (Aeromonas, Proteus, Citrobacter, Pseudomonas, Flavobacterium and Chromobacterium) and three gram positive cocci (Micrococcus, Streptococcus and Staphylococcus) genera of bacteria have been characterized as potential pathogen Aristichthys nobilis and I. idella fingerlings (Shamsudin, 1986; Welker et al., 2005; Verma et al., 2006). Likewise, three bacteria (viz. Vibrio anguillarum, V. alginolyticus and Aeromonas hydrophila) have been reported to cause pathogenecity in Indian magur (Clarias batrachus L.) (Dahiya and Sihag, 2009; Dahiya et al., 2009). Fresh water prawns are also not the exception (Jakhar et al., 2010a). Aeromonas hydrophila, Pseudomonas flourescens and Enterobacter aerogenes were found to cause bacterial necrosis in Macrobrachium rosenbergii (Jakhar et al., 2010b) whereas Micrococcus luteus, M. varians, Cellabiosococcus scuri, Streptococcus grp Q1 and Staphylococcus aureus were found to cause tail rot disease in this animal (Jakhar et al., 2010c).
Over the past two decades, Epizootic Ulcerative Syndrome (EUS) has had serious impact on global fisheries resulting in heavy economic losses. It is one of the most destructive diseases amongst fresh and brackish water fish in the Asia-Pacific region. This disease has spread through rivers, reservoirs and paddy fields to neighboring states, causing considerable loss to fish farmers (Bondad-Reantaso et al., 1992). The first recognized account of a EUS-like condition occurred in Japan in 1971 (Egusa and Masuda, 1971). Later on, similar disease was reported in Australia (1972), Papua Guinea (1975), Indonesia (1980), Thailand (1980), Malaysia (1981), Burma (1984), Laos (1984), Kampuchea (1984) and Srilanka (1988) (Cerro et al., 2002). In natural outbreaks, several fish species, especially air breathing fishes, had been affected by EUS (Roberts et al., 1994). A diverse group of biotic agents such as viruses, bacteria and cutaneous ectoparasites may initiate skin lesions which are subsequently colonized by a fungus (Aphanomyces invadans) and ultimately lead to EUS (Lilley, 1992; Cerro et al., 2002). Different pathogenic organisms, including bacteria (Karunasagar et al., 1995; Pal, 1996), fungus (Roberts et al., 1993) and viruses (Frerichs et al., 1985; Davies and Frerichs, 1989) have been isolated from naturally infected fish. Different bacterial species isolated from the diseased fish include A. punctatus, Flavobacterium sp., Pseudomonas sp., E. tarda, V. parahaemolyticus and Streptococcus sp. (Kumar et al., 1986).
DISEASES AS CONSTRAINT IN AQUACULTURE PRODUCTION
Disease outbreak is being increasingly recognized as one of the most important constraints in aquaculture production in many countries (Nicolas et al., 1989; Grisez et al., 1997; Riquelme et al., 1997). Commercial fish farming creates favorable conditions for the development of infectious diseases in fish. With the increasing fish culture activities, several bacterial diseases, causing morbidity and mortality in fish have been reported in fresh water aquaculture. Pathogenic microorganisms (bacteria, fungi, viruses and protozoan) generally enter the fish through the gills, the skin or gastrointestinal tract (Birkbeck and Ringo, 2005). These three routes of entry represent physical and immunological barriers against pathogens. Thus, their integrity, both at the cell and tissue levels, is vital for the later outcome of the host-pathogen interaction.
Bacterial diseases in fishes were broadly classified as surface ulcerative, acute systemic and chronic granulomatous types. Surface ulcerative types of diseases were characterized by haemorrhagic surface ulcers and were caused by species of Aeromonas, Pseudomonas, Vibrios, Flexibacter and Myxobacter. These infections lead to acute systemic diseases with passage of time. Systemic disease is characterized by the presence and proliferation of bacteria in internal organs like kidneys, heart, spleen, blood and other visceral organs. These diseases produce necrotic changes like bacterial haemorrhagic septicaemia in all affected organs and cause mass mortality. Chronic granulomatous type of disease conditions are characterized by formation of granulomas with the initiating bacteria in the centre of glands (Kumar et al., 1986; Dey, 1989).
Histopathology of EUS affected fishes were studied by various workers in different species of fishes in India and abroad (Kumar et al., 1986; Dey, 1989; Mukherjee et al., 1991; Nayak, 1993; Roberts et al., 1994; Cerro et al., 2002). The changes associated with the disease include cellular infiltration in the hypodermis and epidermis, oedema, degeneration of kidney tubules, depletion of lymphoid cells of spleen and congestion of cerebral tissue. In the initial stages, the disease is marked by little red spots on the skin surface which progresses in size until eventually, a circular to oval deep haemorrhagic ulcer exposing the skeletal musculature is visible. Studies reveals that there is complete loss of epidermis in the ulcerative area of the skin where hypodermis shows characteristic granulomatous changes (Dey, 1989). Most of the hepatocytes showed cloudy swellings. In case of Chinese carp (grass carp and common carp), the affected fish showed ulcer and deep red hemorrhagic lesions in caudal peduncles. Ecological studies carried on Aeromonas sp. and Pseudomonas sp. present in mrigal (Cirrihinus mrigala) in cultured ponds revealed occurrence of high percentage of these bacterial species in EUS affected fishes (Iqbal et al., 1999). The microbial flora of the liver, gills, intestine and muscle of murrel (Channa striatus), infected with Epizotic Uncreative Sgndrome (EUS), was estimated quantitatively and qualitatively. The most predominant bacterial isolates in all the samples were Aeromonas hydrophila, Enterobacter sp., Vibrio sp., Pseudomonas sp., Escherichia coli, along with Aphanomyces invadans and Aspergillus sp. Total viable microbial count was highest in gills (5.9±0.5x107 cfu (colony forming units) g-1) and lowest in intestine (8.7±1.8x104 cfu g-1) which could reveal the presence of bacterial and mycotic species. This observation confirms the association of some bacteria with fungus in EUS disease (Dhanaraj et al., 2008). This disease was reported to occur due to a mixed infection of A. hydrophila and P. fluorescens which had been identified as the most commonly occurring bacterial agents of fish disease (CIFA, 2003). Besides bacteria and fungi, other microorganism like viruses and parasites were also found to be associated with epizootic ulcerative syndrome (Dey, 1989; Kumar et al., 1991; Chowdhury et al., 2003).
The extensive observational studies made during EUS outbreaks in Karnataka and Tamil Nadu (South India) revealed that Indian Major Carp (IMC) present in many water bodies were not affected by this disease (Vishwanath et al., 1997, 1998). IMC appeared resistant in South India but not in North India. There was an anomaly in susceptibility of IMC with regard to geography (Chinabut and Roberts, 1999). This perhaps was due to the fact that the temperature in South India was high enough for the IMC to resist infection (Roberts et al., 1994). Surprisingly, during severe EUS outbreaks in several North and Northeastern states, IMC were found mildly affected whereas other species like Puntius, Channa, Mastacembalus etc. were found severely affected in southern states (Kumar et al., 1991; Das, 1997). EUS outbreaks in freshwater fishes generally occur during winter months and after periods of heavy rainfall coinciding with a fall in water temperature (Lilley et al., 1998). These conditions favoured sporulation of Aphanomyces invadans (Lumanlan-Mayo et al., 1997) and low temperature had been shown to delay the inflammatory response of fish to fungal infection (Chinabut et al., 1995; Catap and Barry, 1998). In all the North and Northeastern states of India, during winter months the temperature normally goes below 20°C which is ideal for EUS outbreaks. The fingerlings and advanced fingerlings of IMC as compared to adult suffer heavy mortality during disease outbreak (Mohan, 2003).
The higher concentration of certain chemicals in water was the predisposing factor for bacterial fin and tail rot disease in fishes (Chakrabarti, 1994). Bacterial gill disease or gill rottening was caused by A. hydrophila, Flavobacterium columnaris and Myxobaterial organisms in common carp (Cyprinus carpio) which showed gill hyperplasia (CIFA, 2003). Fin and tail rot disease was reported in large number of fresh water fishes in India (Kumar et al., 1986; Karunasagar et al., 1989; Jhingran, 1991). During this infection, a white line was observed on the margin of fin, spreading as an imparting frayed appearance on the appendages which later on eventually putrefies and disintegrates. In gill rot disease, gills showed hyperplasia of the secondary lamellar epithelium, fusion of lamella, clubbing of gill tips, necrosis of primary and secondary lamellae of the gill tubular epithelial cells and in some cases oedema and hypotrophy changes in the affected tissues (Dey, 1989).
TRADITIONAL METHODS OF DISEASE CONTROL IN AQUACULTURE
Hygiene maintenance: Hygien maintenance is the most important part of prevention of fish dieases. In traditional methods, pond hygiene used to be the first priority of pond management. Pond must be periodically dried and then refilled with abundant water of good quality. Pond should be well maintained to avoid silting and weeds are controlled. High stocking densities, over feeding and pollution are avoided. Entry of wild fish or restocking of ponds with unhealthy or sub quality eggs and young fish are prevented. Too frequent handling, transfer or transport of pond fish is avoided. The condition of gills must be periodically checked in random samples of the selected fishes. As soon as the onset of a disease is noticed, the seriously affected, moribund or the dead fish must be removed from ponds and buried with quicklime (FAO, 2008).
Use of chemicals: Prophylactic measures are taken to disinfect the pond and the gears. The nets and other tools are routinely disinfected with benzalkonium chloride solution. Ponds having fish and threatened with disease outbreak are disinfected with either potassium permanganate (0.5 g/100 L) or benzalkonium chloride solution (600 ppm) or calcium cyanamide.
Formalin (40% formaldehyde) is a traditionally tried chemical. It is very effective at concentrations of 167 to 250 mg L-1 (Klontz, 1979) against parasitic infections of protozoan ectoparasites (Costia) and trematodes like Discocotyle. Organophosphates are useful for such ectoparasites where formalin fails. These are effective against crustacean parasites like Lernaea and Argulus and such trematodes as Gyrodactylus (Burridge and Haya, 1995). Malachite green is the next most tried chemical, especially effective against fungal infections including Saprolegnia. It is used at a concentration of 1-2 mg L-1. Copper sulphate is effective against bacterial infections but it is not favoured for reasons of its toxic effects (Gomez-Gil et al., 2000). Quaternary ammonium compounds are very effective against bacterial infections. These are generally used at concentrations of 1-4 mg L-1 (Snieszkd, 1981). Furazolidane, a nitrofuran, popular in veterinary therapy, is used in USA for incorporation in fish food to control/prevent bacterial infection; the recommended dose is 11 g/100 kg fish/day for about a week (Samuelsen et al., 1998). Sulphadiazine, a sulphonamide, mixed with trimethoprim, is also found to be equally effective against bacterial diseases when administered through diet; the recommended dose is 51/2 g/100 kg fish/day for a week (Capone et al., 1996). These chemicals are, however, non specific with low efficacy, that is why, these are less reliable. Epson salt (magnesium sulphate) mixed with diet when given is found very useful in treatment of protozoan parasites of the gut. Di-n-butyl tin oxide also administered via diet with a dose of 25 g/100 kg fish/day for three days is effective against most parasites of the gut other than protozoans (Burridge and Haya, 1995).
Use of antibiotics: Earlier, the control of fish diseases was focused on the use of chemotherapeutants, such as erythromycin and deoxycycline (Munday, 1994). Oxytetracycline an antibiotic used in veterinary therapy is commonly used for bacterial therapy of vibriosis and ulcer disease in fishes; the recommended dose is 71/2 g/100 kg fish/day for one to two weeks (Smith, 1996). But now focus is increasingly being shifted to vaccination (Eldar et al., 1997; Klesius et al., 2000; Evans et al., 2004). Combined with the problem of antibiotic contamination of aquaculture facilities and livestock, the indiscriminate worldwide use of antibiotics in aquaculture has led to the development of drug-resistant bacteria which are becoming increasingly difficult to control and eradicate (Aoki et al., 1980; Aoki and Watanabe, 1993; Hayashi et al., 1993; Aoki et al., 1995; De Paola, 1995; Sahul and Balasubramanian, 2000; Van der Waaij and Nord, 2000; Bruun et al., 2000; Miranda and Zemelman, 2002). To keep a sustainable growth pattern, health management strategies must go beyond antibiotics and chemotherapeutics which create resistance in the bacteria and immunosuppression in the host. Besides development of drug resistant bacteria and pathogens, the adverse effect of antibiotics is caused by their influence on the aquatic microflora and the retention of harmful residues in aquatic animals (Fuller, 1989; Esiobu et al., 2004; Sartar et al., 2007). Subsequently, certain antibiotics such as chloramphenicol have been banned in many countries (Roberts, 1997; FAO, 2000).
Use of probiotics in aquaculture: A growing concern for the high consumption of antibiotics in aquaculture has initiated a search for alternative methods of disease control. One of the methods gaining recognition for controlling pathogens within the aquaculture industry is the use of beneficial or probiotic bacteria (Ringo and Gatesoupe, 1998; Gatesoupe, 1999; Ringo and Birkbeck, 1999; Verschuere et al., 2000; Irianto and Austin, 2002). Use of probiotics has been highlighted by many workers (Yazid et al., 1999; Chang and Chen, 2003; El-Naggar, 2004; Ghafoor et al., 2005; Igbasan et al., 2005; Kabir et al., 2005; Trachoo and Boudreaux, 2006; Anukam, 2007; Deeseenthum et al., 2007; Anukam and Koyama, 2007; Raj et al., 2008; Hung et al., 2008; Radfar and Farhoomand, 2008; Capcarova et al., 2008; Fazeli et al., 2008; Patel et al., 2008; Soundarapandian and Sankar, 2008; Vamanu et al., 2008; Vijayabaskar and Somasundaram, 2008; Abdelhamid et al., 2009; Al-Otaibi, 2009; Raja et al., 2009; Nasrollah, 2009; Dahiya et al., 2010; Nikfar et al., 2010; Sihag, 2010; Vamanu and Vamanu, 2010; Agouz and Anwer, 2011; Bansal et al., 2011; Gharaei-Fathabad and Eslamifar, 2011; Yesillik et al., 2011). Improved resistance against infectious diseases can be achieved by the use of probiotics.
Probiotics are live microorganisms administerted in adequate amounts as feed or food supplements which have beneficial effects on the intestinal microbial balance of the host. These are emerging as significant microbial food supplements in the field of prophylaxis (Geovanny et al., 2007). In aquaculture, the term Aprobiotics@ is often loosely used to describe a microbial formulation responsible for biocontrol or bioremediation of pathogens. The term probiotics comes from the Greek word “pro bios” meaning “for life”. The original definition of probiotics given by Lilly and Stillwell (1965) “substances produced by one protozoan that stimulated the growth of another” was expanded from an agricultural perspective and redefined as “a live microbial feed supplement which beneficially affects the host animal its intestinal microbial balance” (Fuller, 1989). Probiotics are commonly defined as mono- or mixed cultures of live microbes that, when applied to animal or human, generate a beneficial effect on health of the host. These beneficial effects include disease treatment and prevention as well as improvement of digestion and absorption in the host (Havenaar and Huis, 1992).
However, according to Salminen et al. (1999), probiotics include microbial cell preparations or microbial cell components. Gatesoupe (1999) redefined probiotics for aquaculture as microbial cells that are administered in such a way so that these could enter the gastrointestinal tract of the aquatic animal to be kept there alive, with the aim of improving its health. According to Verschuere et al. (2000), “the ability of a probiotic is to modify the host-associated or ambient microbial community and to improve the quality of its surroundings”.
Various bacteria commercially used as probiotics are: Lactobacillus species viz. L. acidophilus, L. casei, L. fermentum, L. gasseri, L. johnsonii, L. lactis, L. paracasei, L. plantarum, L. reuteri, L. rhamnosus, L. salivarius; Bifidobacterium species viz. B. bifidum, B. breve, B. lactis, B. longum and Streptococcus species viz. S. thermophilus. Likewise, non-bacterial sources of probiotics are yeast. The yeast, Saccharomyces cerevisiae, has also been commonly identified as probiotic. It stimulates immune system of fish for the production of inhibitory substances against foreign antigen (Dahan et al., 2003). Yeasts are not affected by antibiotics. This is advantageous in probiotic preparations used for preventing disturbances in the normal microflora in the presence of antibacterial metabolites. Strains of Saccharomyces cerevisiae and Dermocystidium hansenii have been shown to attach and grow in the intestinal mucus of fish.
Several reports suggest that dietary yeast or nucleotides act as probiotics in fishes enhancing their immune function and disease resistance (Ramadan et al., 1994; Wang and Zhang, 2000; Burrells et al., 2001a, b), besides improving their growth performance (Adamek et al., 1996; Kubitza et al., 1997; Burrells et al., 2001b). It has been proposed that de novo synthesis and salvage pathway of nucleotides is a costly metabolic process; a dietary supply of nucleotides or precursors may have a protein sparing effect (Sanderson and He, 1994).
Probiotics protect their host against neighboring or invading pathogens by interfering with their cellular functions. Probiotics may protect their host from pathogens by producing metabolites which inhibit the colonization/growth of other microorganisms or by competing with them for resources such as nutrients or space (Ouwehand et al., 1999; Ouwehand et al., 2001; Forestier et al., 2001; Pinchuk et al., 2001; Mukai et al., 2002; Fiorillo et al., 2002; Servin and Coconnier, 2003; Vine et al., 2004a, b).
The addition of potentially probiotic microorganisms to culture water in larval fish systems is a means of biocontrol. It is possible that some of these may be ingested and have a probiotic effect on the host animal (Nogami and Maeda, 1992; Jory, 1998; Moriarty, 1998; Verschuere et al., 1999; Ruiz-Ponte et al., 1999; Douillet, 2000; Chythanya et al., 2002).
PROBIOTIC SELECTION CRITERIA
In vitro antagonism tests: A common way to select a probiotic is to perform in vitro antagonism tests, in which pathogens are exposed to the probiotics or their extracellular products in liquid (Gram et al., 1999) or solid (Austin et al., 1992) medium. Depending on the exact arrangement of the tests, candidate probiotics can be selected based on the production of inhibitory compounds (Gram et al., 1999) or siderophores, or on the basis of competition for nutrients (Dopazo et al., 1988). The preselection of candidate probionts based on these in vitro antagonism tests has often led to the finding of effective probiotics.
Colonization and adhesion: A probiotic should either be supplied on a regular basis or be able to colonize and persist in the host or in its ambient environment. The ability of a strain to colonize in the gut or an external surface of the host and adhere to the mucus layer may be a good criterion for preselection among the putative probiotics (Shi and Walter, 2004). This involves the viability of the potential probiotic within the host and/or within its culture environment, adherence to host surfaces and the ability to prevent the establishment of potentially pathogenic bacteria (Panigrahi and Azad, 2007).
In vivo evaluation of effects of potential probiotics on the host: The effect of candidate probiotics should be tested in vivo as well. When the probiotic effect is supposed to be nutritional, the candidate probiotics could be added to the culture of the aquatic species and their effect on growth and survival parameters could be assessed. However, when biological control of the microbiota is desired, representative in vivo challenge tests seem to be the appropriate tool to evaluate the potential effect of the candidate probiotic on the host (Gildberg et al., 1995). The approach outlined above should result in formulating a set of strains with a well-established probiotic effect on the target organism without affecting other possibly involved trophic levels. Comparative pilot experiments under hatchery or grow out conditions in the farms should be performed to estimate the economic consequences of the probiotic application. An important factor in the economic evaluation is the mass production of the probiont. Also, effective legislation, if any, should be taken into account before commercial application is begun. Finally, a cost-benefit analysis will determine whether the probiotics could be applied in practice or not (Verschuere et al., 1999; Ruiz-Ponte et al., 1999; Douillet, 2000). In general, the selection criteria for a bacterium to be used as a probiotic should be followed. It should be non-pathogenic; withstand incorporation into a delivery vehicle at high cell counts and remain viable throughout the shelf-life of the product; withstand transit through the gastrointestinal tract, i.e., show acid and bile tolerance; be able to adhere to cells of the intestinal epithelium and/or colonize the lumen of the tract; show antagonistic activity towards enteric pathogens and/or provide demonstrated health benefits (Chythanya et al., 2002).
A probiotic must possess certain properties in order to aid in correct establishment of new, effective and safe products (Verschuere et al., 2000). It should not be harmful and be acceptable to the host through ingestion, potential colonization and replication within the host. It should reach the location where the effect is required to take place. It should actually work in vivo as opposed to in vitro findings. It should preferably not contain virulence resistance or antibiotic resistance genes.
POSSIBLE MODES OF ACTION
Production of antagonistic compounds: Antagonistic compounds are described as chemical substances produced by bacteria which are toxic or inhibitory towards other micro organisms (Ramirez and Dixon, 2003). These substances may be produced as either primary or secondary metabolites and, therefore, have different modes of inhibitory action. Microbial populations may release chemical substances that have a bactericidal or bacteriostatic effect on other microbial populations which can alter interpopulation relationships by influencing the outcome of competition for chemicals or available energy (Fredrickson and Stephanopoulos, 1981). The presence of bacteria producing inhibitory substances in the intestine of the host, on its surface, or in its culture medium is thought to constitute a barrier against the proliferation of opportunistic pathogens.
In general, inhibitory effect of bacteria is due to many factors, either singly or in combination with others. These include production of antibiotics (Williams and Vickers, 1986), bacteriocins (Bruno and Montville, 1993), siderophores, lysozymes, proteases, and/or hydrogen peroxide and also the alteration of pH values by the production of organic acids (Sugita et al., 1997). Lactic acid bacteria are known to produce compounds such as bacteriocins that inhibit the growth of other microorganisms (Vandenbergh, 1993). There are many reports of inhibitory activity of lactic acid bacteria mediated by bacteriocins. Many bacteria have been identified to produce bacteriolytic enzymes against Vibrio parahaemolyticus (Nair et al., 1985). Bacteriocins are proteinacious agents produced by bacteria to inhibit or kill other bacteria. Other inhibitory compounds produced by bacteria include organic acids, hydrogen peroxide, carbon dioxide and siderophores. Siderophores are iron-complexing chemicals secreted by bacteria and fungi. Siderophore-producing bacteria can survive in nutrient-poor environments (Vine et al., 2004a, b).
Competition for chemicals or available energy: Competition for chemicals or available energy may determine how different microbial populations co-exist in the same ecosystem (Fredrickson and Stephanopoulos, 1981). The microbial ecosystem in aquaculture environments is generally dominated by heterotrophs competing for organic substrates as both carbon and energy sources. Specific knowledge of factors governing the composition of microbiota in aquaculture systems is required to manipulate the latter. Verschuere et al. (1999) selected several strains of bacteria with a positive effect on the survival and growth of Artemia juveniles. It was suggested that the selected bacteria exerted their protective action by competing with the pathogen for chemicals and available energy.
Competition for iron: Virtually all microorganisms require iron for growth (Reid et al., 1993). Siderophores are low-molecular-weight (<1,500), ferric ion-specific chelating agents which can dissolve precipitated iron and make it available for microbial growth. The ecological significance of siderophores resides in their capacity to scavenge an essential nutrient from the environment and deprive competitors of it. Successful bacterial pathogens are able to compete successfully for iron in the highly iron-stressed environment of the tissues and body fluids of the host. The ecological significance of siderophores in soils as important tools for iron acquisition by microorganisms and plants and their involvement in suppression of plant root pathogens have been established (Wang and Zhang, 2000). Harmless bacteria which can produce siderophores could be used as probiotics to compete with pathogens whose pathogenicity is known to be due to siderophore production and competition for iron or to out compete all kind of organisms requiring ferric iron from solution (Gullian et al., 2004). The possible effectiveness of siderophore-producing probiotics can be illustrated by the fact that the addition of bacterial siderophore to live food (rotifers) increased the resistance of turbot (Scophthalmus maximus) larvae challenged with the pathogenic Vibrio strain. The addition of a siderophore-producing Vibrio strain protected the turbot larvae slightly more than without such addition (Gatesoupe, 1997).
Competition for adhesion sites: One possible mechanism for preventing colonization by pathogens is competition for adhesion sites on gut or other tissue surfaces. The ability to adhere to enteric mucus and wall surfaces is necessary for bacteria to become established in fish intestines (Westerdahl et al., 1991). Adhesion can be non specific, based on physicochemical factors, or specific, involving adhesin molecules on the surface of adherent bacteria and receptor molecules on epithelial cells. Adhesion capacity and growth on or in intestinal or external mucus has been demonstrated in vitro for fish pathogens like Vibrio anguillarum and Aeromonas hydrophila (Krovacek et al., 1987) and for candidate probiotics such as Carnobacterium strain K1 and also for several isolates inhibitory to V. anguillarum. In one of these studies, the aim was to measure the in vitro capacity of the strains to adhere to and grow in turbot intestinal mucus in order to investigate their potential to colonize the intestine of farmed turbot as a means of protecting the host from infection by V. anguillarum (Chabrillon et al., 2006; Watson et al., 2008).. The intestinal isolates generally adhered much better to a film of turbot intestinal mucus, skin mucus and bovine serum albumin than did V. anguillarum, indicating that they could compete effectively with the pathogen for adhesion sites on the mucosal intestinal surface (Olsson et al., 1992; Isolauri et al., 2001).
Enhancement of the immune response: Immunostimulants are chemical compounds that activate the immune systems of animals and provide them more resistance to infections by viruses, bacteria, fungi and parasites (Raa, 1996). Fish larvae, shrimps and other invertebrates have immune systems that are less well developed than adult fish and are dependent primarily on nonspecific immune responses for their resistance to infection (Sakai et al., 1999; Balcazar, 2003). Observations obtained in various experiments with warm-blooded animals indicate that probiotic (lactic acid) bacteria administered orally may induce increased resistance to enteric infections (Nikoskelainen et al., 2003). The ingestion of bacteria and subsequent endocytosis in cod and herring larvae have been suggested to be involved in the stimulation of their developing immune system (Brunt and Austin, 2005; Selvaraj et al., 2005).
Improvement of water quality: In several studies, water quality has been recorded to be improved during the addition of the probiotics, especially Bacillus spp. The rationale is that gram-positive Bacillus spp. are generally more efficient in converting organic matter back to CO2 than are gram-negative bacteria which would convert a greater percentage of organic carbon to bacterial biomass or slime (Stanier et al., 1993). It is reasoned that by maintaining higher levels of these gram-positive bacteria in the production pond, farmers can minimize the buildup of dissolved and particulate organic carbon during the culture cycle while promoting more stable phytoplankton blooms through the increased production of CO2 (Nogami and Maeda, 1992; Jory, 1998; Moriarty, 1998; Verschuere et al., 1999; Ruiz-Ponte et al., 1999; Douillet, 2000; Chythanya et al., 2002).
Interaction with phytoplankton: When probiotic bacteria are selected to be used in a culture environment comprising algae, their possible interaction with these unicellular algae must be taken into consideration when the mode of action is being investigated (Ramirez and Dixon, 2003). Recent reports demonstrate that some bacterial strains have a significant algicidal effect on many species of microalgae, particularly on red tide plankton (Fukami et al., 1997). Some bacterial strains inhibited the growth of the unicellular alga (Pavlova lutheri) to various degrees (Munro et al., 1995). However, bacterial antagonism toward algae would be undesirable in the situations of larval rearing where unicellular algae are added (e.g., the green-water technique). It would be advantageous when undesired algal species develop in the culture pond. Conversely, positive effects of bacteria on cultured microalgae have also been observed (Fukami et al., 1997). Therefore, it is conceivable that bacteria can indirectly influence the health or the zootechnical performance of the cultured aquatic animals through their effect on the microalgae used as food.
Vitamin production: Lactic acid bacteria produce small amounts of certain B vitamins, including foliates and vitamin B (Kumari and Sahoo, 2005; McFarland, 2006). Microbial synthesis of vitamin K and vitamin C in the intestine appears to have nutritional significance in most animal species (Conway, 2001; Nayak et al., 2007). Bifidobacteria, streptococci and enterococci have been shown to produce vitamin K (Bentley and Meganathan, 2007).
Reduction of cholesterol: Some probiotics can lower total serum cholesterol and low density lipoprotein cholesterol. In vitro studies have shown that Lactobacillus casei and L. acidophilus effectively remove cholesterol from culture media. It is postulated that lactic acid bacteria assimilate cholesterol in the gut or deconjugate bile acids, thus disrupting the intestines-to-liver circulation of cholesterol (Anderson and Gilliland, 1999; Dambekodi and Gilliland, 1998; Sanders, 2000).
Short-chain fatty acid (SCFA) production: Probiotics, especially the bifidobacteria (Bifidibacterium bifidum, B. breve, B. lactis, B. longum) are able to break down and metabolize non-digestible carbohydrates such as fiber. The major by-products of this process are Short-chain Fatty Acids (SCFA) such as lactate, acetate, propionate and butyrate. SCFA lower down intestinal pH and create an environment inhospitable to pathogenic bacteria such as E. coli and Salmonella species. SCFA nourish mucosal cells of colon supplying 60-70% of colonocyte energy needs. Butyrate is the preferred energy source for colonocytes. Studies in animals and humans have found that SCFA directly stimulated colonic calcium, magnesium and potassium absorption, increased colonic blood flow, enhanced tissue oxygenation and transport of nutrients and might be of therapeutic value for various intestinal disorders (Scharrer and Lutz, 1990; Topping and Clifton, 2001; Zhenming et al., 2006; Jha et al., 2007).
Mode of application of the probiotics: The probiotics can be added to the host or its ambient environment in different ways e.g., (1) addition to the artificial diet (2) addition to the culture water and (3) bathing and addition through live food (Verschuere et al., 2000; Gomez-Gil et al., 2000).
The concept of inoculating the culture system and adding to live food is advisable when the small volumes are used. The inclusion in the artificial diet is more adapted when the greater volumes are used; however, this requires sufficient number of probiotics to last when they reach the gastrointestinal tract and exhibit capacity to adhere to the epithelial mucosa.
ROLE OF PROBIOTICS IN AQUACULTURE
Although plenty of work has been done on the role of probiotics in aquaculture during the past decade, yet the research has generally been of an applied nature with few discussions on the bacterial mode of action.
The use of lactic acid bacteria as probiotic (Perdigon et al., 1995; Salminen et al., 1998; Kontula, 1999; Tannock, 1999) and knowledge regarding aspects of this organism’s biology, culture and suitability has been of benefit to aquaculture (Gildberg and Mikkelsen, 1998; Ringo and Gatesoupe, 1998; Nikoskelainen et al., 2003). A number of general review articles on the use of probiotics in aquaculture are also available (Ringo and Gatesoupe, 1998; Gatesoupe, 1999; Skjermo and Vadstein, 1999; Ringo and Birkbeck, 1999; Verschuere et al., 2000; Irianto and Austin, 2002). In aquaculture, the exposure to a diverse bacterial microflora is limited to the available resources. Therefore, the gastrointestinal flora usually resembles the microflora initially present in the rearing water, microalgae and livefood (Tanasomwang and Muroga, 1989; Munro et al., 1994; Ringo et al., 1996; Gatesoupe, 1999; Riquelme et al., 2001). The development of the microbial community in Artemia populations is influenced by both deterministic and stochastic factors (Verschuere et al., 1997). Deterministic factors include salinity, temperature, feed quality etc., while organisms in the right place at the right time to enter the habitat and proliferate make up the stochastic factors. The combination of controllable and chance factors determines the composition of the resulting microflora, making research on probiotics for larviculture both interesting and challenging (Gildberg and Mikkelsen, 1998; Calo-Mata et al., 2008).
Manipulation of beneficial bacterial population present in the bivalve larval cultures is a potentially useful strategy for the enhancement of oyster production. Douillet and Langdon (1994) have reported the use of probiotics for the culture of larvae of the Pacific oyster (Crassostrea gigas Thunbeerg). They added the probiotic bacteria as a food supplement to the larval cultures of this species which consistently enhanced the growth of larvae during different seasons of the year. They suggested that the action of probiotic bacteria were to provide essential nutrients that were not present in the algal diets or to improve the oyster's digestion by supplying digestive enzymes to the larvae or to remove the metabolic substances released by the bivalves or the algae. Likewise, based on the photosynthesis of micro algae mainly, it was clarified that the bacteria, protozoa and other microorganisms from microbial food assemblages play a significant role in the aquatic food chain and help in increasing the crustacean production (Maeda and Liao, 1994).
Cell free extracts of three strains of Lactic Acid Bacteria (LAB) i.e., Lactobacillus acidophilus, Lactobacillus bulgaricus-56 and Lactobacillus bulgaricus-57 inhibited growth of Vibrio alginolyticus in nutrient broth (Ajitha et al., 2004). The antagonism of lactic acid bacteria to Vibrio alginolyticus was further confirmed by streak plating wherein suppression of growth of Vibrio was reported. Animals such as shrimp maintained on a diet devoid of bacterial biomass exhibited 80% mortality while no external or internal pathological changes were observed in shrimp fed with the lactic acid bacteria incorporated diets. These results showed an inhibition of V. alginolyticus by the lactic acid bacteria and the stimulation of the non-specific immune response resulting in the resistance to the disease in the shrimp fed on the lactic acid bacteria incorporated diets. Bacillus spp. has been shown to possess adhesion abilities, produce bacteriocins (antimicrobial peptides) and provide immunostimulation (Duc et al., 2004; Barbosa et al., 2005). The strains appear to be the effective probiotics and the commercial products containing such strains have been demonstrated to improve the shrimp production (Hong et al., 2005; Ghosh et al., 2008).
The marine Bacillus subtilis AB65, Bacillus pumilus AB58 and Bacillus licheniformis AB69 bacteria, isolated from the local marine environment could be used for bioremediation in the shrimp hatcheries and were observed to reduce total nitrogen in Artemia cysts as an alternative to antibiotics. The action of the marine bacteria appeared to be significant in protecting the host shrimp against pathogenic bacteria (Misra et al., 2006). Adenosine 5-diphosphoribose (ADP-ribose); a hydrolysis product of Beta-nicotine Adenine Dinucleotide (NAD) acted as a significant contributor to the anti-cytotoxic activity of probiotic Lactobacillus bulgaricus (John et al., 2007).
The probiotic bacterium acted as an effective treatment against vibriosis and white spot diseases in the giant tiger shrimp (Penaeus monodon Fabricius). Inhibitory activity of two Lactobacillus sp. against Vibrio sp., E. coli, Staphylococcus sp. and Bacillus subtilis was established (Jiravanichpaisal et al., 1997).
Likewise, Vibrio spp. were focused to act as probiotics against some shrimp pathogens (Direkbusarakom et al., 1998). Two isolates of Vibrio spp. which were the dominant composition of the flora in shrimp hatchery showed the antiviral activities against Infectious Haematopoietic Necrosis Virus (IHNV) and Oncorhynchus Masou Virus (OMV) by reducing the number of plaques. The result demonstrates the possibility of using the Vibrio flora as probiotic against the pathogenic viruses in shrimp culture. The use of V. alginolyticus as a probiotic agent may increase survival and growth in Penaeus vannamei postlarvae by competitively excluding potential pathogenic bacteria and can effectively reduce or eliminate the need for antibiotic prophylaxis in intensive larvae culture system (Austin et al., 1995; Garriques and Arevalo, 1995).
Higher survival and molt rates of larvae of prawn (P. monodon) were observed in the experiment treated with soil extract (Maeda and Liao, 1992). It was assumed that if a specific bacterium was cultured and added to the prawn ecosystem to the level of 10 million cells mL-1, other bacteria might hardly inhabit the same biotype because of protozoan activity which would be one of the ways to biologically control the aquacultural pathogenic biotypes.
Microbial food assemblages have been reported to have great utility in the culture of a crab, Portunus trituberculatus (Maeda et al., 1997). Assemblages of microorganisms were produced by adding several nutrients, urea, glucose and potassium phosphate to the natural seawater with gentle aeration in which bacteria and yeast were prevailing. The crab larvae were fed on these microorganisms successively. It was found that some strains of bacteria promoted larval growth. By adopting these assemblages of microorganisms, a high yield was obtained in the larvae of prawn P. japonicus.
Both marine and freshwater fishes have been shown to have a specific indigenous microflora in gastrointestinal tract (Sakata, 1990; Ringo and Strom, 1994; Ringo et al., 1997; Ringo and Gatesoupe, 1998; Kennedy et al., 1998a ,b). The composition of microflora may change with environmental stresses, diet (Munro et al., 1993; Ringo and Strom, 1994; Douillet and Langdon, 1994; Gildberg et al., 1995; Ringo et al., 1997) and fish age (Bergh et al., 1994; Prayitno and Latchford, 1995; Olafsen, 2001). The most common members of the microflora of healthy fish are Vibrio spp., Pseudomonas spp., Acinetobacter spp. (Muroga et al., 1987; Tanasomwang and Muroga, 1989; Nicolas et al., 1989; Cormack and Fraile, 1990; Bergh et al., 1994; Munro et al., 1994; Gatesoupe, 1997), Aeromonas spp., Plesiomonas spp., Pseudomonas spp. and members of the family Enterobacteriaceae (Ugajin, 1979; Sugita et al., 1988; Sakata, 1990; Sugita et al., 1991; Ringo et al., 1995).
It has also been reported that the bacteria isolated from intestine of seven kinds of freshwater cultured fish possess the antibacterial abilities and the presence of the intestinal bacteria can protect the fish against the infection by pathogenic bacteria (Sugita et al 1996).
Lactic acid bacteria were found to produce many kinds of metabolites which affected the other microbes in the mid gut of fish and reduced pH in their luminal contents in the stomach. These bacteria showed higher antagonistic activity by releasing extra cellular proteins or bacteriocins against A. hydrophila and enhanced the production of rotifers which act as biocarriers of probiotic (Vijayabaskar and Somasundaram, 2008).
Bandyopadhyay and Mohapatra (2009) analyzed the effect of a probiotic bacterium Bacillus circulans PB7 in the formulated diets on growth, nutritional quality and immunity of Catla catla (Ham.) The effect of supplement on the growth performance, feed utilization efficiency and immune response was also evaluated.
Likewise, a strain of Carnobacterium sp., isolated from the intestine of Atlantic salmon, was evaluated for potential use as a probiotic for salmonids. In vitro studies demonstrated antagonism of this bacterium against Aeromonas hydrophila, A. salmonicida, Flavobacterium psychrophilum, Photobacterium damselae subsp. piscicida, Streptococcus milleri, Vibrio anguillarum and V. ordalii. Feeding salmonids with diets containing the probiotic having a strain of Carnobacterium sp., revealed that the isolate remained viable in the gastrointestinal tract (Robertson et al., 2000).
Some fluorescent strains of pseudomonad bacteria can also competitively inhibit the growth of fish pathogen, A. salmonicida. The fluorescent pseudomonad is capable of inhibiting the growth of A. salmonicida in culture media and that this inhibition is probably due to competition for free iron (Smith and Davey, 1993).
The micro algae (Tetraselmis suecica) too inhibited Aeromonos hydrophila, A. salmonicida, Serrstia liquefaciens, Vibrio anguillarum, V. salmonicida and Yersnia ruckeri bacteria. When used as a food supplement, the algal cells inhibited laboratory-induced infection in Atlantic salmon. When used therapeutically, the algal cells and their extracts reduced mortalities caused by A. salmonicida, Ser. liquefaciens, V. anguillarum, V. salmonicida and Yersnia ruckeri. It was suggested that there might be some bioactive compounds in the algal cells and there appeared to be a significant role for Tetraselmis in the control of fish diseases (Austin et al., 1992).
Micrococcus luteus and Pseudomonas species from the gonads and intestine of Nile tilapia, Oreochromis niloticus were found to antagonize Aeromonas hydrophila with inhibition zone of 4 and 9 cm diameter, respectively; M. luteus acted as a probiotic in vivo, while Pseudomonas species showed probiotics effects in vitro only (Azza et al., 2009).
The intestinal microflora of turbot (Scophthalmus maximus) larvae was strongly affected by the addition of Vibrio pelagius to the rearing water (Ringo et al., 1996). Similarly, 70% of the intestinal microflora of cod (Gadus morhua) larvae was comprised of Lactobacillus plantarum when exposed to the bacteria in the larval rearing water compared to 1% in unexposed tanks (Strom and Ringo, 1993).
Studies were made to selection, identification and characterization of bacteria. To control a disease in the rearing turbot (Scophthalmus maximus) larvae, thirty-four out of 400 bacterial strains that mainly identified as Roseobacter spp. based on phenotypic criteria exhibited in vitro anti-bacterial activity against three fish larval pathogens (V. anguillarum, V. splendidus and Pseudoalteromonas). The mortality of larvae decreased significantly in treatments where Roseobacter sp. strains at 107 cfu mL-1 was added, indicating a probiotic potential of this bacterium (Hjelm et al., 2004).
Two probiotics viz. lactic acid bacterium (Pediococcus acidilactici) and yeast (Saccharomyces cerevisiae var. boulardii) were tested as alternative treatment to limit the prevalence of the Vertebral Column Compression Syndrome (VCCS) in rainbow trout, compared with a preventive treatment with florfenicol. Either the antibiotic or a lactic acid bacterium was introduced into the experimental diets which were compared with the control diet without supplementation. Pediococcus acidilactici provided the same level of protection as did the antibiotic, without any adverse effect on survival of rainbow trout. The long-term dietary supplementation with P. acidilactici seemed promising as a preventive treatment against this syndrome (Aubin et al., 2005).
To determine the influence of probiotics on the growth and gut microbial load of juveniles of goldfish, three probiotics (sporolac, lactobacil and yeast) were added to the basal diets at 0.5% level and the juvenile goldfish were fed for a period of 60 days. Sporolac incorporated feed recorded a maximum mean weight gain of 0.673 g and a six-fold increase in growth compared to control feed. The percentage composition of Lactobacillus in the Carassius auratus fed with control feed was comparatively low (2.7%) than those supplemented with probiotics (Ahilan et al., 2004).
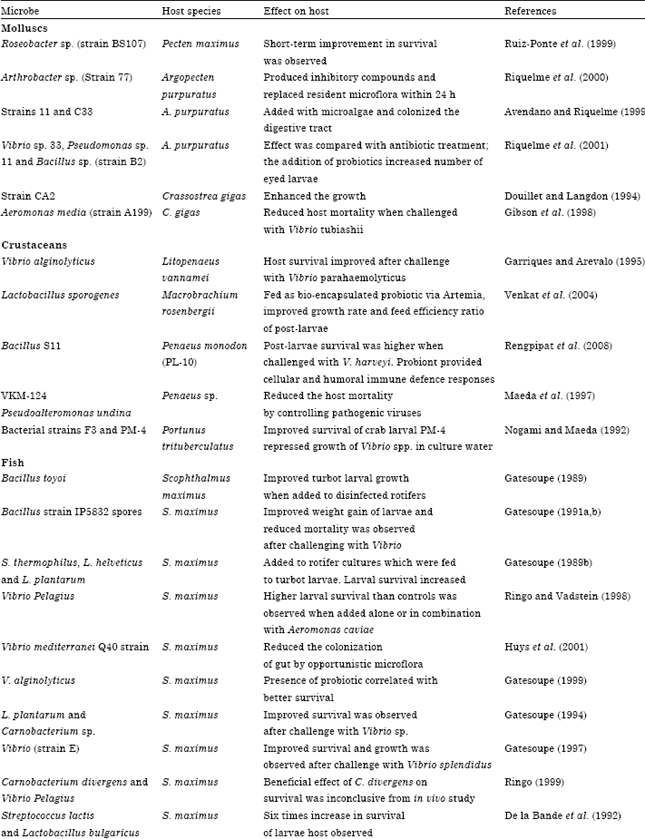
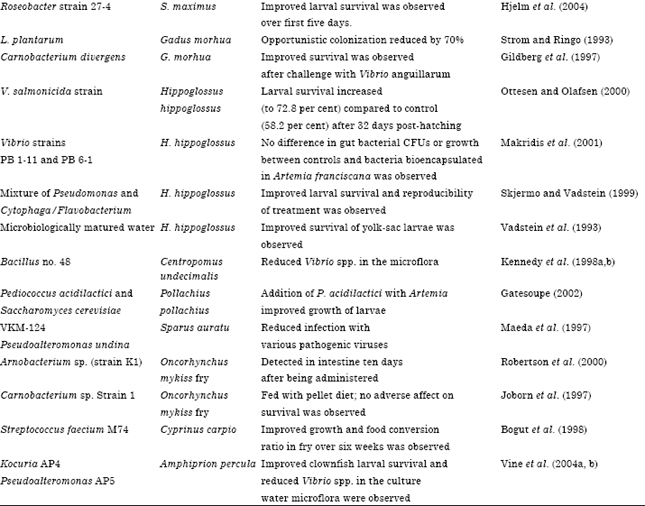
A variety of microorganisms have been used as probiotics to improve the growth or survival of larvae of aquatic species (Table 1).
| Table 1: | Intestinal probiotics used in fish/shellfish and their effects on the host |
  | |
PROBIOTICS AND HAEMATOLOGICAL PARAMETERS
Hematological parameters of fish are used as indicators of their physiological state and their study has become widespread in the control of pathogens and manipulation of stress in fish (Bansal et al., 1979). Bacillus subtilis, a Gram-positive, aerobic, endospore forming bacterium, evaluated for its probiotic potential in Indian major carp, Labeo rohita (Rajesh et al., 2006, 2008). The fish (15±2 g) were given a feed containing B. subtilis in three concentrations for 2 weeks, e.g., 0.5x107, 1.0x107 and 1.5x107 cfu g-1 feed. The control group was given feed without B. subtilis for the same period. Haematological and serum parameters were monitored at weekly intervals. The response variables were total erythrocyte count, Total Leucocyte Count (TLC), haemoglobin, total protein, albumin, globulin, albumin-globulin ratio, alkaline phosphatase activity, alanine aminotransferase activity and aspartate aminotransferase activity. Fish were challenged intraperitoneally with a virulent strain of Aeromonas hydrophila after 2 weeks of feeding to the treatment groups and positive control group, while the negative control group was challenged with phosphate-buffered saline only. Clinical signs and symptoms and mortality/survival percentage were noted in each group. The B. subtilis-treated fish at 1.5x107 cfu g-1 feed showed maximum percent survival (87.50%), weight gain (35.5%), TLCs (3.23x104 cells mm3), haemoglobin content (7.4 g/100 mL), total protein (2.37 g dL-1) and globulin content (1.28 g dL-1) during the pre-challenge. Enzymes showed higher activities during post challenge. The result suggests that B. subtilis can be used effectively as a commercial product for use in aquaculture.
However, nonsignificant changes in the hematological and the biochemical parameters were revealed after 1 day of infection with the fungus (Saprolegnia parasitica) on the hematological, serum biochemical and pathological in Tilapia nilotica but after 7 days of post-infection and 10 days of post-treatment, a significant decrease in red blood cell, haemoglobin, packed cell volume and significant increase in aspartate aminotransferase, urea, creatinine, sodium, cortisol, insulin and glucose were seen (Mona-Mori and Matsumoto, 2008). Iron showed a significant decrease during the same period of sampling. The pathological examination revealed a massive fungal growth resembling a tuft of cotton wool threads in eyes, gills, fins and in localized areas of the skin. Microscopically, the fungal hyphae and spores appeared on eyes, gills, skin and underlying muscles with marked degenerative, necrotic and inflammatory reactions. These reactions were evident after 7 days of post-infection but the severity of the lesions were markedly decreased after 10 days of post-treatment. It could be concluded that fungus infection induced marked tissue alterations as well as some hematological and serum biochemical changes.
Likewise, decrease in the level of blood in the common carp after exposure to Cyanobacteria extract was observed (Palikova et al., 2004). When the spleen, liver and kidney of unhealthy Nile tilapia (Oreochromis niloticus) held fairly severe infection, the haematopoiesis was also severely affected. The latter, in turn, affected the peripheral blood by decreasing erythrocyte volume (Ishikawa, 1998). The erythrocytes count and haematocrit of Nile tilapia decreased when inoculated with Mycobacterium marinum. Perhapes this decrease led to a tendency to develop hypochromic and microcytic anaemia in this fish (Ranzani-Paiva et al., 2004).
But there was an increase in the erythrocyte count in some other fish fed on probiotic bacteria (Irianto and Austin, 2002). Rajesh et al. (2006) also reported that the probiotics used in carps increased the blood parameter values as a result of hemopiotic stimulation. Similarly, an increase in total leukocyte counts and proportion of neutrophils and monocytes was observed when the carp (Cyprinus carpio) were fed with Saccharomyces cerviase (Selvaraj et al., 2005). Irianto and Austin (2002) used dead probiotic cells to control disease and observed higher number of leucocytes, erythrocytes, macrophages in rainbow trout (Oncorhynchus mykiss). Rengipipat et al. (2000) in black tiger shrimp (Penaeus monodon) and Siwicki et al. (1994) in rainbow trout (Oncorhynchus mykiss) found disease resistance or protection in use of probiotics Bacillus sp. and also reported increase in the level of selected hematological parameters in the blood (Red Blood Cell count (RBC), Hematocrit (Ht), Hemoglobin (Hb), various leukocyte counts, the total leukocyte level and MCV, MHC and MCHC counts.
CONCLUSION
Diseases take a heavy toll of the aquaculture production. Traditionally, these diseases are controlled by using chemical compounds. The use of antimicrobials is a common practice in practically all types of hatcheries. The latter can result in the development of resistant strains of bacteria. Such resistance can be readily transferred to other strains. Therefore, the need of alternative measures to control these diseases is of prime importance. In recent years probiotics have a center stage and are used as alternative measures to control the aquatic diseases. Probiotics inhibit pathogenic microorganisms and have been used therapeutically to treat a variety of gastrointestinal and even systemic disorders. The use of probiotics will prove a new ecofriendly alternative measure for sustainable aquaculture.
ACKNOWLEDGMENTS
We are thankful to the Head, Department of Zoology and Aquaculture, CCS Haryana Agricultural University, Hisar for the general facilities. Parvati Sharma received financial assistance in the form of Merit Stipend from CCS HAU during the course of preparation of this review.
REFERENCES
- Ajitha, S., M. Sridhar, N. Sridhar, I.S.B. Singh and V. Varghese, 2004. Probiotic effects of lactic acid bacteria against Vibrio alginolyticus in Penaeus (Fenneropenaeus) Indicus. Asian Fish. Sci., 17: 71-80.
Direct Link - Anderson, J.W. and S.E. Gilliland, 1999. Effect of fermented milk (yogurt) containing Lactobacillus acidophilus L1 on serum cholesterol in hypercholesterolemic humans. J. Am. Coll. Nutr., 18: 43-50.
CrossRefDirect Link - Austin, B., E. Baudet and M.B.C. Stobie, 1992. Inhibition of bacterial fish pathogens by Tetraselmis suecica. J. Fish Dis., 15: 55-61.
Direct Link - Austin, B., L.F. Stuckey, P.A.W. Robertson, I. Effendi and D.R.W. Griffith, 1995. A probiotic strain of Vibrio alginolyticus effective in reducing diseases caused by Aeromonas salmonicida, Vibrio anguillarum and Vibrio ordalii. J. Fish Dis., 18: 93-96.
CrossRef - Avendano, R.E. and C.E. Riquelme, 1999. Establishment of mixed-culture probiotics and microalgae as food for bivalve larvae. Aquacult. Res., 30: 893-900.
Direct Link - Azza, M.A., A.E. Khattab and M.E. Shalaby, 2009. Micrococcus luteus and Pseudomonas species as probiotics for promoting the growth performance and health of nile tilapia, Oreochromis niloticus. Fish Shellfish Immunol., 27: 175-180.
CrossRef - Bader, J.A., C.A. Shoemaker and P.H. Klesius, 2003. Rapid detection of columnaris disease in channel catfish (Ictalurus punctatus) with a new species specific 16S rRNA gene based PCR primer for Flavobacterium columnare. J. Microcbiol. Met., 52: 209-220.
CrossRef - Misra, C.K., B.K. Das, S.C. Mukherjee and P.K. Meher, 2006. The immunomodulatory effects of tuftsin on the non-specific immune system of Indian major carp, Labeo rohita. Fish Shellfish Immunol., 20: 728-738.
CrossRef - Barbosa, T.M., C.R. Serra, R.M. La Ragione, M.J. Woodward and A.O. Henriques, 2005. Screening for Bacillus isolates in the broiler gastrointestinal tract. Applied Environ. Microbiol., 71: 968-978.
CrossRefDirect Link - Bentley, R. and R. Meganathan, 1982. Biosynthesis of vitamin K (Menaquinone) in bacteria. Microbiol. Rev., 46: 241-280.
Direct Link - Bergh, O., K.E. Naas and T. Harboe, 1994. Shift in the intestinal microflora of Atlantic halibut (Hippoglossus hippoglossus) larvae during first feeding. Can. J. Fish Aquat. Sci., 51: 1899-1903.
CrossRefDirect Link - Bogut, I., Z. Milakovic, Z.I. Bukvic, S. Brkic and R. Zimmer, 1998. Influence of probiotic (Streptococcus faecium M74) on growth and content of intestinal microflora in carp (Cyprinus carpio). Czech J. Anim. Sci., 43: 231-235.
Direct Link - Bondad-Reantaso, M.G., S.C. Lumanlan, J.M. Natividad and M.J. Phillips, 1992. Environmental Monitoring of the Epizootic Ulcerative Syndrome (EUS) in Fish from Munoz, Nueva Ecija in the Philippines. In: Diseases in Asian Aquaculture, Shariff, M., R.P. Subasinghe and J.R. Arthur (Eds.). Fish Health Section, Asian Fish Soc., Manila, The Philippines, pp: 475-490.
- Bruun, M.S., A.S. Schmidt, L. Madsen and I. Dalsgaard, 2000. Antimicrobial resistance patterns in Danish isolates of Flavobacterium psychrophilum. Aquaculture, 187: 201-212.
CrossRef - Bruno, M.E.C. and T.J. Montville, 1993. Common mechanistic action of bacteriocins from lactic acid bacteria. Applied Environ. Microbiol., 59: 3003-3010.
PubMedDirect Link - Brunt, J. and B. Austin, 2005. Use of a probiotic to control lactococcosis and streptococcosis in rainbow trout, Oncorhynchus mykiss (Walbaum). J. Fish. Dis., 28: 693-701.
CrossRefDirect Link - Burrells, C., P.D. Williams and P.F. Forno, 2001. Dietary nucleotides: A novel supplement in fish feeds. Effects on resistance to disease in salmonids. Aquaculture, 199: 159-169.
CrossRef - Burrells, C., P.D. Williams, P.J. Southgate and S.L. Wadsworth, 2001. Dietary nucleotides: A novel supplement in fish feeds. Effects on vaccination, salt water transfer, growth rates and physiology of Atlantic salmon (Salmo salar L.). Aquaculture, 199: 171-184.
CrossRef - Burridge, L.E. and K. Haya, 1995. A review of Di-n-butylphthalate in the aquatic environment: Concerns regarding its use in salmonid aquaculture. J. World Aqua. Soc., 26: 1-13.
CrossRef - Capone, D.G., D.P. Weston, V. Miller and C. Shoemaker, 1996. Antibacterial residues in marine sediments and invertebrates following chemotherapy in aquaculture. Aquaculture, 145: 55-75.
CrossRefDirect Link - Chabrillon, M., S. Arijo, P. Diaz-Rosales, M.C. Balebona and M.A Morinigo, 2006. Interference of listonella anguillarum with potential probiotic microorganisms isolated from farmed gilthead seabream (Sparus aurata, L.). Aquacult. Res., 37: 78-86.
CrossRef - Zhenming, C., L. Zhiqiang, G. Lingmei, G. Fang, M. Chunling, W. Xianghong and L. Haifeng, 2006. Marine yeasts and their applications in mariculture. J. Ocean Uni. China, 5: 251-256.
CrossRef - Chinabut, S., R.J. Roberts, G.R. Willoughby and M.D. Pearson, 1995. Histopathology of Snakehead, Channa striatus (Bloch), experimentally infected with the specific Aphanomyces fungus associated with Epizootic Ulcerative Syndrome (EUS) at different temperatures. J. Fish. Dis., 18: 41-47.
CrossRefDirect Link - Chowdhury, M.B.R., M. Muniruzzaman, U.A. Zahura, K.Z.A. Habib and M.D. Khatun, 2003. Ulcer type of disease in the fishes of small scale farmers pond in Bangladesh. Pak. J. Biol. Sci., 6: 544-550.
Direct Link - Chythanya, R., I. Karunasagar and I. Karunasagar, 2002. Inhibition of shrimp pathogenic vibrios by a marine Pseudomonas I-2 strain. Aquaculture, 208: 1-10.
CrossRef - Conway, P.L., 2001. Prebiotics and human health: The state-of-the-art and future perspectives. Scand. J. Nutr., 45: 13-21.
Direct Link - Dahan, S., G. Dalmasso, V. Imbert, J.F. Peyron, P. Rampal and D. Czerucka, 2003. I>Saccharomyces boulardii interferes with enterohemorrhagic Escherichia coli induced signaling pathways in T84 cells. Infect. Immune., 71: 766-773.
CrossRef - Noh, D.O., S.H. Kim and S.E. Gilliland, 1997. Incorporation of cholesterol into the cellular membrane of Lactobacillus acidophilus ATCC 43121. J. Dairy Sci., 80: 3107-3113.
CrossRefDirect Link - Davies, R.L. and G.N. Frerichs, 1989. Morphological and biochemical differences among isolates of Yersinia ruckeri obtained from wide geographical areas. J. Fish Dis., 12: 357-365.
CrossRefDirect Link - Dhanaraj, M., M.A.K. Haniffa, C.M. Ramakrishnan and S.V.A. Singh, 2008. Microbial flora from the Epizootic Ulcerative Syndrome (EUS) infected murrel Channa striatus (Bloch, 1797) in Tirunelveli region. Turk. J. Vet. Anim. Sci., 32: 221-224.
Direct Link - Direkbusarakom, S., M. Yoshimizu, Y. Ezura, L. Ruangpah and Y. Danayadol, 1998. Vibrio spp., the dominant flora in shrimp hatchery against some fish pathogenic viruses. J. Mar. Biotechnol., 6: 266-267.
Direct Link - Dopazo, C.P., M.L. Lemos, C. Lodeiros, J. Bolinches, J.L. Barja and A.E. Toranzo, 1988. Inhibitory activity of antibiotic-producing marine bacteria against fish pathogens. J. Applied Bacteriol., 65: 97-101.
CrossRef - Douillet, P.A., 2000. Bacterial additives that consistently enhance rotifer growth under synxenic culture conditions: 2. Use of single and multiple bacterial probiotics. Aquaculture, 182: 241-248.
CrossRef - Douillet, P.A. and C.J. Langdon, 1994. Use of a probiotic for the culture of larvae of the pacific oyster (Crassostrea gigas Thunberg). Aquaculture, 119: 25-40.
CrossRefDirect Link - Duc, L.H., H.A. Hong, T.M. Barbosa, A.O. Henriques and S.M. Cutting, 2004. Characterization of Bacillus probiotics available for human use. Applied Environ. Microbiol., 70: 2161-2171.
CrossRef - Eldar, A., A. Horovitcz and H. Bercovier, 1997. Development and efficacy of a vaccine against Streptococcus iniae infection in farmed rainbow trout. Vet. Immunol. Immunopathol., 56: 175-183.
CrossRefDirect Link - Frerichs, G.N., J.A. Stewart and R.O. Collins, 1985. A typical infection of rainbow trout, Salmo gairdneri Richardson, with Yersinia ruckeri. J. Fish Dis., 8: 383-387.
CrossRef - Fukami, K., T. Nishijima and Y. Ishida, 1997. Stimulative and inhibitory effects of bacteria on the growth of microalgae. Hydrobiologia, 358: 185-191.
CrossRefDirect Link - Fuller, R., 1989. Probiotics in man and animals. J. Applied Bacteriol., 66: 365-378.
CrossRefPubMedDirect Link - Gatesoupe, F.J., 1997. Siderophore production and probiotic effect of Vibrio sp. associated with turbot larvae, Scophthalmus maximus. Aquat. Living Resour., 10: 239-246.
CrossRef - Gatesoupe, F.J., 1999. The use of probiotics in aquaculture. Aquaculture, 180: 147-165.
CrossRefDirect Link - Gatesoupe, F.J., 2002. Probiotic and formaldehyde treatments of Artemia nauplii as food for larval pollack, Pollachius pollachius. Aquaculture, 212: 347-360.
CrossRefDirect Link - Gatesoupe, F.J., 1993. Bacillus sp. Spores as Food Additive for the Rotifer Brachionus plicatilis: Improvement of their Bacterial Environment and their Dietary Value for Larval Turbot, Scopthalamus maximus L. In: Fish Nutrition in Practice, Kaushik, S. (Ed.). Institut National de la Recherche Agronomique, Paris, pp: 561-568.
- Gatesoupe, F.J., 1991. The effect of three strains of lactic bacteria on the production rate of rotifers, Brachionus plicatilis and their dietary value for larval turbot, Scophthalmus maximus. Aquaculture, 96: 335-342.
CrossRefDirect Link - Gatesoupe, F.J., 1994. Lactic acid bacteria increase the resistance of turbot larvae, Scophthalmus maximus, against pathogenic Vibrio sp. Aquat. Living Resour., 7: 277-282.
CrossRefDirect Link - Geovanny, D.G.R., J.L. Balczar and S. Ma, 2007. Probiotics as control agents in aquaculture. J. Ocean Univ. China, 6: 76-79.
CrossRefDirect Link - Ghosh, S., A. Sinha and C. Sahu, 2008. Bioaugmentation in the growth and water quality of livebearing ornamental fishes. Aqua. Int., 16: 393-403.
CrossRef - Gibson, L.F., J. Woodworth and A.M. George, 1998. Probiotic activity of Aeromonas media on the Pacific oyster, Crassostrea gigas, when challenged with Vibrio tubiashii. Aquaculture, 169: 111-120.
CrossRefDirect Link - Gildberg, A., H. Mikkelsen, E. Sandaker and E. Ringo, 1997. Probiotic effect of lactic acid bacteria in the feed on growth and survival of fry of Atlantic cod (Gadus morhua). Hydrobiologia, 352: 279-285.
CrossRefDirect Link - Gildberg, A. and H. Mikkelsen, 1998. Effects of supplementing the feed to Atlantic cod (Gadus morhua) fry with lactic acid bacteria and immuno-stimulating peptides during a challenge trial with Vibrio anguillarum. Aquaculture, 167: 103-113.
CrossRefDirect Link - Gildberg, A., A. Johansen and J. Bogwald, 1995. Growth and survival of Atlantic salmon (Salmo salar) fry given diets supplemented with fish protein hydrolysate and lactic acid bacteria during a challenge trial with Aeromonas salmonicida. Aquaculture, 138: 23-34.
Direct Link - Gomez-Gil, B., A. Roque and J.F. Turnbull, 2000. The use and selection of probiotic bacteria for use in the culture of larval aquatic organisms. Aquaculture, 191: 259-270.
CrossRefDirect Link - Gram, L., J. Melchiorsen, B. Spanggaard, I. Huber and T. Nielsen, 1999. Inhibition of Vibrio anguillarum by Pseudomonas fluorescens strain AH2 - a possible probiotic treatment of fish. Appl. Environ. Microbiol., 65: 969-973.
Direct Link - Grisez, L., J. Reyniers, L. Verdonck, J. Swings and F. Ollevier, 1997. Dominant intestinal microflora of sea bream and sea bass larvae, from two hatcheries, during larval development. Aquacult., 155: 387-399.
CrossRef - Gullian, M., F. Thompson and J. Rodriguez, 2004. Selection of probiotic bacteria and study of their immunostimulatory effect in Penaeus vannamei. Aquaculture, 233: 1-14.
CrossRefDirect Link - Huys, L., P. Dhert, R. Robles, F. Ollevier, P. Sorgeloos and J. Swings, 2001. Search for beneficial bacterial strains for turbot (Scophthalmus maximus L.) larviculture. Aquaculture, 193: 25-37.
CrossRef - Jha, A.K., A.K. Pal, N.P. Sahu, S. Kumar and S.C. Mukherjee, 2007. Haemato-immunological responses to dietary yeast RNA, Ω-3 fatty acid and β-carotene in Catla catla juveniles. Fish Shellfish Immunol., 23: 917-927.
CrossRef - Jiravanichpaisal, P., P. Chuaychuwong and P. Menasveta, 1997. The use of Lactobacillus sp. as the probiotic bacteria in the giant tiger shrimp (Penaeus monodon Fabricius). Proceedings of the Poster Session of the 2nd Asia-Pacific Marine Biotechnology Conference and 3rd Asia-Pacific Conference on Algal Biotechnology, May 7-10, Phuket, Thailand, pp: 1-16.
- Joborn A., J.C. Olsson, A. Westerdahl, P.L. Conway and S. Kjelleberg, 1997. Colonization in the fish intestinal tract and production of inhibitory substances in intestinal mucus and faecal extracts by Carnobacterium sp. strain K1. J. Fish Dis., 20: 383-392.
CrossRef - Ampofo, J.A. and G.C. Clerk, 2002. Infestation of fish-culturing communities with fish-borne bacteria: The Ghanain case. I. J. Environ. Health Res., 12: 277-282.
CrossRef - Karunasagar, I., G. Sugumar and I. Karunasagar, 1995. Viruilence Characters of Aeromonas sp. Isolated from EUS-Affected Fish. In: Diseases in Asian Aquaculture II, Shariff, M., J.R. Arthur and R.P. Subasinghe (Eds.). Fish Health Section, Asian Fisheries Society, Manila, The Philippines, pp: 307-314.
- Kennedy, S.B., J.W. Jr. Tucker, C.L. Neidig, G.K. Vermeer, V.R. Cooper, J.L. Jarrell and D.G. Sennett, 1998. Bacterial management strategies for stock enhancement of warmwater marine fish: A case study with common snook (Centropomus undecimalis). B. Mar. Sci., 62: 573-588.
Direct Link - Klesius, P.H., C.A. Shoemaker and J.J. Evans, 2000. Efficacy of a single and combined Streptococcus iniae isolate vaccine administered by intraperitoneal and intramuscular routes in Tilapia (Oreochromis niloticus). Aquaculture, 88: 237-246.
CrossRef - Kubitza, F., L.L. Lovshin and R.T. Lovell, 1997. Identification of feed enhancers for juvenile largemouth bass Micropterus salmoides. Aquaculture, 148: 191-200.
CrossRef - Kumari, J. and P.K. Sahoo, 2005. High dietary vitamin C affects growth, non-specific immune responses and disease resistance in Asian catfish, Clarias batrachus. Mol. Cell. Biochem., 280: 25-33.
Direct Link - Cormack, W.P.M. and E.R. Fraile, 1990. Bacterial flora of newly caught antarctic fish Notothenia neglecta. Polar Biol., 10: 413-417.
CrossRef - Maeda, M. and I.C. Liao, 1994. Microbial processes in aquaculture environment and their importance for increasing crustacean production. J. ARQ., 28: 283-288.
Direct Link - Maeda, M., K. Nogami, M. Kanematsu and K. Hirayama, 1997. The concept of biological control methods in aquaculture. Hydrobiology, 358: 285-290.
Direct Link - Makridis, P., O. Bergh, J. Skjermo and O. Vadstein, 2001. Addition of bacteria bioencapsulated in Artemia metanauplii to a rearing system for halibut larvae. Aquacult. Int., 98: 225-235.
Direct Link - Calo-Mata, P., S. Arlindo, K. Boehme, T. de Miguel, A. Pascoal and J. Barros-Velazquez, 2008. Current applications and future trends of lactic acid bacteria and their bacteriocins for the biopreservation of aquatic food products. Food Bioprocess. Technol., 1: 43-63.
CrossRefDirect Link - McFarland, L.V., 2006. Meta-analysis of probiotics for the prevention of antibiotic associated diarrhea and the treatment of Clostridium difficile disease. Am. J. Gastroenterol., 101: 812-822.
Direct Link - Hjelm, M., O. Bergh, A. Riaza, J. Nielsen and J. Melchiorsen et al., 2004. Selection and identification of autochthonous potential probiotic bacteria from turbot larvae (Scophthalmus maximus) rearing units. Syst. Applied Microbiol., 27: 360-371.
CrossRefDirect Link - Miranda, C.D. and R. Zemelman, 2002. Bacterial resistance to oxytetracycline in Chilean salmon farming. Aquaculture, 212: 31-47.
CrossRef - Mukai, T., T. Asasaka, E. Sato, K. Mori, M. Matsumoto and H. Ohori, 2002. . Inhibition of binding of Helicobacter pylori to the glycolipid receptors by probiotic Lactobacillus reuteri. Fems Immunol. Med. Microbiol., 32: 105-110.
PubMed - Munro, P.O., A. Barbour and T.H. Blrkbeck, 1994. Comparison of the gut bacterial flora of start feeding larval turbot under different conditions. J. Applied Bacteriol., 77: 560-566.
Direct Link - Munro, P.D., A. Barbour and T.H. Birkbeck, 1995. Comparison of the growth and survival of larval turbot in the absence of culturable bacteria with those in the presence of Vibrio anguillarum, Vibrio alginolyticus, or a marine Aeromonas sp. Appl. Environ. Microbiol., 61: 4425-4428.
Direct Link - Muroga, K., M. Higashi and H. Keitoku, 1987. The isolation of intestinal microflora of farmed red seabream (Pagrus major) and black seabream (Acanthopagrus schlegeli) at larval and juvenile stages. Aquaculture, 65: 79-88.
CrossRef - Nair, S., K. Tsukamoto and U. Shimidu, 1985. Distribution of bacteriolytic bacteria in the coastal marine environments of Japan. Bull. Jpn. Soc. Sci. Fish., 51: 1469-1473.
Direct Link - Nayak, S.K., P. Swain and S.C. Mukherjee, 2007. Effect of dietary supplementation of probiotic and vitamin C on the immune response of Indian major carp, Labeo rohita (Ham.). Fish Shellfish Immunol., 23: 892-896.
PubMed - Nicolas, J.L., E. Robic and D. Ansquer, 1989. Bacterial flora associated with a tropical chain consisting of microalgae, rotifers and turbot larvae: Influence of bacteria on larval survival. Aquaculture, 83: 237-248.
CrossRef - Nikoskelainen, S., A.C. Ouwehand, G. Bylund, S. Salminen and E.M. Lilius, 2003. Immune enhancement in rainbowtrout (Oncorhychus mykiss) by potential probiotic bacteria (Lactobacillus rhamnosus). Fish Shellfish Immunol., 15: 443-452.
CrossRef - Nogami, K. and M. Maeda, 1992. Bacteria as biocontrol agents for rearing larvae of the crab Portunus trituber culatus. Can. J. Fish Aquat. Sci., 4: 2373-2376.
CrossRefDirect Link - Olafsen, J.A., 2001. Interactions between fish larvae and bacteria in marine aquaculture. Aquaculture, 200: 223-247.
CrossRefDirect Link - Ottesen, O.H. and J.A. Olafsen, 2000. Effects on survival and mucous cell proliferation of Atlantic halibut, Hippoglossus hippoglossus L., larvae following microflora manipulation. Aquaculture, 187: 225-238.
CrossRef - Ouwehand, A.C., P.V. Kirjavainen, M.M. Gronlund, E. Isolauri and S.J. Salminen, 1999. Adhesion of probiotic micro-organisms to intestinal mucus. Int. Dairy J., 9: 623-630.
CrossRefDirect Link - Ouwehand, A.C., E.M. Tuomola, S. Tolkko and S. Salminen, 2001. Assessment of adhesion properties of novel probiotic strains to human intestinal mucus. Int. J. Food Microbiol., 64: 119-126.
CrossRef - Panigrahi, A. and S. Azad, 2007. Microbial intervention for better fish health in aquaculture: The Indian scenario. Fish Physiol. Biochem., 33: 429-440.
CrossRef - Pinchuk, I.V., P. Bressollier, B. Verneuil, B. Fenet, I.B. Sorokulova, F. Megraud and M.C. Urdaci, 2001. In vitro anti-Helicobacter pylori activity of the probiotic strain Bacillus subtilis 3 is due to secretion of antibiotics. Antimicrob. Agents Ch., 45: 3156-3161.
Direct Link - Prayitno, S.B. and J.W. Latchford, 1995. Experimental infections of crustaceans with luminous bacteria related to Photobacterium and Vibrio species Effect of salinity and pH on infectivity. Aquaculture, 132: 105-112.
CrossRef - Raa, J., 1996. The use of immunostimulatory substances in fish and shellfish farming. Rev. Fish. Sci., 4: 229-288.
CrossRefDirect Link - Lalloo, R., D. Maharajh, J. Gorgens and N. Gardiner, 2008. Functionality of a Bacillus cereus biological agent in response to physiological variables encountered in aquaculture. Applied Microbiol. Biotechnol., 79: 111-118.
PubMed - Kumar, R., S.C. Mukherjee, K.P. Prasad and A.K. Pal, 2006. Evaluation of Bacillus subtilis as a probiotic to Indian major carp Labeo rohita (Ham.). Aquacult. Res., 37: 1215-1221.
CrossRef - Ramadan, A., N.A. Afifi, M.M. Moustafa and A.M. Samy, 1994. The effect of ascogen on the immune response of tilapia fish to Aeromonas hydrophila vaccine. Fish Shellfish Immunol., 4: 159-165.
CrossRef - Ramirez, R.F. and B.A. Dixon, 2003. Enzyme production by obligate intestinal anaerobic bacteria isolated from oscars (Astronotus ocellatus), angelfish (Pterophyllum scalare) and southern flounder (Paralichthys lethostigma). Aquaculture, 227: 417-426.
CrossRef - Ranzani-Paiva, M.J.T., C.M. Ishikawa, A.C. das Eiras and V.R. da Silveira, 2004. Effects of an experimental challenge with Mycobacterium marinum on the bood parameters of Nile tilapia, Oreochromis nitoticus (Linnaeus, 1757). B. Archives Bio. Tech., 47: 945-953.
Direct Link - Reid, R.T., D.H. Live, D.J. Faulkner and A. Butler, 1993. A siderophore from a marine bacterium with an exceptional ferric ion affinity constant. Nature, 366: 455-458.
CrossRef - Rengpipat, S., T. Rueangruklikhit and S. Piyatiratitivorakul, 2008. Evaluations of lactic acid bacteria as probiotics for juvenile seabass Lates calcarifer. Aquacult. Res., 39: 134-143.
CrossRef - Ringo, E., 1999. Does Carnobacterium divergens isolated from Atlantic salmon, Salmo salar L., colonise the gut of early developing turbot, Scophthalmus maximus L., larvae?. Aqua. Res., 30: 229-232.
CrossRef - Ringo, E. and F.J. Gatesoupe, 1998. Lactic acid bacteria in fish: A review. Aquaculture, 160: 177-203.
CrossRefDirect Link - Ringo, E. and E. Strom, 1994. Microflora of Arctic charr, Salvelinus alpinus (L.): Gastrointestinal microflora of free-living fish and effect of diet and salinity on intestinal microflora. Aquacult. Res., 25: 623-629.
CrossRefDirect Link - Ringo, E., T.H. Birkbeck, P.O. Munro, O. Vadstein and K. Hjelmeland, 1996. The effect of early exposure to Vibrio pelagius on the aerobic bacterial flora of turbot, Scophthalmus maximus (L.) larvae. J. Applied Microbiol., 81: 207-211.
CrossRef - Ringo, E., R.E. Olsen, O. Overli and F. Lovik, 1997. Effect of dominance hierarchy formation on aerobic microbiota associated with epithelial mucosa of subordinate and dominant individuals of Arctic charr, Salvelinus alpinus (L.). Aqua. Res., 28: 901-904.
Direct Link - Ringo, E., E. Strom and J.A. Tabachek, 1995. Intestinal microflora of salmonids: A review. Aquacult. Res., 26: 773-789.
CrossRef - Riquelme, C.E., M.A. Jorquera, A.I. Rojas, R.E. Avendano and N. Reyes, 2001. Addition of inhibitor-producing bacteria to mass cultures of Argopecten purpuratus larvae (Lamarck, 1819). Aquaculture, 1992: 111-119.
CrossRef - Riquelme, C., R. Araya and R. Escribano, 2000. Selective incorporation of bacteria by Argopecten purpuratus larvae: Implications for the use of probiotics in culturing systems of the Chilean scallop. Aquaculture, 181: 25-36.
CrossRef - Riquelme, C., R. Araya, N. Vergra, A. Rojas, M. Guaita and M. Candia, 1997. Potential probiotic strain in the culture of Chilean scallop, Argopecten purpuratus (Lamark 1819). Aquaculture, 154: 17-26.
CrossRef - Roberts, R.J., L.G. Willoughby and S. Chinabut, 1993. Mycotic aspects of Epizootic Ulcerative Syndrome (EUS) of Asian fishes. J. Fish Dis., 16: 169-183.
CrossRef - Robertson, P.A.W., C.O. Dowd, C. Burrells, P. Williams and B. Austin, 2000. Use of Carnobacterium sp. as a probiotic for Atlantic salmon (Salmo salar L.) and rainbow trout (Oncorhynchus mykiss, Walbaum). Aquaculture, 185: 235-243.
CrossRefDirect Link - Ruiz-Ponte, C., J.F. Samain, J.L. Sanchez and J.L. Nicolas, 1999. The benefit of a Roseabacter species on the survival of scallop larvae. Mar. Biotechnol., 1: 52-59.
CrossRef - Hameed, A.S.S. and G. Balasubramanian, 2000. Antibiotic resistance in bacteria isolated from Artemia nauplii and efficacy of formaldehyde to control bacterial load. Aquaculture, 183: 195-205.
CrossRef - Sakai, M., T. Otubo, S. Atsuta and M. Kobayashi, 1995. Enhancement of resistance to vibriosis in rainbow trout, Oncorhynchus mykiss (Walbaum), by oral administration of Clostridium butyricum bacterin. J. Fish Dis., 18: 187-190.
CrossRefDirect Link - Salminen, S., A. von Wright, L. Morelli, P. Marteau and D. Brassart et al., 1998. Demonstration of safety of probiotics: A review. Int. J. Food Microbiol., 44: 93-106.
CrossRefDirect Link - Salminen, S., A. Ouwehand, Y. Benno and Y.K. Lee, 1999. Probiotics: How should they be defined? Trends Food Sci. Technol., 10: 107-110.
CrossRefDirect Link - Sanders, M.E., 2000. Considerations for use of probiotic bacteria to modulate human health. J. Nutr., 130: 384S-390S.
Direct Link - Sanderson, I.R. and Y. He, 1994. Nucleotides uptake and metabolism by intestinal epithelial cells. J. Nutr., 124: 131-137.
Direct Link - Rengpipat, S., S. Rukpratanporn, S. Piyatiratitivorakul and P. Menasaveta, 2000. Immunity enhancement in black tiger shrimp (Penaeus monodon) by a probiont bacterium (Bacillus S11). Aquaculture, 191: 271-288.
CrossRef - Scharrer, E. and T. Lutz, 1990. Effects of short chain fatty acids and K on absorption of Mg and other cations by the colon and caecum. Z. Ernahrungswiss., 29: 162-168.
CrossRef - Selvaraj, V., K. Sampath and V. Sekar, 2005. Administration of yeast glucan enhances survival and some non-specific and specific immune parameters in carp (Cyprinus carpio) infected with Aeromonas hydrophila. Fish Shellfish Immunol., 19: 293-306.
Direct Link - Servin, A.L. and M.H. Coconnier, 2003. Adhesion of probiotic strains to the intestinal mucosa and interaction with pathogens. Best Pract. Res. Clin. Gastroenterol., 17: 741-754.
CrossRefPubMedDirect Link - Siwicki, A.K., D.P. Anderson and G.L. Rumsey, 1994. Dietary intake of immunostimulants by rainbow trout affects non-specific immunity and protection against furunculosis. Vet. Immunol. Immunopathol., 41: 125-139.
PubMedDirect Link - Skjermo, J. and O. Vadstein, 1999. Techniques for microbial control in the intensive rearing of marine larvae. Aquaculture, 177: 333-343.
CrossRefDirect Link - Smith, P., 1996. Is sediment deposition the dominant fate of OTC used in marine salmonid farms. A review of available evidence. Aquaculture, 146: 157-169.
CrossRef - Smith, P. and S. Davey, 1993. Evidence for competitive exclusion of Aeromonas salmonicida from fish with stress - inducible furunculosis by a florescent Pseudomonad. J. Fish Dis., 16: 521-524.
Direct Link - Strom, E. and E. Ringo, 1993. Changes in the Bacterial Composition of Early Developing Cod, Gadus morhua (L.) Larvae Following Inoculation of Lactobacillus plantarum into the Water. In: Physiology and Biochemical Aspects of Fish Development, Walther, B.T. and H.J. Fyhn (Eds.). University of Bergen, Bergen, Norway, pp: 226-228.
- Sugita, H., K. Shibuya, H. Shimooka and Y. Deguchi, 1996. Antibacterial abilities of intestinal bacteria in freshwater cultured fish. Aquaculture, 145: 195-203.
CrossRef - Tanasomwang, V. and K. Muroga, 1989. Intestinal microflora of rockfish Sebastes schlegeli, tiger puffer Takifugu rubripes and red grouper Epinephelus akaara at their larval and juvenile stages. Nippon. Suisan. Gakk., 55: 1371-1377.
Direct Link - Pal, J., 1996. The fish disease, epizootic ulcerative syndrome: The present state of research and its impact in North Bengal. Proceedings of the National Workshop on Fish and Prawn Diseases, Epizootics and Quarantine Adoption in India. (FPDEQAI'96), Central Inland Capture Research Institute, Barrackpore, pp: 95-98.
- Topping, D.L. and P.M. Clifton, 2001. Short-chain fatty acids and human colonic function: Roles of resistant starch and non-starch polysaccharides. Physiol. Rev., 81: 1031-1064.
PubMed - Van der Waaij, D. and C.E. Nord, 2000. Development and persistence of multi-resistance to antibiotics in bacteria; an analysis and a new approach to this urgent problem. Int. J. Antimicrob. Ag., 16: 191-197.
Direct Link - Vandenbergh, P.A., 1993. Lactic acid bacteria, their metabolic products and interference with microbial growth. FEMS Microbiol. Rev., 12: 221-238.
CrossRefDirect Link - Venkat, H.K., N.P. Sahu and K.K. Jain, 2004. Effect of feeding Lactobacillus-based probiotics on the gut microflora, growth and survival of postlarvae of Macrobrachium rosenbergii (de Man). Aquacult. Res., 35: 501-507.
CrossRefDirect Link - Verma, R.K., V. Jakhar and S.K. Gahlawat, 2006. Epizootic ulcerative syndrome in fresh water fishes in Haryana. Proceedings of the 21st National Symposium of Indian Society of Life Sciences on Recent Advances in Life Sciences and Environmental Conservation in Welfare of Human Society, February 3-5, 2006, K.U. Kurukshetra, pp: 67-68.
- Verschuere, L., G. Rombaut, G. Huys, J. Dhont, P. Sorgeloos and W. Verstraete, 1999. Microbial control of the culture of Artemia juveniles through preemptive colonization by selected bacterial strains. Appl. Environ. Microbiol., 65: 2527-2533.
Direct Link - Verschuere, L., G. Rombaut, P. Sorgeloos and W. Verstraete, 2000. Probiotic bacteria as biological control agents in aquaculture. Microbiol. Mol. Biol. Rev., 64: 655-671.
CrossRefPubMedDirect Link - Vijayabaskar, P. and S.T. Somasundaram, 2008. Isolation of bacteriocin producing lactic acid bacteria from fish gut and probiotic activity against common fresh water fish pathogen Aeromonas hydrophila. Biotechnology, 7: 124-128.
CrossRefDirect Link - Vine, N.G., W.D. Leukes and H. Kaiser, 2004. In vitro growth characteristics of five candidate aquaculture probiotics and two fish pathogens grown on fish intestinal mucus. Fems Microbiol. Lett., 231: 145-152.
PubMed - Vishwanath, T.S., C.V. Mohan and K.M. Shankar, 1998. Epizootic Ulcerative Syndrome (EUS) associated with a fungal pathogen, in Indian fishes: Histopathology-a cause for invasiveness. Aquaculture, 165: 1-9.
CrossRef - Welker, T.L., C.A. Sheomaker, C.R. Arias and P.H. Kleius, 2005. Transmission and detection of Flavobacterium columnare in Channel catfish (Ictalurus punctatus). Dis. Aquacult. Org., 63: 129-138.
Direct Link - Westerdahl, A., J.C. Olsson, S. Kjelleberg, P.L. Conway, 1991. Isolation and characterization of turbot (Scophtalmus maximus)-associated bacteria with inhibitory effects against Vibrio anguillarum. Applied Environ. Microbiol., 57: 2223-2228.
Direct Link - Williams, S.T. and J.C. Vickers, 1986. The ecology of antibiotic production. Microb. Ecol., 12: 43-52.
CrossRefDirect Link - Bansal, S.K., S.R. Verma, A.K. Gupta and R.C. Dalela, 1979. Physiological dysfunction of the haemopoietic system in a fresh water teleost, Labeo rohita, following chronic chlordane exposure. Part I-Alterations in certain haematological parameters. Bull Environ. Contam. Toxicol., 22: 666-673.
PubMedDirect Link - Garriques, D. and G. Arevalo, 1995. An evaluation of the production and use of a live bacterial isolate to manipulate the microbial flora in the commercial production of Penaeus vannzamei postlarvae in Ecuador. Proceedings of the Special Session on Shrimp Farming, (SSSF'95), Publ. World Aquaculture Society, Baton Rouge, Louisiana, USA, pp: 53-59.
- Lumanlan-Mayo, S.C., R.B. Callinan, J.O. Paclibare, E.S. Catup and G.C. Fraser, 1997. Epizootic Ulcerative Syndrome (EUS) in Rice-Fish Culture Systems: An Overview of Field Experiments 1993-1995. In: Diseases in Asian Aquaculture III: Fish Health Section, Flegel, T.W. and I.H. MacRae (Eds.). Asian Fisheries Society, Manila, The Philippines, pp: 129-138.
- Iqbal, M.M., M.B.R. Chaowdhury, M.A. Islam, M. Baqui, S.M.R. Karim, K. Tajima and Y. Ezure, 1999. Seasonal fluctuations of motile Aeromonads and Pseudomonads in a cultured pond of mrigal, Cirrhinus mrigala in Bangaldesh. J. Environl. Health Res., 10: 267-279.
Direct Link - Isolauri, E., Y. Sutas, P. Kankaanpaa, H. Arvilommi and S. Salminen, 2001. Probiotics: Effects on immunity. Am. J. Clin. Nutr., 73: 444s-450s.
PubMedDirect Link - Hong, H.A., L.H. Duc and S.M. Cutting, 2005. The use of bacterial spore formers as probiotics. FEMS Microbiol. Rev., 29: 813-835.
CrossRefPubMedDirect Link - John, R.P., K.M. Nampoothiri and A. Pandey, 2007. Fermentative production of lactic acid from biomass: An overview on process developments and future perspectives. Applied Microbial. Biotechnol., 74: 524-534.
CrossRef - Bandyopadhyay, P. and P.K.D. Mohapatra, 2009. Effect of a probiotic bacterium Bacillus circulans PB7 in the formulated diets: On growth, nutritional quality and immunity of Catla catla (Ham.). Fish Physiol. Biochem., 35: 467-478.
CrossRefDirect Link - Yazid, A.M., A.B. Salina, M. Shuhaimi, H. Osman and J. Normah, 1999. Inhibitory effects of probiotic bacteria against selected food-borne pathogens. Pak. J. Biol. Sci., 2: 660-663.
CrossRefDirect Link - Chang, M.H. and T.C. Chen, 2003. Reduction of broiler house malodor by direct feeding of a lactobacilli containing probiotic. Int. J. Poult. Sci., 2: 313-317.
CrossRefDirect Link - El-Naggar, M.Y.M., 2004. Comparative study of probiotic culture to control the growth of Escherichia coli O157: H7 and Salmonella typhimurium. Biotechnology, 3: 173-180.
CrossRefDirect Link - Ghafoor, A., S. Naseem, M. Younus and J. Nazir, 2005. Immunomodulatory effect of multistrain probiotics (ProtexinTM) on broiler chicken vaccinated against avaian influenza virus (H9). Int. J. Poult. Sci., 4: 777-780.
CrossRefDirect Link - Igbasan, F.A., V.O. Oyetayo and O.O. Jawo, 2005. Faecal strain of lactobacillus acidophilus prevents diarrhoea and improved the health of rats challenged with clinical strain of Shigella dysenteriae. Pak. J. Biol. Sci., 8: 1507-1510.
CrossRefDirect Link - Kabir, S.M.L., M.M. Rahman, M.B. Rahman, M.Z. Hosain, M.S.I. Akand and S.K. Das, 2005. Viability of probiotics in balancing intestinal flora and effecting histological changes of crop and caecal tissues of broilers. Biotechnology, 4: 325-330.
CrossRefDirect Link - Trachoo, N. and C. Boudreaux, 2006. Therapeutic properties of probiotic bacteria. J. Biol. Sci., 6: 202-208.
CrossRefDirect Link - Anukam, K.C., 2007. Probiotic toxicity, any evidence?. J. Pharmacol. Toxicol., 2: 590-598.
CrossRefDirect Link - Deeseenthum, S., V. Leelavatcharamas and J.D. Brooks, 2007. Effect of feeding Bacillus sp. as probiotic bacteria on growth of giant freshwater prawn (Macrobrachium rosenbergii de Man). Pak. J. Biol. Sci., 10: 1481-1485.
CrossRefPubMedDirect Link - Anukam, K.C. and T.E. Koyama, 2007. Bile and acid tolerance of Lactobacillus plantarum KCA-1: A potential probiotical agent. Int. J. Dairy. Sci., 2: 275-280.
CrossRefDirect Link - Capcarova, M., A. Kolesarova, P. Massanyi and J. Kovacik, 2008. Selected blood biochemical and haematological parameters in turkeys after an experimental probiotic Enterococcus faecium M-74 strain administration. Int. J. Poult. Sci., 7: 1194-1199.
CrossRefDirect Link - Patel, P., T. Parekh and R. Subhash, 2008. Development of probiotic and symbiotic chocolate mousse: A functional food. Biotechnology, 7: 769-774.
CrossRefDirect Link - Raj, A.J.A., A.V. Suresh, K. Marimuthu and S. Appelbaum, 2008. Probiotic performance on fish fry during packing, transportation stress and post-transportation condition. J. Fish. Aquatic Sci., 3: 152-157.
CrossRefDirect Link - Radfar, M. and P. Farhoomand, 2008. The role of probiotic and source of lactose as feed additives on performance and gut improvement in broilers. Asian J. Anim. Vet. Adv., 3: 179-182.
CrossRefDirect Link - Soundarapandian, P. and S. Sankar, 2008. Effect of probiotics on the survival and production of black tiger shrimp Penaeus monodon (Fabricius). Int. J. Zool. Res., 4: 35-41.
CrossRefDirect Link - Vamanu, E., A. Vamanu, O. Popa, T. Vassu and R. Ghindea et al., 2008. Effect of the yeast and bacteria biomass on the microbiota in the rumen. Pak. J. Biol. Sci., 11: 2217-2223.
CrossRefPubMedDirect Link - Vali, N., 2009. Probiotic in quail nutrition: A review. Int. J. Poult. Sci., 8: 1218-1222.
CrossRefDirect Link - Abdelhamid, A.M., A.I. Mehrim, M.I. El-Barbary, S.M. Ibrahim and A.I. Abd El-Wahab, 2009. Evaluation of a New Egyptian probiotic by African catfish fingerlings. J. Environ. Sci. Technol., 2: 133-145.
CrossRefDirect Link - Al-Otaibi, M.M., 2009. Evaluation of some probiotic fermented milk products from Al-Ahsa markets, Saudi Arabia. Am. J. Food Technol., 4: 1-8.
CrossRefDirect Link - Raja, A., P. Gajalakshmi, M.M.M. Raja and M.M. Imran, 2009. Effect of Lactobacillus lactis cremoris Isolated from Kefir against Food Spoilage Bacteria. Am. J. Food Technol., 4: 201-209.
CrossRefDirect Link - Nikfar, S., M. Darvish-Damavandi and M. Abdollahi, 2010. A review and meta-analysis of the efficacy of antibiotics and probiotics in management of pouchitis. Int. J. Pharmacol., 6: 826-835.
CrossRefDirect Link - Vamanu, E. and A. Vamanu, 2010. Viability of the Lactobacillus rhamnosus IL1 strain in simulated gastrointestinal conditions. Int. J. Pharmacol., 6: 732-737.
CrossRefDirect Link - Agouz, H.M. and W. Anwer, 2011. Effect of biogen® and Myco-ad® on the growth performance of common carp (Cyprinus carpio) fed a mycotoxin contaminated aquafeed. J. Fish. Aquat. Sci., 6: 334-345.
CrossRefDirect Link - Bansal, G.R., V.P. Singh and N. Sachan, 2011. Effect of probiotic supplementation on the performance of broilers. Asian J. Anim. Sci., 5: 277-284.
CrossRefDirect Link - Yesillik, S., N. Yildirim, A. Dikici, A. Yildiz and S. Yesillik, 2011. Antibacterial effects of some fermented commercial and homemade dairy products and 0.9% lactic acid against selected foodborne pathogens. Asian J. Anim. Vet. Adv., 6: 189-195.
CrossRefDirect Link - Gharaei-Fathabad, E. and M. Eslamifar, 2011. Isolation and applications of one strain of Lactobacillus paraplantarum from tea leaves (Camellia sinensis). Am. J. Food Technol., 6: 429-434.
CrossRef - Lilly, D.M. and R.H. Stillwell, 1965. Probiotics: Growth-promoting factors produced by microorganisms. Science, 147: 747-748.
CrossRefDirect Link - Hung, A.T.Y., T.M. Su, C.W. Liao and J.J. Lu, 2008. Effect of probiotic combination fermented soybean meal on growth performance, lipid metabolism and immunological response of growing-finishing pigs. Asian J. Anim. Vet. Adv., 3: 431-436.
CrossRefDirect Link - Fazeli, Z., G. Azari-Takami and S.A. Fazeli, 2008. Effects of yeast probiotic (Thepax) enrichment on biochemical parameters of A. urmiana Naupli. Pak. J. Biol. Sci., 11: 643-647.
CrossRefPubMedDirect Link - Vine, N.G., W.D. Leukes, H. Kaiser, S. Daya, J. Baxter and T. Hecht, 2004. Competition for attachment of aquaculture candidate probiotic and pathogenic bacteria on fish intestinal mucus. J. Fish Dis., 27: 319-326.
CrossRefDirect Link - Kesarcodi-Watson, A., H. Kaspar, M.J. Lategan and L. Gibson, 2008. Probiotics in aquaculture: The need, principles and mechanisms of action and screening processes. Aquaculture, 274: 1-14.
CrossRefDirect Link - Moriarty, D.J.W., 1998. Control of luminous Vibrio species in penaeid aquaculture ponds. Aquaculture, 164: 351-358.
CrossRefDirect Link - Irianto, A. and B. Austin, 2002. Use of probiotics to control furunculosis in rainbow trout, Oncorhynchus mykiss (Walbaum). J. Fish Dis., 25: 333-342.
CrossRefDirect Link
sujani Reply
Hi
Very good work but I just want to know is there any work done on Effect of probiotics to prevent Argulus (Fish Lice).
Waiting for ur reply